
油茶籽粕发酵菌株的筛选鉴定及发酵产物特性分析
2024-05-03 15:58朱晓丽赵锦绣李富松张星柯玉鑫申烨华张子夜王军强
西北大学学报(自然科学版) 2024年2期
朱晓丽 赵锦绣 李富松 张星 柯玉鑫 申烨华 张子夜 王军强


摘要 油茶行業的迅速发展导致油茶籽粕废弃物的大量产生,其落后的处理方式造成了严重的资源浪费,制约了我国油茶行业的可持续发展。油茶籽粕中富含有机质,利用微生物发酵法降解其中的纤维素、木质素及茶皂素等生物大分子来制备有机肥是实现油茶籽粕合理资源化利用的有效途径。通过筛选获得3株针对油茶籽粕中纤维素和木质素的高效降解菌,包括1株真菌X1-1,属于Pleurostomarichardsiae,以及2株细菌M4-2和M5-2,分别为蒙氏假单胞菌(Pseudomonas monteilii)和类芽孢杆菌(Paenibacillussp)属,可有效降低发酵产物中茶皂素的含量,实现油茶籽粕的无害化处理。实验结果显示,经菌株X1-1、M4-2和M5-2发酵处理28 d后,油茶籽粕中纤维素含量的降解率分别为29.74%、19.65%和13.19%,木质素含量的降解率分别为12.84%、17.85%和26.39%,茶皂素含量的降解率分别为34.06%、45.05%和46.15%;且发酵后的油茶籽粕腐熟度、有机质含量和养分含量等指标均符合有机肥标准。研究结果拓展了油茶籽粕的资源化利用途径,同时为木质素和纤维素的高效降解提供了良好的菌种资源。
关键词 油茶籽粕;纤维素;木质素;茶皂素;降解菌;有机肥
Screening and characterization of fermentation strains of camellia
sinensis seed meal and fermentation products
Abstract The rapid development of camellia oleifera industry has led to a large number of camellia oleifera seed residue, which caused serious waste of resources, restricting the sustainable development of Chinas camellia oleifera industry.Considering the high contents of organic matter, using camellia oleifera seed residue to prepare organic fertilizer is an effective way for resource utilization. In the process of composting, cellulose, lignin and tea saponin and other biological macromolecules should be effectively biodegraded by microorganisms. In this paper, three highly efficient degrading strains targeting cellulose and lignin in camellia oleifera seed residue were obtained by screening and domestication, including 1 strain of fungus X1-1(Pleurostomarichardsiae), and 2 strains of bacteria M4-2 (Pseudomonas monteilii) and M5-2 (Paenibacillus sp.).The three strains can also effectively reduce the content of tea saponin in fermentation products, to realize the harmless treatment of camellia oleifera seed residue.The experimental results showed that after 28 days of fermentation with strain X1-1, M4-2 and M5-2, the degradation rates of cellulose content in camellia oleifera seed residue were 29.74%, 19.65% and 13.19%, the degradation rates of lignin content were 12.84%, 17.85% and 26.39%, and the degradation rates of tea saponin content were 34.06%, 45.05% and 46.15%, respectively.Moreover, the maturity, organic matter content, andnutrient content in fermentation product of camellia oleifera seed residue all met the organic fertilizer standards.The results of this study expand the resource utilization path of camellia oleifera seed residue, and provide good strain resources for the efficient degradation of lignin and cellulose.
Keywords camellia oleifera seed residue; cellulose; lignin; tea saponins; fermentation strains; organic fertilizer
作为我国特有的食用木本油料作物,油茶在湖南、江西等地广泛分布,种植面积高达400多万亩,年产油茶籽超过100多万吨[1-2]。自2016年油茶正式被列为我国大宗油料作物以来,其产量及产值逐年稳步增长[2],与此同时,油茶行业的快速发展也产生了大量的油茶籽粕废弃物,并造成了严重的资源浪费。据估算,我国每年的油茶籽粕废弃物产量超过70万吨,而其高值化利用技术的发展则严重滞后,极大地制约了油茶行业的健康可持续发展[3-4]。
油茶籽粕废弃物中富含丰富的营养物质,包括蛋白质、糖类和多酚等[5],可广泛应用于医药、食品和饲料等领域[6]。然而,从我国目前的油茶籽粕废弃总量和发展趋势考虑,现有的废弃物利用方式均难以满足油茶行业的快速发展。相较之下,以油茶籽粕为底物进行发酵处理制备有机肥,是实现油茶籽粕大规模资源化利用的有效处理方式[7]。油茶籽粕中富含大分子物质木质素和纤维素,木质素是以苯基丙烷为结构单元的一种高分子聚合物[8],纤维素是以β-D-葡萄糖为结构单元,通过β-1, 4-糖苷键连接的一种线性高分子多糖化合物[9]。这些大分子物质不能被作物直接吸收,在自然界中降解缓慢[10],因此在使用油茶籽粕制备有机肥的过程中,需要通过微生物降解将其转化为能够被作物吸收利用的小分子营养物质。研究发现,白腐菌、褐腐菌、软腐菌、木霉属、青霉属、曲霉属等真菌均对木质素和纤维素表现出较强的降解性能,其中,部分菌株对木质素、纤维素的降解率可达20%以上[11-14]。有研究显示,通过添加地衣芽孢杆菌、酿酒酵母及植物乳杆菌对油茶籽粕进行发酵,大部分粗纤维中的高分子芳香化合物分解为低分子物质,有效破坏了木质纤维素分子结构,将部分糖类物质转化为菌体蛋白,从而使得粗纤维含量降低,粗蛋白的含量升高[15-17],因此制备的有机肥的肥力也明显提升。除木质素、纤维素外,油茶籽粕中还含有较多的茶皂素,作为一种糖甙化合物,茶皂素对多种微生物的生长具有显著抑制作用[18-19],因此不仅会阻碍油茶籽粕固态发酵的进程,在进入土壤后也会对土壤益生菌产生不利影响[20]。此外,茶皂素随有机肥施入土壤后,还会对作物种子发芽产生抑制作用,同时减缓土壤有机质的分解,阻碍土壤营养物质循環过程,降低土壤肥力[21]。然而,现有关于油茶籽粕发酵的研究中大多关注木质素、纤维素的降解以及养分物质的变化,而忽略了油茶籽粕中茶皂素的降解,难以获得腐熟程度理想的有机肥产物。
由此可知,以油茶籽粕发酵生产有机肥,需要综合考虑木质素、纤维素和茶皂素的降解,以实现对油茶籽粕的脱毒处理。因此,本研究通过筛选获得可高效降解油茶籽粕中纤维素、木质素及茶皂素的功能菌株,评价了功能菌株对纤维素、木质素和茶皂素的降解效果,将其用于油茶籽粕发酵制备有机肥,获得了腐熟度、有机质含量、养分和重金属含量等指标均符合标准的有机肥,为油茶籽粕废弃物的大规模资源化利用提供了良好的思路和可行的方法。
1 材料与方法
1.1 实验材料
1.1.1 原料及试剂
本实验所用油茶籽粕原料购自湖南大三湘茶油资源有限公司。葡萄糖、磷酸二氢钾、磷酸氢二钠、氯化钠、磷酸氢二钾、羧甲基纤维素钠硫酸铵和七水合硫酸镁均为分析纯。酵母浸粉和蛋白胨均为BR生物试剂。
马铃薯液体(PDA)培养基:将200 g马铃薯切成小块煮沸20 min,用8层纱布过筛,冷却后加入20.00 g葡萄糖,加蒸馏水定容至1 000 mL,灭菌处理。
木质素初筛液体培养基:碱性木质素2.00 g、硫酸铵 1.00 g、七水合硫酸镁 0.50 g、磷酸二氢钾 1.00 g、磷酸氢二钠 0.20 g溶于蒸馏水,pH调至7.00,蒸馏水定容至1 000 mL,灭菌处理。
苯胺蓝显色培养基:酵母浸粉5.00 g、氯化钠 10.00 g、蛋白胨10.00 g、琼脂20.00 g、苯胺蓝0.25 g、蒸馏水1 000 mL,pH调至7.00~7.20[22],灭菌处理。
刚果红纤维素培养基:刚果红0.20 g、 微晶纤维素1.88 g、 七水合硫酸镁0.25 g、 磷酸氢二钾0.50 g、 琼脂20.00 g、 去离子水1 000 mL, 灭菌处理。
羧甲基纤维素钠(CMC-Na)液体培养基:硫酸铵1.00 g、氯化钙0.02 g、七水合硫酸镁0.20 g、羧甲基纤维素钠(CMC-Na) 10.00 g、六水合氯化铁0.05 g、磷酸氢二钾1.00 g、茶皂素5.00 g、琼脂20.00 g、去离子水1 000 mL[23-24],灭菌处理。
LB培养基:氯化钠10.00 g、蛋白胨10.00 g、酵母提取物5.00 g、蒸馏水1 000 mL,灭菌处理。
固体培养基:在其对应的液体培养基中加入20 g琼脂,灭菌处理。
1.1.2 试剂配制
中性洗涤剂:称取乙二胺四乙酸(EDTA)18.6 g 和四硼酸钠(Na2B4O7·10H2O)6.8 g,加一定量的去离子水适当加热,再加乙二醇乙醚10 mL后加30 g十二烷基硫酸钠(SDS);用天平量取 4.65 g磷酸氢二钠放入到另一个三角瓶中,加适量去离子水后溶解,可适当加热,待溶液温度降至室温后,将配好的2种溶液放入1 000 mL容量瓶中用去离子水定容到刻度线。此溶液pH值应在6.9~7.1。
酸性洗涤剂:移液枪抽取 27.87 mL 98%的浓硫酸用玻璃棒引流缓慢滴加500 mL 去离子水中,待温度稳定后,向体系内加入20 g十六烷基三甲基溴化铵(CTAB)拌混匀, 待液体呈现透明并带有少量气泡证明溶解完全, 将溶液定容到1 000 mL即可,备用。
72%的硫酸:量取质量分数为 98.3%的浓硫酸 398 mL, 用玻璃棒引流至去离子水中, 定容至1 000 mL即可。
8%(w/v)香草醛无水乙醇溶液:准确称取8.0 g香草醛,用无水乙醇溶解,置于100 mL容量瓶中定容。为使测定的数据具有良好的重现性和稳定性,每次进行样品分析时,都需新配制香草醛无水乙醇溶液。同时,反应过程中,需振摇容量瓶3次,使反应均匀。
77%(v/v)硫酸:取硫酸(98%w/w) 77 mL加到23 mL去离子水中。
1.2 油茶籽粕理化性质测定
油茶籽粕样品的水分、灰分、总酸、粗纤维、粗脂肪和茶皂素的含量测定分别参照《食品安全国家标准食品中水分的测定》(GB 5009.3—2016)[25]、《食品安全国家标准食品中灰分的测定》(GB 5009.4—2016)[26]、《食品中总酸的测定》(GB/T 12456—2008)[27]、《植物类食品中粗纤维的测定》(GB/T 5009.10—2003)[28]、《食品安全国家标准食品中脂肪的测定》(GB 5009.6—2016)[29]和《油茶籽粕中茶皂素的提取工艺研究》[30]的规定方法。
1.3 菌株的筛选
1.3.1 纤维素降解菌株的筛选
取油茶籽粕样品10.00 g,加入少许无菌水,发酵7 d后转入100 mL的PDA液体培养基,在28 ℃、120 r/min条件下培养,待长出大量菌体后,一部分转入纤维素初筛液体培养基用于纤维素降解菌的筛选,另一部分转入木质素初筛液体培养基用于木质素降解菌的筛选。转入羧甲基纤维素钠(CMC-Na)液体初筛培养基中的菌体在摇床继续培养2 d后,将菌液进行10倍梯度的稀释,得到稀释度为
10-1~10-6的培养液。随后,取不同浓度的菌液各100 μL,通过平板涂布法接种到刚果红纤维素固体培养基上,并在28 ℃、120 r/min条件下培养3~5 d。观察菌落生长情况,当出现有透明圈的菌落即初步认为菌株具备纤维素降解能力,选取其中透明圈直径较大的菌落进行多次划线并分离纯化。将初筛得到的单菌落挑取在CMC-Na液体培养基中,孵育2 d后进行稀释,并采用稀释涂布法进一步在羧甲基纤维素钠(CMC-Na)固体培养基上纯化纤维素降解菌,挑选出长势良好的菌株。将复筛选出的菌株接种至LB培养基中培养1 d,成为菌种活化液,将活化液与50%的甘油混合,冻存至-80 ℃冰箱中待用。
1.3.2 木质素降解菌株的筛选
转入木质素液体初筛培养基的菌体,在摇床中保持28 ℃、120 r/min条件下培养2 d。之后,同样进行10倍梯度稀释处理,得到10-1~10-6稀释度的培养液。随后将不同浓度梯度的菌液各取100 μL用平板涂布法接种到木质素初筛固体培养基上,并在28 ℃、120 r/min条件下培养3~5 d。选择木质素初筛固体培养基上长出的单独菌落,挑到木质素液体培养基中培养3 d之后,取菌液稀释至10-5后在苯胺蓝显色培养基上进行不同浓度梯度的菌液涂布,筛选出变色圈较大或者使培养基完全变色的菌株。将筛选出的菌株挑到LB培养基中培养1 d,成为菌种活化液,将活化液与50%的甘油保存,冻存至-80 ℃冰箱。
1.4 菌株鉴定
1.4.1 显微形态特征
霉菌采用水浸片法观察细胞形态,细菌采用革兰氏染色法染色后镜检。
1.4.2 ITS、16S rDNA 序列鉴定
筛选出的菌株送至上海凌恩生物科技有限公司进行ITS、16S rDNA测序。具体过程为:以提取的菌株总DNA为模板进行ITS、16S rDNA基因PCR扩增、测序,X1-1、M4-2和M5-2分别获得556bp、1422bp和1446bp的PCR扩增片段,GenBank数据库登录号分别为OL677875、OL855974和OL872184。将测序得到的菌株ITS、16S rDNA与Genbank数据库进行Blast比对,采用MEGA软件对比对结果进行分析,将目标菌株鉴定到属,并绘制系统发育树分析菌株种属特性。
1.5 降解实验
1.5.1 纯木质素降解实验
分别配制浓度为0.00、0.10、0.20、0.30、0.40和0.50 g/L的碱性木质素水溶液,测定其OD280值以制作木质素标准吸收曲线,所得回归方程为Y=4.234 9X-0.01,R2=0.999 5。
将菌液(OD=1)按30%(v/m)的接種量接入100 mL木质素液体培养基中, 并在28 ℃、120 r/min条件下振荡培养。使用5.00 mL无菌EP管每隔7 d取1次样品。取样时摇匀培养基,每次吸取4.00 mL液体,于8 000 r/min条件下离心10 min,所得上清液用0.45 μm的水系滤膜过滤,测定其OD280浓度为0.40 g/L的值,所得结果代入回归方程计算碱性木质素降解率,
式中: C1表示初始培养基中碱性木质素浓度, g/L;C2表示待测样品中碱性木质素浓度, g/L。
1.5.2 油茶籽粕中木质素、纤维素的含量测定
参照范式粗纤维分析法[31]测定油茶籽粕发酵过程中的纤维素和木质素含量。具体方法为:将发酵后的油茶籽粕于105 ℃烘干,取烘干样1 g于直筒烧杯,加入100 mL中性洗涤后,剩余的残渣烘干,其成分是半纤维素、纤维素、木质素和硅酸盐;再经过100 mL酸性洗涤剂洗涤后,剩余残渣烘干,其成分为为纤维素、木质素和硅酸盐,质量差为半纤维素质量;随后用72%硫酸洗涤,剩余物质烘干,其成分是木质素和硅酸盐,质量差为纤维素质量,再经马弗炉550 ℃烧制4 h,剩余的灰分即为硅酸盐质量,其质量差为木质素质量。
1.5.3 油茶籽粕中茶皂素的测定
采用香草醛-浓硫酸显色法测定茶皂素的含量变化[30],其原理为:茶皂素中含有皂苷, 皂苷是一类含有糖苷键的四环或五环三萜的化合物, 其C3、C12上的羟基可与香草醛上的醛基发生反应形成缩醛, 成为新的共轭体系而显色[32]。具体方法为:①茶皂素提取,称取105 ℃烘干的油茶粕5.00 g,与70%(v/v)乙醇按1∶7比例混合,于60 ℃水浴锅加热2 h,将滤液用70%(v/v)乙醇定容至50.00 mL;②茶皂素的测定,移取上述样液0.10 mL、蒸馏水0.40 mL及8%(v/m)香草醛无水乙醇溶液0.50 mL于10 mL试管中,将试管移至冰水浴中,加入77% (v/v)硫酸溶液4.00 mL,摇匀,再将试管放在恒温60 ℃的水浴锅中加热15 min,进行显色反应,取出试管放置到室温,用70%乙醇定容至10 mL,摇匀,以70%乙醇为空白,在波长为550 nm时测定,外标法定量[30]。
1.5.4 油茶籽粕的降解实验
将油茶籽粕原料置于烘箱中保持105 ℃条件烘干至恒重,粉碎后称取10.00 g放入锥形瓶中,加入10.00 mL的无菌蒸馏水,使其含水率为50%,并在121 ℃下灭菌20 min以去除样品中原有微生物的干扰。待冷却后,在无菌环境下加入30%筛选出来的菌种活化液,置于25 ℃、120 r/min条件下以半固态进行好氧发酵。发酵实验共持续28 d,在7、14、28 d取样,分别依据1.5.2节和1.5.3节中的步骤测定发酵后的油茶籽粕中纤维素、木质素及茶皂素含量,并计算3种物质的降解率。
1.5.5 生物炭的制备
将小麦秸秆用去离子水进行清洗,经烘干、粉碎后过0.150 mm筛,密封保存备用。将处理过的小麦秸秆置于马弗炉中700 ℃下缺氧炭化2 h,冷却后留备用[33]。
1.6 发酵产物指标测定
1.6.1 腐熟度测定
称取发酵28 d后的油茶籽粕试样(鲜样)10.00 g于250 mL锥形瓶中, 按照固液比1∶10加入相应质量的水,密封后垂直固定于摇床中,以25 ℃,120 r/min震荡浸提1 h,之后静置0.5 h,取上清液过滤摇匀后备用。
在培养皿中放置定性滤纸,之后均匀放入50粒大小一致、颗粒饱满的小白菜种子,加入上述浸提液10.00 mL,盖上保鲜膜并戳孔保持透气,在培养箱中25 ℃条件下避光培养48 h,统计发芽种子的粒数,并用游标卡尺逐一测量主根长,以水培发芽结果作为对照。通过公式(2)计算种子发芽指数(GI)[34]。
式中:A1为油茶籽粕有机肥料浸提液培养下的种子中发芽粒数百分比, %;A2为油茶籽粕机肥料浸提液培养的种子平均根长, mm;B1为水培种子中发芽粒数百分比, %;B2为水培种子的平均根长, mm。
取3次测定结果的算术平均值为最终测定结果,平行实验结果的绝对误差值保持在5%以内。其中,发芽指数大于50%视为基本腐熟,发芽指数大于85%视为完全腐熟。
1.6.2 产物pH及养分含量测定
称取5.00 g的油茶籽粕风干样(过0.250 mm筛)于烧杯中,加入50.00 mL不含CO2的水(经煮沸10 min驱除CO2),使用磁力搅拌器搅动3 min后静置30 min,用pH计测定产物的pH值。
将发酵28 d后的油茶籽粕样品烘干、粉碎并过0.150 mm筛,称取1.00 mg左右的样品用锡纸包覆后,使用EA3000元素分析仪测定其有机碳和养分氮含量[35]。
SOM=SOC×1.724 (3)
式中:SOM为有机质含量, g/kg;SOC为有机碳含量, g/kg;1.724为有机质和有机碳换算系数。
2 结果与分析
2.1 基本理化性质
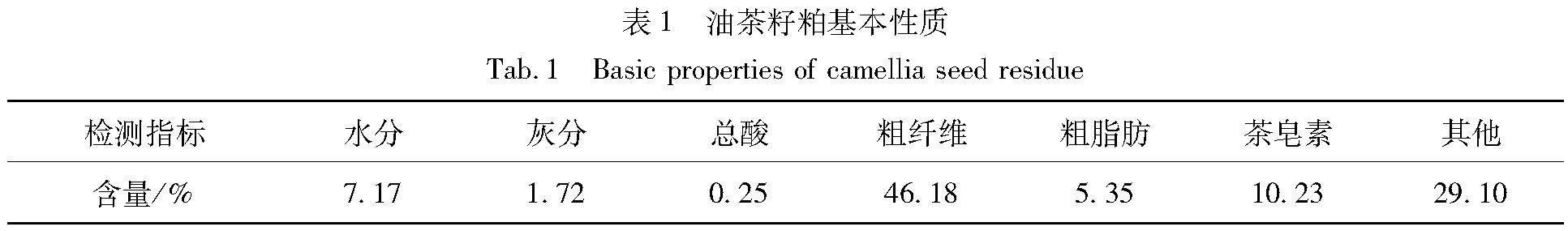
油茶籽粕的基本理化性质如表1所示。可以看出,粗纤维(主要成分是木质素和纤维素)和茶皂素含量较高,占总质量比例超过50%,如果不对其进行有效处理,难以得到效果较好的有機肥。
2.2 菌株筛选
微生物降解纤维素的效果可以根据其在富含纤维素的培养基上产生透明圈的大小来判断[36]。 实验结果表明, 菌株在2种培养基上呈现出明显的透明圈〔见图1(a)、(b)〕, 表明筛选的菌株具有较强的纤维素分解能力。 同一个平板经过多次分离、 纯化后, 得到1株纤维素降解菌, 编号为X1-1。
苯胺蓝固体培养基的褪色程度可以反应木质素降解菌的降解能力[37]。实验结果显示,部分菌株使苯胺蓝培养基完全褪色〔见图1(c)、(d)〕,说明该类菌株对木质素降解能力较强。同一个平板经过多次分离、纯化后,初步得到3株木质素降解菌,分别编号为M4-2、M4-3和M5-2。杜海萍等研究表明[38],单是苯胺蓝培养基的褪色情况仅能定性判断菌种是否产生过氧化氢酶,因此需要进一步分析菌株对于纯木质素的降解率。图2显示了菌株在7 d和14 d时对碱性纯木质素的降解率。可以看出,菌株对碱性木质素的降解率随发酵时间的增加而提高,且筛选的4种菌株中,M4-2和M5-2对碱性木质素的降解效果最好。
2.3 菌株降解性能
2.3.1 降解菌株的优选
通过筛选的4株降解菌株对油茶籽粕进行发酵,测定发酵产物中木质素、纤维素及茶皂素的降解率,以筛选出对油茶籽粕发酵效果最佳的菌株。4株降解菌株对油茶籽粕进行7 d发酵处理后,其对纤维素、木质素和茶皂素的降解率结果如图3所示。其中,菌株X1-1对纤维素和茶皂素的降解率最高,分别达到36.24%和14.28%;而菌株M5-2对木质素的降解率最高的,为20.55%;相较其他3株菌而言,菌株M4-2对3种物质的降解能力均不是最高,但综合效果较好。
因此,综合考虑4种菌株在纯木质素和油茶籽粕中的降解结果,确定选取菌株X1-1、M4-2和M5-2为目标进行进一步的菌种鉴定和后续发酵模拟试验。
2.3.2 菌株发酵对油茶籽粕中纤维素、木质素和茶皂素的降解效果
菌株X1-1、 M4-2和M5-2对油茶籽粕中纤维素、 木质素和茶皂素在不同发酵时间下的降解率如图4所示。 由图4(a)可知, 在菌株X1-1、 M4-2和M5-2的作用下, 木质素降解率与发酵时间呈正相关, 并且M5-2对木质素的降解率显著高于X1-1和M4-2, 在第28 d时最高降解率达到26.39%。 由图4(b)可知, 菌株M5-2对纤维素的降解率随着发酵时间的增加呈现升高趋势, 而菌株X1-1和M4-2对纤维素的降解率则呈现先降低后增加的趋势。 推测此现象的主要原因为范式粗纤维分析法所测纤维素初始含量并不包括被木质素包埋的形式所禁锢的部分[31], 导致纤维素降解率计算公式中分母取值偏小, 菌株在降解木质素过程中会释放部分额外的纤维素[39], 致使计算降解率数值降低。 伴随发酵过程的进行, 释放出的纤维素会进一步被菌株降解,使得计算降解率升高。 相较之下, 菌株M5-2对木质素降解能力较强, 在短期内即可释放被包埋的纤维素。 因此, 在整个发酵期内对纤维素的计算降解率较低, 但始终保持增长趋势。这与张仲卿等人研究发现混合真菌对玉米秸秆发酵产物中木质素和纤维素的降解率为36.80%和28.87%的结果相似[40]。 图4(c)显示 3种菌株对茶皂素也表现出了显著的降解性能, 而且降解率随着发酵时间延长而迅速增加, 在28 d后, X1-1、 M4-2和M5-2对油茶籽粕中的茶皂素降解率分别达到34.06%、 45.05%和46.15%。 此结果高于黄浦等人采用桔青霉(Penicillium citrinum)对茶皂素的降解率(36.71%)[41]。
综合木质素、纤维素和茶皂素的降解率随发酵时间的变化情况来看,第28 d的降解效果较好,说明适当延长发酵时间有助于3种物质特别是茶皂素的充分降解。
2.4 菌株鉴定
根据油茶籽粕发酵试验,综合其降解效果,确定菌株X1-1、M4-2和M5-2为最优目的菌株,对其进行形态观察以及DNA测序和同源性分析。如图5(a)和(d)、(b)和(e)、(c)和(f)分别为纯化后菌株X1-1、M4-2和M5-2的菌落形态及显微形态镜检图。菌株X1-1的菌落生长迅速,呈粉状至绒毛状或簇状,灰褐色,反面灰褐色至寡黑色,其为真菌,菌丝生长迅速,呈细长状,无隔中空且无分枝,孢子是由菌丝逐渐断裂形成的节孢子,孢子呈椭圆形。菌株M4-2菌落呈浅黄色不透明状,具有光滑的表面和规则的边缘,镜检图表明其呈短杆状,革兰氏染色为红色,革兰氏阴性菌。菌株M5-2形成菌落呈透明状圆形、表面光滑湿润、边缘整齐,镜检图表明其呈长杆状,革兰氏染色为红色,革兰氏阴性菌。
将X1-1的ITS和M4-2、M5-2的16S rDNA测序结果在NCBI中进行BLAST比对,构建系统发育树(见图6)。结果表明,X1-1、M4-2和M5-2分别与Pleurostomarichardsiae、Pseudomonas monteilii和Paenibacillussp位于同一个系统发育分支中,与模式菌株Pleurostomarichardsiae、Pseudomonas monteilii和Paenibacillussp菌株的序列相似性达99%,结合形态学特征、生理生化特性和系统发育的分析结果,将X1-1菌株鉴定为Pleurostomarichardsiae,M4-2菌株鉴定为蒙氏假单胞菌(Pseudomonas monteilii),M5-2菌株鉴定为类芽孢杆菌(Paenibacillus sp.)。
2.5 油茶籽粕发酵产物性质测定
图7为菌株处理的油茶籽粕发酵不同时间后发酵产物的种子发芽指数。按照农业行业标准《有机肥料》(NY/T 525—2021)要求,种子发芽指数(GI)需要达到70%以上。由图7可以看出,种子发芽指数随着发酵时间的延长而增加,只有在发酵时长达到28 d时油茶籽粕发酵产物的腐熟度才能达标。经X1-1、M4-2和M5-2发酵28 d后的浸提液培养的种子发芽指数分别为70.65%、72.18%和104.41%,全部符合有机肥的国家标准,且M5-2腐熟度最高,说明M5-2这株菌在发酵中更具有优势。
按照农业行业标准《有机肥料》(NY/T 525—2021)的要求,有机肥的pH值需要在5.5~8.5。测定发现,经过X1-1、M5-2和M4-2发酵后的油茶籽粕pH分别为5.52、5.0、4.74,发酵产物呈酸性的原因可能是微生物在发酵过程中产生了有机酸类物质,而真菌比细菌发酵产物pH偏高的原因可能是真菌在发酵的过程中会生成生物碱中和了一部分酸。生物炭具有微孔结构、较大的比表面积及富含碳等特点,可以有效改善土壤的酸碱度,提高土壤的保水性和通透性,给微生物制造一个良好的栖息空间和生存环境[42]。其pH为10.07,比表面积为71.209 m2/g,灰分含量为26.53%,产率为13.21%。因此将有机肥与生物炭按照1∶1的质量比向X1-1、M5-2和M4-2混合,得到混合有机肥的pH分别为7.08、6.34和6.16,全部符合有机肥的国家标准。由图8可以看出,由X1-1、M4-2和M5-2发酵28 d后的油茶籽粕制备的混合有机肥中有机质含量分别为89.15%、90.89%和82.51%,全部符合有机肥的国家标准(大于30%)。总养分(N+P+K)的含量分别为9.89%、8.73%、13.01%,同样均符合国家标准(大于4.0%)。
3 结论
本文筛选了能同时降解木质素、纤维素及茶皂素的功能菌株,并将其用于油茶籽粕发酵,成功制备了有机肥,主要结论如下。
1)从油茶籽粕中筛选纯化出3株木质素降解菌(M4-2、M4-3、M5-2)和1株纤维素降解菌(X1-1)。通过测定4种菌株对油茶籽粕中纤维素、木质素和茶皂素的降解率,综合分析得出,X1-1、M4-2和M5-2对油茶籽粕发酵28 d的效果较好。其中,X1-1对纤维素、木质素、茶皂素的降解率分别为29.74%、12.84%、34.06%; M4-2对纤维素、木质素、茶皂素的降解率分别为19.65%、17.85%、45.05%;M5-2对纤维素、木质素、茶皂素的降解率分别为13.19%、26.39%、46.15%。进而对X1-1、M4-2和M5-2进行菌种鉴定,分别是Pleurostomarichardsiae、蒙氏假单胞菌 (Pseudomonas monteilii)和类芽孢杆菌(Paenibacillus sp)。
2)油茶籽粕经过X1-1、M4-2和M5-2发酵28 d后,发酵产物提取液的种子发芽指数分别为70.65%、72.18%和104.41%,表明其腐熟度均达标,且M5-2的腐熟度最佳。经生物炭调节后的pH分别为7.08、6.34和6.16,有机质含量分别为89.15%、90.89%和82.51%,总养分含量为9.89%、8.73%、13.01%,均符合国家标准。
综上所述,本研究筛选出的3株菌株对木质素、纤维素及茶皂素均有较好的降解效果,添加功能菌株对油茶籽粕进行发酵制备有机肥,具有成本低、腐熟度高、养分充足、安全无害等优点,为油茶籽粕的大规模资源化利用提供了有效途径,具有良好的应用前景。
參考文献
[1] 钟旭美, 张百刚, 朱杰. 我国油茶籽的综合利用[J]. 粮油食品科技, 2007, 15(2): 34-36.
ZHONG X M, ZHANG B G, ZHU J, The comprehensive utilization of camellia oleifera seed[J]. Science and Technology of Cereals,Oils and Foods, 2007, 15(2): 34-36.
[2] 侯黔灵, 范丽美. 低共熔溶剂提取油茶籽壳中原花青素的工艺优化[J]. 中国油脂, 2023, 48(2): 129-132.
HOU Q L, FAN L M. Optimization of extraction of procyanidins from oil-tea camellia seed shell with deep eutectic solvents[J]. China Oils and Fats, 2023, 48(2): 129-132.
[3] 方飞, 王力生. 油茶粕资源综合开发利用研究进展[J]. 饲料博览, 2011(6): 30-32.
FANG F, WANG L S. Research progress of comprehensive development and utilization on the resources of oil-tea-cake[J]. Feed Review, 2011(6): 30-32.
[4] ZHAO Y, SU R Q, ZHANG W T, et al. Antibacterial activity of tea saponin from camellia oleifera shell by novel extraction method[J]. Industrial Crops and Products, 2020, 153: 112604.
[5] ZHANG J P, YING Y, LI X B, et al. Evaluation of three kinds of nutshell with respect to utilization as culture media[J]. BioResources, 2018, 13(4): 7508-7518.
[6] 赵光远, 刘倩茹, 王瑛瑶, 等. 油茶籽加工的研究进展[J]. 食品研究与开发, 2011, 32(10): 169-172.
ZHAO G Y, LIU Q R, WANG Y Y, et al. Research progress on the processing of camellia oleifera seeds[J]. Food Research and Development, 2011, 32(10): 169-172.
[7] WANG R, ZHANG J Y, SUI Q W, et al. Effect of red mud addition on tetracycline and copper resistance genes and microbial community during the full scale swine manure composting[J]. Bioresource Technology, 2016, 216: 1049-1057.
[8] TAHERZADEH M J, KARIMI K. Pretreatment of lignocellulosic wastes to improve ethanol and biogas production: A review[J]. International Journal of Molecular Sciences, 2008, 9(9): 1621-1651.
[9] 张晓燕. 不同预处理方法对玉米秸秆和沙柳纤维素降解率和乙醇产量的影响[D]. 呼和浩特: 内蒙古工业大学, 2017.
[10]BHALLA A, BANSAL N, KUMAR S, et al. Improved lignocellulose conversion to biofuels with thermophilic bacteria and thermostable enzymes[J]. Bioresource Technology, 2013, 128: 751-759.
[11]ANDLAR M, REZIC T, MARDETKO N, et al. Lignocellulose degradation: An overview of fungi and fungal enzymes involved in lignocellulose degradation[J]. Engineering in Life Sciences, 2018, 18(11): 768-778.
[12]賈辉, 陈秀蓉, 芦光新, 等. 纤维素降解细菌的筛选、生物学特性及降解效果[J]. 草业学报, 2016, 25(3): 60-66.
JIA H, CHEN X R, LU G X, et al, lsolation of cellulose-degrading bacteria and determination of their degradation activity[J]. Acta Prataculturae Sinica, 2016, 25(3): 60-66.
[13]王洪媛, 范丙全. 三株高效秸秆纤维素降解真菌的筛选及其降解效果[J]. 微生物学报, 2010, 50(7): 870-875.
WAGN H Y, FAN B Q. Screening of three straw-cellulose degrading microorganism[J]. Acta Microbiologica Sinica, 2010, 50(7): 870-875.
[14]张晨敏. 低温纤维素降解菌的筛选及复合菌剂在秸秆还田中的应用[D]. 南京: 南京农业大学, 2014.
[15]王郝为. 油茶粕发酵处理及饲用价值评定[D]. 长沙: 中南林业科技大学, 2016.
[16]邓桂兰. 混合菌发酵油茶粕生产菌体蛋白饲料的研究[J]. 粮食与饲料工业, 2008(6): 30-32.
DENG G L. Research on protein feed produced from oil-tea meal fermented by mixed germs[J]. Cereal & Feed Industry, 2008(6): 30-32.
[17]黄丹莲. 堆肥微生物群落演替及木质素降解功能微生物强化堆肥机理研究[D]. 长沙: 湖南大学, 2011.
[18]黄卫文, 敖常伟, 钟海雁. 油茶皂素抑菌效果研究[J]. 经济林研究, 2002, 20(1): 17-19.
HUANG W W, AO C W, ZHONG H Y. The Antibacterial effect of oiltea saponin[J].Non-wood Forest Research, 2002, 20(1): 17-19.
[19]文莉, 芦苇, 蒋倩, 等. 茶皂素毒性刺激性试验及抑菌作用研究[J]. 中国油脂, 2011, 36(6): 58-60.
WEN L, LU W, JIANG Q, et al. Toxicity,irritation and bacteriostasis of tea saponin[J]. China Oils and Fats, 2011, 36(6): 58-60.
[20]叶雪良. 茶皂素的开发与利用[J]. 化工生产与技术, 2002, 9(2): 6-8.
YE X L. Development and utilization of tea saponin[J]. Chemical Production and Technology, 2002, 9(2): 6-8.
[21]侯俊杰, 李宁宁, 吕辉雄, 等. 茶皂素对种子发芽、根长及土壤酶活性的影响[J]. 农业环境科学学报, 2015, 34(4): 660-665.
HOU J J, LI N N, LYU H X, et al. Effects of tea saponin on seed germination, root lengths and soil enzyme activities[J]. Journal of Agro-Environment Science, 2015, 34(4): 660-665.
[22]李灵灵, 王敬红, 赵铎, 等. 木质素降解菌BYL-7的筛选及降解条件优化[J]. 微生物学通报, 2020, 47(12): 4059-4071.
LI L L, WANG J H, ZHAO D, et al. Screening of lignin degrading strain BYL-7 and optimization of degradation conditions[J]. Microbiology China, 2020, 47(12): 4059-4071.
[23]孟建宇, 陳勿力吉玛, 郭慧琴, 等. 常温和低温纤维素降解菌的分离及其降解特性[J]. 农业生物技术学报, 2021, 29(1): 73-84.
MENG J Y, CHEN W L J M, GUO H Q, et al. Isolation and degradation characteristics of cellulose-degradation bacteria at room and low temperature[J]. Journal of Agricultural Biotechnology, 2021, 29(1): 73-84.
[24]杨勇, 聂旭, 王健, 等. 油茶饼粕纤维素降解菌的筛选与鉴定[J]. 江苏农业科学, 2014, 42(5): 299-301.
YANG Y, NIE X, WAGN J, et al. Screening and characterization of cellulose degrading bacteria in oil tea cake meal[J]. Jiangsu Agricultural Sciences, 2014, 42(5): 299-301.
[25]国家卫生和计划生育委员会. 食品安全国家标准食品中水分的测定: GB 5009.3—2016[S].北京: 中国标准出版社, 2017.
[26]国家卫生和计划生育委员会. 食品安全国家标准食品中灰分的测定: GB 5009.4—2016[S].北京: 中国标准出版社, 2017.
[27]国家质量监督检验检疫总局. 食品中总酸的测定: GB/T 12456—2008[S]. 北京: 中国标准出版社, 2009.
[28]国家卫生健康委员会. 植物类食品中粗纤维的测定: GB/T 5009.10—2003[S]. 北京: 中国标准出版社, 2004.
[29]国家卫生和计划生育委员会,食品安全国家标准食品中脂肪的测定: GB 5009.6—2016[S].北京: 中国标准出版社, 2017.
[30]LI M, WANG C. Study on extraction of tea saponin from camellia cake [J]. Journal of the Chinese Cereals and Oils Association, 2011, 26(5): 38-41.
[31]SOEST P J V.Use of detergents in the analysis of fibrous feeds. 2. A rapid method for the determination of fiber and lignin[J].Journal of AOAC International, 1963, 49(4):546-551.
[32]陈莹, 刘松柏, 何良兴, 等. 油茶籽粕和茶皂素中皂苷的定量检测方法研究[J]. 中国粮油学报, 2012, 27(2): 105-111.
CHEN Y, LIU S B, HE L X, et al. Quantitative analysis of saponins in camellia seed cake and tea saponins[J]. Journal of the Chinese Cereals and Oils Association, 2012, 27(2): 105-111.
[33]张杰. 生物炭固定化高效解磷菌对铅污染土壤的修复研究[D]. 西安: 西北大学, 2019.
[34]中华人民共和国农业部. 有机肥料: NY 525—2012[S].北京: 中国标准出版社, 2012.
[35]WANG J, FU X, ZHAO F Z, et al. Response of soil carbon fractions and dryland maize yield to mulching[J]. Soil Science Society of America Journal, 2018, 82(2): 371-381.
[36]骆婷, 夏虹, 冯定胜, 等. 森林土壤中多功能降解菌的分离筛选及鉴定[J]. 西南农业学报, 2018, 31(5): 1032-1040.
LUO T, XIA H, FENG D S, et al. Isolation and identification of multi-function deqradation bacteria from forest soi[J]. Southwest China Journal of Agricultural Sciences, 2018, 31(5): 1032-1040.
[37]张东升, 徐一馨, 宋双民, 等. 细菌产纤维素酶和脂肪酶固体培养基及接种和培养时间的研究[J]. 微生物学杂志, 2022, 42(3): 44-53.
ZHAGN D S, XU Y X, SONG S M, et al. Inoculation and culture time of bacterial cellulase and lipase production with solid culture medium[J]. Journal of Microbiology, 2022, 42(3): 44-53.
[38]杜海萍, 宋瑞清, 王钰祺. 几种真菌产木质素降解酶的比较研究[J]. 林业科技, 2006, 31(4): 20-24.
DU H P, SONG R Q, WANG Y Q. Comparative study on the lignocellulolytic enzymes produced by fungi[J]. Forestry Science & Technology, 2006, 31(4): 20-24.
[39]李希越, 王洪波, 赵玉晓, 等. 好氧生物预处理时间对玉米秸秆水解酸化的影响[J]. 生物质化学工程, 2022, 56(1): 13-22.
LI X Y, WAGN H B, ZHAO Y X, et al. Effect of aerobic biological pretreatment time on hydrolytic acidification of corn straw[J]. Biomass Chemical Engineering, 2022, 56(1): 13-22.
[40]张仲卿, 张爱忠, 姜宁. 混合真菌发酵对玉米秸秆纤维素与木质素降解率的影响[J]. 动物营养学报, 2019, 31(3): 1385-1395.
ZHANG Z Q, ZHAGN A Z, JIANG N. Effects of mixed fungal fermentation on degradation rate of cellulose and lignin of corn straw[J]. Chinese Journal of Animal Nutrition, 2019, 31(3): 1385-1395.
[41]黃浦, 肖瑜, 刘以清, 等. 桔青霉降解茶皂素的实验研究[J]. 桂林理工大学学报, 2016, 36(3): 592-596.
HUANG P, XIAO Y, LIU Y Q, et al. Degradation of tea saponin by penicillium citrinum[J]. Journal of Guilin University of Technology, 2016, 36(3): 592-596.
[42]HUSSAIN R, KUMAR GHOSH K, RAVI K. Impact of biochar produced from hardwood of mesquite on the hydraulic and physical properties of compacted soils for potential application in engineered structures[J]. Geoderma, 2021, 385: 114836.
猜你喜欢
纺织科技进展(2021年3期)2021-06-09
陶瓷学报(2021年1期)2021-04-13
今日农业(2020年19期)2020-12-14
中国化肥信息(2020年9期)2020-03-29
上海包装(2019年8期)2019-11-11
中国化肥信息(2017年3期)2017-12-23
中国化肥信息(2017年7期)2017-12-13
天津造纸(2016年1期)2017-01-15
中国造纸学报(2015年1期)2015-12-16
中国塑料(2014年4期)2014-10-17
