
不同产地蒙古黄芪的非靶向代谢组学分析
2024-04-23 05:54宋诗娟雷振宏郭旭王圆圆燕翔牛景萍赵成萍梁建萍
山西农业科学 2024年1期
宋诗娟 雷振宏 郭旭 王圆圆 燕翔 牛景萍 赵成萍 梁建萍
摘要:黄芪(Astragalus membranaceus)是一种传统的道地药材,药用历史悠久,分布范围广,不同产地黄芪的药材品质和疗效相差甚远,次生代谢物的类型和含量是决定药材品质的物质基础。为了研究不同产地黄芪次级代谢物的差异,为解析黄芪品质形成提供重要依据,同时为优质黄芪的选育奠定基础,试验对来自山西浑源(S2-7)、山西五台(S2-9)、甘肃和政药园(S2-10)的3 种蒙古黄芪根进行基于LC-MS/MS 的非靶向代谢组学测定,利用主成分分析(PCA)和正交偏最小二乘法判别分析(OPLS-DA)方法进行差异代谢物筛选,并通过KEGG 数据库进行通路富集分析。结果显示,共鉴定出674 种代谢产物,分为29 类,主要包括萜类物质、黄酮类物质、苯丙素类物质和生物碱类物质。依据P<0.05、变量投影重要度(VIP)>1 筛选差异代谢物,甘肃黄芪(S2-10)与山西黃芪组(S2-7、S2-9)间筛选出81 种差异代谢物;山西五台(S2-9)和浑源(S2-7)2 个产地黄芪间筛选出67 种差异代谢物;这些代谢物在含量上存在显著差异。对通路进行富集分析可知,甘肃黄芪与山西黄芪组富集到3 条差异显著的代谢通路:异喹啉生物碱生物合成、吲哚生物碱生物合成以及烟酸盐和烟酰胺代谢通路。综上可见,不同产地黄芪次级代谢物的类型和含量存在一定差异。
关键词:蒙古黄芪;不同产地;LC-MS/MS;PCA;OPLS-DA;非靶向代谢组学;差异代谢物
中图分类号:R284文献标识码:A文章编号:1002?2481(2024)01?0043?12
黄芪是传统的大宗中药材,为豆科植物蒙古黄芪(Astragalus membranaceus(Fisch.)Bge. var. mon?gholicus(Bge.)Hsiao)或膜荚黄芪(A. membranaceus(Fisch.)Bge.)的干燥根,最早记载于《神农本草经》[1]。黄芪为多年生草本植物[2],从幼苗到收获周期为2~5 a,分布范围广,主要生长在我国山西、内蒙古、甘肃、陕西、宁夏等省份,具有丰富的遗传多样性[3-4]。迄今为止,已从黄芪中鉴定出多种代谢物,包括皂苷、类黄酮、多糖和氨基酸等,其主要生物活性成分为黄酮、皂苷和多糖[2]。更重要的是,黄芪具有止汗、抗利尿、抗肿瘤、抗氧化和抗炎等作用[5-9],黄芪在中成药、保健品、化妆品等领域倍受青睐,而其代谢成分是黄芪药材品质形成的重要物质基础,因此,开展黄芪代谢组学研究,对于黄芪种质资源鉴定、品质评价、新品种选育、代谢成分体外合成机制极为重要[10]。
代谢组学分析是一种基于高通量技术对整个生物体内小分子代谢物进行研究的方法,能够检测生物体代谢物并筛选出显著差异代谢物,在此基础上研究相应的代谢通路及其变化机制[11]。近年来代谢组学技术被广泛应用于中药材质量控制与品质评价、植物分类、资源鉴定、亲缘关系评估以及代谢调控网络等领域[10]。其中非靶向代谢组学主要通过寻找试验组和对照组之间的差异代谢产物以及分析代谢途径与表型之间的关系,从整体反映代谢物的变化,提供更广泛的代谢物覆盖,实现不同样品间代谢特征的比较,有利于发现新的代谢通路[12-14]。WU 等[2]基于UPLC-Q-Orbitrap 方法对蒙古黄芪进行综合转录组和代谢组学分析,共检测到5 435 种代谢物,阐明苯丙素生物合成、类黄酮生物合成和异黄酮生物合成的完整途径,发现了新的类黄酮衍生物,并推断出它们在黄芪中可能的合成途径。WANG 等[15]利用UHPLC-ESI-Q-TOF-MS/MS 方法,对蒙古黄芪和膜荚黄芪进行了化学鉴别,共检测到53 种差异代谢物,初步推断膜荚黄芪比蒙古黄芪有更强的药理活性。牛媛婧等[16]通过非靶向代谢组学方法分析不同产地党参之间的差异,筛选得到72 种差异代谢物,主要代谢通路为苯丙素生物合成。孟益德等[10]利用UPLC-QTOF/MS方法对不同产地杜仲雄花进行了非靶向代谢组学分析,共筛选出45 个显著差异代谢物,主要参与氨酰基tRNA 生物合成、类黄酮生物合成、黄酮和黄酮醇生物合成以及ABC 转运蛋白等代谢途径。但是运用非靶向代谢组学方法分析不同产地黄芪中代谢物的差异尚未见报道。
本研究基于实验室前期对来源于8 个省份的80 份不同蒙古黄芪种质资源的聚类分析[17],选择分属2 个亚群遗传背景差异较大的3 个种质为试验材料,分别为来自甘肃政药园的S2-10、来自山西五台的S2-9 与来自山西浑源的S2-7,采用LC-MS/MS 系统组合型四极杆Orbitrap 质谱技术,结合多元变量模式识别分析,检索常规数据库及测定公司自建的中药材数据库,从代谢组学的角度探讨不同产地蒙古黄芪有效成分积累的差异,旨在为挖掘和选育高品质黄芪资源奠定基础。
1材料和方法
1.1 试验材料与仪器
供试材料为3 种位于2 个亚群[17]的不同产地蒙古黄芪,于2018 年采自山西和甘肃,分别记为S2-7(山西省浑源县)、S2-9(山西省五台县)、S2-10(甘肃省和政药用植物园)。2019 年5 月采用种子直播种植于汾阳市山西农业大学经济作物研究所黄芪种质资源圃(111°47 ′42.13 ″E,37°14 ′55.90 ″N)内,2020 年11 月选取长势一致的健康黄芪植株,每个产地3 次重复,剪掉地上茎叶,清理根部表面泥土后带回实验室,测量基本生物量后烘干备用。
试验仪器包括:1290 UPHLC超高效液相(Ameri?can Agilent),Q Exactive Focus 高分辨质谱(Ameri?can Thermo Fisher Scientific),Heraeus Fresco17 离心机(American Thermo Fisher Scientific),BSA124S-CW 电子天平(Germany Sartoriu),JXFSTPRP-24 研磨仪(上海净信科技有限公司),色谱柱:ACQUITY UPLC BEH C18(American Waters),明澈D24 UV 纯水仪(Merck Millipore),YM-080S超声仪(深圳方奥微电子有限公司),乙腈、甲酸、甲醇(色谱纯,CNW Technologies,Germany),L-2-氯苯丙氨酸(分析纯,上海恒柏生物科技有限公司)。
1.2 试验方法
1.2.1 代谢物提取
将3 个产地的黄芪根样品进行研磨(60 Hz,1 min);称取100 mg 样本,加2 个小钢珠,加入500 μL 提取液(甲醇∶水=4∶1);涡旋30 s 后45 Hz/240 s 研磨,冰水浴超声1 h;-40 ℃静置1 h 后将样本4 ℃,12 000 r/min(离心力13 800×g,半径8.6 cm)离心15 min;取出上清过0.22 μm 滤膜至进样瓶中上机检测;所有样本取30 μL 上清混合成QC 样本上机。
1.2.2 色谱条件
采用Waters 的UPLC BEH C18色谱柱(100 mm×2.1 mm,1.7 μm),柱温45 ℃,流速0.4 mL/min,进样体积为5 μL。流动相A 是水(含0.1% 甲酸),B 是乙腈(含0.1% 甲酸),梯度洗脫条件为0~3.5 min,95%~85%A;3.5~6.0 min,85%~70%A;6.0~6.5 min,70%~70%A;6.5~12.0 min,70%~30%A;12.0~12.5 min,30%~30%A;12.5~18.0 min,30%~0%A;18.0~25.0 min,0%~0%A ;25.0~26.0 min ,0%~95%A ;26.0~30.0 min,95%~95%A。
1.2.3 质谱条件
利用Q Exactive Focus 质谱仪,在控制软件(Xcalibur,Thermo Fisher Scientific)控制下,采用FullScan-ddMS2 功能进行一级和二级质谱数据的采集。详细参数如下:鞘气流量为45 L/min;辅助气体流量为15 L/min;毛细管温度为400 ℃;总质谱分辨率为70 000;二级质谱分辨率为17 500;碰撞能量为15 eV/30 eV/45 eV(负离子模式),喷淋电压为4.0 kV(正离子)或-3.6 kV(负离子)。
1.3 数据处理
将质谱原始数据导入XCMS 软件,进行保留时间校正、峰鉴定、峰提取、峰积分、峰对齐等,采用派森诺公司自建中药数据库及相应匹配方法对含有二级质谱数据的峰进行鉴定。采用主成分分析(PCA)和正交偏最小二乘判别分析(OPLS-DA)等多元统计分析来区分组间代谢的总体差异,筛选差异代谢物。利用MetaboAnalyst 软件依据KEGG数据库对差异代谢物进行途径富集分析,以获得代谢途径富集结果,利用Matlab 绘制火柴杆图。
2结果与分析
2.1 不同产地黄芪的非靶向代谢组学分析
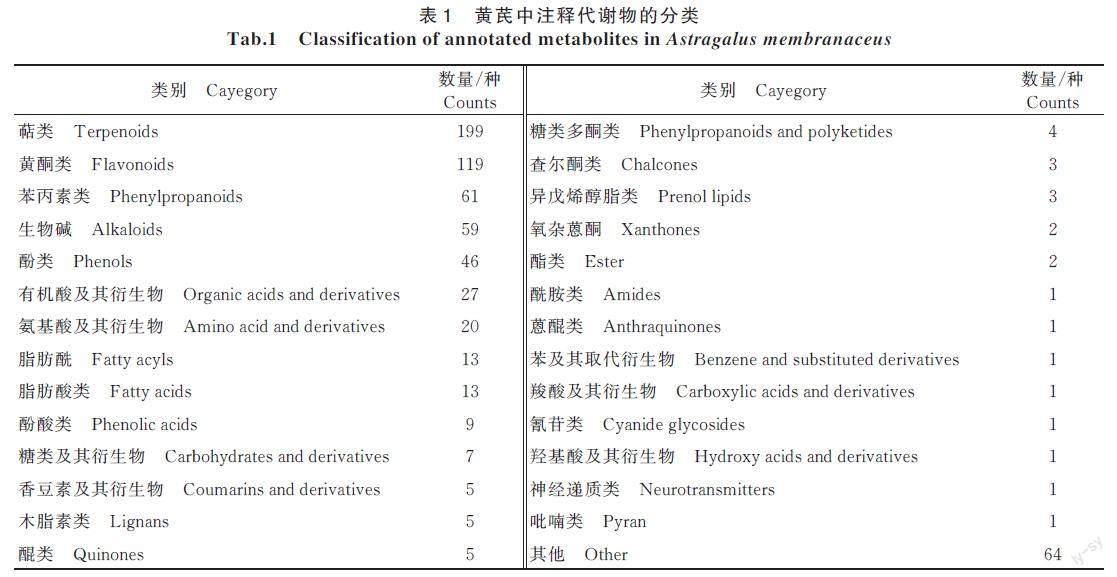
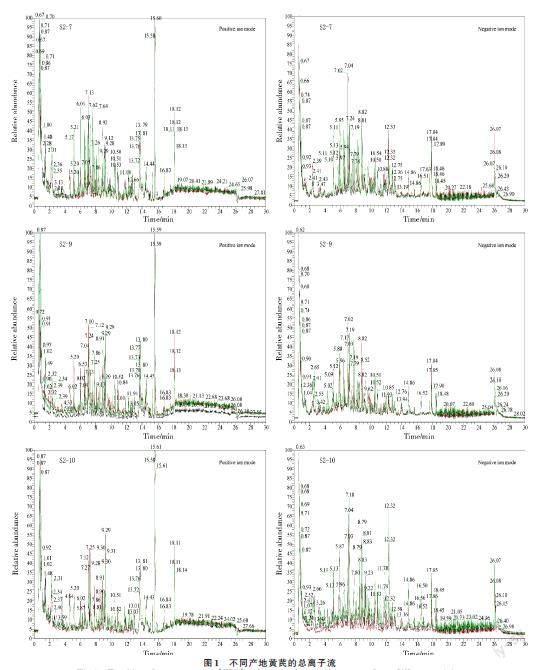
依据来源,将S2-7 和S2-9 分为山西省黄芪资源(SXZY)组,S2-10 为甘肃省黄芪资源组。为深入了解不同产地黄芪之间药用品质的差异,对3 种不同产地黄芪进行了LC-MS/MS 非靶向代谢组学测定,得到总离子流图(图1)。如图1 所示,正负离子模式下的TIC 重叠度均较高,所检测到物质峰都比较丰富,且3 种不同种质黄芪总离子流图变化趋势存在差异,如保留时间在4~12 min,不同种质出现的峰对应的物质种类和相对含量明显不同,此研究结果为后续统计学分析差异性代谢物奠定基础。LC-MS/MS 原始数据预处理后,采用测定公司自建中药数据库进行检索匹配,共检测出674 种代谢物,分为29 类,其中,萜类物质最多,为199 种;黄酮类物质位居第2,有119 种;苯丙素类物质位居第3,有61 种(表1)。
2.2 不同产地黄芪的代谢物判别分析
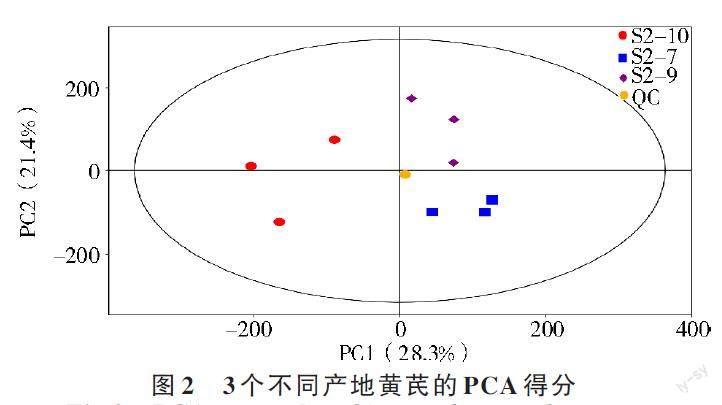
为理清S2-7、S2-9 和S2-10 之间化学成分的不同,采用多元统计法对其代谢物进行分析。首先利用SIMCA 软件将处理后的质谱数据进行PCA分析,观察各样本间的总体分布和整个分析过程的稳定性,得到3 种不同产地黄芪的PCA 得分图(图2)。由图2 可知,3 种不同产地黄芪的数据点在得分图上可以明显区分,S2-10 位于第1、4 象限,S2-7位于第3 象限,S2-9 位于第2 象限,表明3 种不同产地黄芪样品中的代谢物存在明显差异。
将3 种不同产地黄芪分为3 组S2-9 vs S2-7、S2-10 vs S2-7、S2-10 vs S2-9;同时,将甘肃省黄芪与山西省黄芪进行比较,分为S2-10 vs SXZY组。采用OPLS-DA 进一步对S2-9 vs S2-7、S2-10 vs S2-7、S2-10 vs S2-9 和S2-10 vs SXZY 4 组数据进行分析,由图3-A-D 可知,4 组样品分布在置信区间的左右两侧,判别效果明显,显示了4 组数据之间的显著差异。对4 组模型进行置换验证,由图3-E-G 可知,置换检验随机模型的Q2 值均小于原模型的Q2 值,图3-H 的置换检验随机模型的Q2值不小于原模型的Q2 值,但与纵轴负半轴相交,模型成立。综上表明,这4 组原始模型具有良好的稳定性,不存在过拟合现象,可用于进一步筛选差异代谢物。
2.3 不同产地黄芪的差异代谢物筛选与鉴别
2.3.1 甘肃与山西黄芪的差异代谢物筛选与鉴别
采用多维分析和单维分析相结合的方法,依据Student?s t-test 的P<0.05,且OPLS-DA 的VIP>1 筛选组间差异代谢物。由图4 可知,甘肃省黄芪(S2-10)与山西省黄芪(SXZY)共检测到81 种差异代谢物,其中,43 种代谢物在S2-10 中丰度高,38 种代谢物在SXZY 组中丰度高。进一步对差异代谢物的log2(FC)值进行排序,筛选上调和下调前15 种代谢物(图5),在甘肃黄芪中上调幅度前15 的差异代谢物包含:萜类4 种,生物碱、黄酮类、苯丙素类均为2 种,三萜类、有机酸及其衍生物、氨基酸衍生物、酚类和其他类物质均为1 种;下调幅度前15 的差异代谢物有2 种萜类,2 种氨基酸衍生物,2 种黄酮类,苯丙素类、环烯醚萜类、糖类多酮类、糖类及其衍生物、有机氧化合物均为1 种,以及4 种其他类物质。
综上可见,S2-10 与SXZY 的主要差异代谢物为萜类、黄酮类和苯丙素类物质,其中,差异倍数最大的代谢物为萜类物质Oleanane-2H,+1O,1COOH,O-HexA-HexA,其差异倍数为115 倍,这些差异代谢物与甘肃和山西黄芪之间的药材品质差异有关。
2.3.2 五台与浑源黄芪的差异代谢物筛选与鉴别
为阐明五台黄芪(S2-9)和浑源黄芪(S2-7)之间的药材品质差异,同2.3.1 对S2-9 和S2-7 进行组间分析,根据VIP 值和P 值(VIP>1,P<0.05)筛选到67 种差异代谢物,其中,32 种代谢物在S2-9 中丰度高,35 种代谢物在S2-7 中丰度高(图6)。进一步对差异代谢物的log2(FC)值排序,列出差异倍数前15(上调和下调)的代谢物(图7),S2-9 与S2-7相比,上调的差异代谢物有4 种黄酮类,3 种苯丙素类,2 种生物碱,萜类、酚酸类、异戊烯醇脂类、蒽醌类均为1 种,其他类物质2 种;下调的差异代谢物有11 种萜类,脂肪酸类、酯类、异戊烯醇脂类和其他类物质均为1 种。
综上可见,S2-9 的多数黄酮类、苯丙素类和生物碱类物质高于S2-7(1≤log2FC≤3),S2-7 的多数萜类物质高于S2-9(1≤-log2FC≤3),这些代谢物的差异可能是山西五台与浑源黄芪药材品质差异的物质基础。
2.4 差异代谢物通路分析
分别将S2-10 vs SXZY 组的81 个差异代谢物及S2-9 vs S2-7 组的67 个差异代谢物在KEGG 数据库映射后得到的通路进行富集分析[18] (图8),S2-10 vs SXZY 组共富集到8 条通路,有3 条差异显著的通路(P<0.05),分别是异喹啉生物碱生物合成、吲哚生物碱生物合成和烟酸盐和烟酰胺代谢,其差异代谢物为多巴胺、色胺和烟酸,且都作用于植物抗逆性与生长调节,推測与不同产地的气候条件和土壤条件相关;S2-9 vs S2-7 组富集到8 条通路,无差异显著通路,说明山西省内两组黄芪种质在种源上差异不大,可能与早期各地引种导致种质混杂有关[19],综上可见,来源于不同省份的黄芪亲缘关系较远。
3结论与讨论
本研究基于液相色谱-串联质谱技术对3 种不同产地黄芪进行了非靶向代谢组学分析,共检测出674 种代谢物,分为29 类,3 个产地的黄芪均含有较多的萜类、黄酮类和苯丙素类化合物。随后对质谱数据进行PCA 分析,发现3 个种质可以明显区分,由于PCA 是一种无监督的分析方法,环境和系统误差都会对结果造成影响。为排除因试验无关的某些因素引起的代谢变化,同时理清不同种质两两之间的对比,将3 个种质分为4 组,进行可以过滤掉代谢物中与分类变量不相关的正交变量,并对非正交变量和正交变量分别分析的OPLS-DA 分析[21],4 组数据皆区分明显,且数据稳定。综合2 种分析方法表明,不同产地黄芪的代谢物在类型和含量上具有一定的差异。
甘肃黄芪(S2-10)与山西黄芪(S2-9、S2-7)组比较,共筛选出81 种差异代谢物;进一步筛选差异前15 的代谢物,发现甘肃黄芪与山西黄芪的主要差异代谢物为萜类、黄酮类和苯丙素类物质。戴瑜婷等[22]研究表明,不同产地黄芪的总皂苷、总黄酮、总多糖的含量也不同,可以作为黄芪质量标志物预测分析指标。本研究中上调第一的萜类物质Oleanane-2H, +1O, 1COOH, O-HexA-HexA 主要存在于S2-10 中,通过Mass Bank 检索发现,Oleanane-2H, +1O, 1COOH, O-HexA-HexA 是一种三萜皂苷类物质,可进一步对该物质进行探索,以期找到可鉴别甘肃黄芪(S2-10)与山西黄芪(S2-7、S2-9)的潜在标志性物质,为甘肃黄芪的应用奠定基础。山西省内两产区五台与浑源黄芪组,共筛选出67 种差异代谢物,进一步发现五台黄芪(S2-9)中黄酮类、苯丙素类和生物碱类成分显著高于浑源黄芪(S2-7),而浑源黄芪中的萜类成分显著高于五台黄芪。表明即使是山西产黄芪,因分布区地理位置、土壤、温度等生态条件不同,也会影响黄芪体内代谢物的合成。但本研究中根据差异倍数,没有发现可作为区分浑源黄芪(S2-7)与五台黄芪(S2-9)的特征代谢物,后续可重复试验进一步验证,以期为区分不同道地产区黄芪提供思路。
对差异代谢物进行富集通路分析,S2-10 vsSXZY 组富集到3 条差异显著通路,其中,异喹啉生物碱生物合成为最主要的代谢途径,该通路从2 种酪氨酸衍生物的缩合开始,经一系列反应形成反式心果碱,并伴随产生多巴胺等物质。研究发现,该通路富集分析中多巴胺含量上调,推测反式心果碱含量也相应增加,而反式心果碱是大部分生物碱的前体物质[23],因而甘肃黄芪(S2-10)的生物碱类物质合成随之增多,这与通路富集分析结果一致。生物碱是一类应对生境胁迫的次生代谢产物,具有防御作用[24],与生境胁迫耐受性相关。另外,在烟酸盐和烟酰胺代谢通路中,烟酸含量上调,推测烟酰胺含量也得到提高,而烟酰胺与植物抗逆相关,是真核生物对逆境反应应答而产生的一种信号物质[25]。由上分析,来源于不同省份的黄芪亲缘关系较远。同时,甘肃黄芪(S2-10)生物碱类物质含量较多,可能与其应对生境变化而提高抗逆性密不可分,后续可将本研究结果进一步应用于黄芪抗逆性等研究。
综上可见,甘肃与山西黄芪具有较远的亲缘关系,与实验室前期对80 份不同黄芪种质的遗传多样性研究结果一致[17],说明利用非靶向代谢组学分析不同种质黄芪方法可行;而山西五台与浑源黄芪的亲缘关系较近,但在代谢物上也存在差异,可进一步研究应用于临床药理药效分析。但本研究中甘肃、山西五台和浑源黄芪均引种到山西汾阳,不排除生境改变引起的差异,仍需收集大量样品进行试验。
本研究采用非靶向代谢组学技术,从PCA、OPLS-DA 等多元统计分析、差异代谢通路KEGG富集分析等方面进行了系统研究,发现不同产地黄芪次级代谢物存在差异。同时,富集通路分析发现,S2-10 vs SXZY 组富集到3 条差异显著通路,而S2-9 vs S2-7 组没有差异显著通路,进一步证明不同省份黄芪之间亲缘关系较远,该研究为评价不同产地黄芪品质提供新思路,并且环境因素对植物代谢产物的影响值得进一步深入研究。
参考文献:
[1] 国家药典委员会. 中华人民共和国药典[S]. 北京:中国医药科技出版社,2020:1088.
Chinese Pharmacopoeia Commission. Pharmacopoeia of thePeople?s Republic of China[S]. Beijing:China Medical SciencePress,2020:1088.
[2] WU X T,LI X T,WANG W,et al. Integrated metabolomicsand transcriptomics study of traditional herb Astragalus membra?naceus Bge. var. mongolicus(Bge.)Hsiao reveals global meta?bolic profile and novel phytochemical ingredients[J]. BMC Ge?nomics,2020,21(Suppl 10):697.
[3] 吴宏辉,李红丽,侯俊玲,等. 黄芪种质资源研究进展[J]. 中医药导报,2016,22(24):76-79.
WU H H,LI H L,HOU J L,et al. Research progress on germ ?plasm resources of huangq(i Radix astragali)[J]. Guiding Journalof Traditional Chinese Medicine and Pharmacy,2016,22(24):76-79.
[4] 冯学金,刘根科,梁素明. 蒙古黄芪种质资源研究进展[J]. 山西农业科学,2010,38(8):95-98.
FENG X J,LIU G K,LIANG S M. Review of studies on germ ?plasm resources of Mongolia milkvetch[J]. Journal of Shanxi Ag?ricultural Sciences,2010,38(8):95-98.
[5] CHO W C S,LEUNG K N. In vitro and in vivo immunomodu?lating and immunorestorative effects of Astragalus membrana?ceus[J]. Journal of Ethnopharmacology,2007,113(1):132-141.
[6] KUO Y H,TSAI W J,LOKE S H,et al. Astragalus membrana?ceus flavonoids(AMF) ameliorate chronic fatigue syndrome in?duced by food intake restriction plus forced swimming[J]. Jour?nal of Ethnopharmacology,2009,122(1):28-34.
[7] TSENG A,YANG C H,CHEN C H,et al. An in vivo molecu?lar response analysis of colorectal cancer treated with Astragalusmembranaceus extract[J]. Oncology Reports,2016,35(2):659-668.
[8] REN S,ZHANG H,MU Y P,et al. Pharmacological effects ofAstragaloside IV:a literature review[J]. Journal of TraditionalChinese Medicine,2013,33(3):413-416.
[9] 张莹,李文倩,胡瑜辉,等. 黄芪营养吸收规律及其与产品品质形成的关系[J]. 山西农业科学,2020,48(4):540-546.
ZHANG Y,LI W Q,HU Y H,et al. Nutritional absorption lawof Astragalus membranaceus and its relation with product qualityformation[J]. Journal of Shanxi Agricultural Sciences,2020,48(4):540-546.
[10] 孟益德,吕庚鑫,刘攀峰,等. 不同产地杜仲雄花的非靶向代谢组学比较与分析[J]. 中南林业科技大学学报,2022,42(1):156-166.
MENG Y D,LYU G X,LIU P F,et al. Comparison and analy?sis of untargeted metabolomics of male flowers of Eucommiaulmoides from different origins[J]. Journal of Central SouthUniversity of Forestry & Technology,2022,42(1):156-166.
[11] 霍冬敖,田瑞丰,任永权,等. 基于UPLC-MS/MS 技术的野生及栽培韭菜籽的代谢组学研究[J]. 广西植物,2022,42(12):1995-2006.
HUO D A,TIAN R F,REN Y Q,et al. UPLC-MS/MS-basedmetabolomic characterization and contrastive analysis betweenAllium wallichii and A.tuberosum seeds[J]. Guihaia,2022,42(12):1995-2006.
[12] LYU X R,YAN K M,CHEN W J,et al. The characterizationof metabolites alterations in white adipose tissue of diabetic GKRats after ileal transposition surgery by an untargeted metabolo?mics approach[J]. Adipocyte,2021,10(1):275-284.
[13] JOHNSON C H,IVANISEVIC J,SIUZDAK G. Metabolo?mics:beyond biomarkers and towards mechanisms[J]. NatureReviews Molecular Cell Biology,2016,17:451-459.
[14] 徐天润,刘心昱,许国旺. 基于液相色谱-质谱联用技术的代谢组学分析方法研究进展[J]. 分析测试学报,2020,39(1):10-18.
XU T R,LIU X Y,XU G W. Advances of analytical methodsfor liquid chromatography-mass spectrometry-based metabolo?mics[J]. Journal of Instrumental Analysis,2020,39(1):10-18.
[15] WANG Y M,LIU L,MA Y K,et al. Chemical discriminationof Astragalus mongholicus and Astragalus membranaceusbased on metabolomics using UHPLC-ESI-Q-TOF-MS/MSapproach[J]. Molecules,2019,24(22):4064.
[16] 牛媛婧,温嘉琪,姬惠鑫,等. 基于超高效液相色谱串联高分辨质谱技术的不同产地党参的非靶向代谢组学研究[J]. 药学学报,2023,58(7):1842-1850.
NIU Y J,WEN J Q,JI H X,et al. Study on untargeted metabo?lomics of Codonopsis pilosula from different producing areasbased on ultra-performance liquid chromatography tandem highresolution mass spectrometry[J]. Acta Pharmaceutica Sinica,2023,58(7):1842-1850.
[17] 牛景萍,石志勇,田洪岭,等. 基于GBS 简化基因组测序的不同黄芪种质资源遗传多样性分析[J]. 分子植物育种,2022,20(24):8291-8298.
NIU J P,SHI Z Y,TIAN H L,et al. Genetic diversity analysisof different Astragalus membranaceus germplasm resources bygenotyping-by-sequencing technology[J]. Molecular Plant Breeding,2022,20(24):8291-8298.
[18] 牟红梅,慈志娟,艾沙江·买买提,等. 基于非靶向代谢组学的茄梨和红茄梨成熟期果皮代谢产物的差异分析[J]. 色谱,2021,39(11):1203-1212.
MOU HM,CI Z J,MAMAT A,et al. Analysis of metabolitedifferences in skin between Clapp?s Favorite and its mutantRed Clapp?s Favorite through non-targeted metabolomics[J].Chinese Journal of Chromatography,2021,39(11):1203-1212.
[19] 胡艳改,杜宇忠,郝磊,等. 中药材蒙古黄芪与膜荚黄芪优质种源的研究[J]. 山西农业科学,2017,45(10):1662-1666,1690.
HU Y G,DU Y Z,HAO L,et al. Study on high quality prov?enances of Astragalus membranaceus(fisch.)bge. var. mongho?licus(bge.) Hsiao and Astragalus. membranaceus(fisch.) bge[J]. Journal of Shanxi Agricultural Sciences,2017,45(10):1662-1666,1690.
[20] TRYGG J,WOLD S. Orthogonal projections to latent struc?tures(O-PLS)[J]. J ournal of Chemometrics,2002,16(3):119-128.
[21] 娄港归,夏杰,杨健,等. 基于UPLC-Q-TOF-MS/MS 代谢组学的铁皮石斛和玫瑰石斛化学成分差异研究[J]. 药学学报,2021,56(12):3331-3344.
LOU G G,XIA J,YANG J,et al. Differences in the chemicalcomposition of Dendrobium officinale Kimura et Migo andDendrobium crepidatum Lindl based on UPLC-Q-TOF-MS/MS and metabolomics[J]. Acta Pharmaceutica Sinica,2021,56(12):3331-3344.
[22] 戴瑜婷,张雪燕,王艺璇,等. 黄芪的现代研究进展及其质量标志物的预测分析[J]. 中国中药杂志,2022,47(7):1754-1764.
DAI Y T,ZHANG X Y,WANG Y X,et al. Research prog?ress on Astragali Radix and prediction of its quality markers(Q-markers)[J]. China Journal of Chinese Materia Medica,2022,47(7):1754-1764.
[23] 高世虎,徐辉筠,王华光,等. 生物碱对动植物生长促进作用的研究进展[J]. 江苏林业科技,2013,40(2):39-42.
GAO S H,XU H Y,WANG H G,et al .Research progress ofthe growth promoting effect of alkaloids[J]. Journal of JiangsuForestry Science & Technology,2013,40(2):39-42.
[24] 张贵友,朱瑞宇,戴尧仁. 烟酰胺诱导的植物细胞凋亡及其机理研究[C]//中国植物学会七十周年年会论文摘要汇编(1933—2003). 成都:中国植物学会,2003:447.
ZHANG G Y,ZHU R Y,DAI Y R. Nicotinamide-inducedapoptosis in plant cells[C]//Compilation of abstracts of the70th anniversary annual meeting of the Botanical Society ofChina.Chengdu:Botanical Society of China,2003:447.
[25] 覃美,肖京林,王曉霞,等. 烟酰胺对甘蔗锰毒的缓解效应[J].热带作物学报,2021,42(11):3339-3344.
QIN M,XIAO J L,WANG X X,et al. Mitigation effect ofnicotinamide on manganese toxicity in sugarcane[J]. ChineseJournal of Tropical Crops,2021,42(11):3339-3344.
猜你喜欢
现代临床医学(2022年4期)2022-09-29
国际口腔医学杂志(2019年3期)2019-05-31
天然产物研究与开发(2018年2期)2018-04-04
当代化工研究(2016年5期)2016-03-20
分析测试学报(2015年7期)2016-01-13
医学研究杂志(2015年11期)2015-06-10
质谱学报(2015年5期)2015-03-01
天然产物研究与开发(2014年6期)2014-04-27
西安工程大学学报(2014年2期)2014-02-28
西南军医(2014年4期)2014-01-19
