
番茄SlMYB48基因生物信息学及表达分析
2024-04-30 06:17刘佳凤郭晓青王桂强王虹云朱桐曹守军姚建刚张丽莉张瑞清赵婧李涛
中国瓜菜 2024年4期
刘佳凤 郭晓青 王桂强 王虹云 朱桐 曹守军 姚建刚 张丽莉 张瑞清 赵婧 李涛

收稿日期:2023-10-10;修回日期:2023-12-20
基金项目:烟台市科技计划项目(2022XCZX091);国家现代农业产业技术体系专项(CARS-23-G11);重庆市巫山县科技项目(wskjdxbxm2023004);国家自然科学基金面上项目(32372737);西南作物基因资源发掘与利用国家重点实验室开放课题(SKL-KF202224)
作者简介:刘佳凤,女,在读硕士研究生,研究方向为番茄抗逆基因的验证。E-mail:19558916823@163.com
通信作者:李 涛,男,正高级农艺师,研究方向为蔬菜育种及分子生物学。E-mail:ytnkyscs@163.com
DOI:10.16861/j.cnki.zggc.202423.0652
摘 要:MYB转录因子是植物转录因子家族中数量最多、用途最广的成员之一,为挖掘更多番茄(Solanum lycopersicum)MYB转录因子家族成員信息,初步探究其表达模式及功能,以番茄Ailsa Craig为试材,采用RT-PCR的方法克隆SlMYB48基因,并对其进行生物信息学及表达、定位分析。结果表明,番茄SlMYB48基因的开放阅读框(ORF)长度为708 bp,编码235个氨基酸,在番茄的根中表达量最高,叶中次之。SlMYB48蛋白含有保守的MYB结构域,定位于细胞核中,属于不稳定、亲水性蛋白。对SlMYB48启动子分析,发现其含有大量的逆境响应元件,qRT-PCR及RNA-seq 数据库分析结果表明,高盐、生物逆境胁迫条件下,SlMYB48基因表达量均随处理时间延长而升高,干旱胁迫条件下表达量下降,推测其可能参与番茄生物及非生物逆境胁迫反应。研究结果为进一步深入探究番茄SlMYB48基因的功能及作用机制提供了参考。
关键词:番茄;SlMYB48;表达分析;定位
中图分类号:S641.2 文献标志码:A 文章编号:1673-2871(2024)04-027-09
Bioinformatics and expression analysis of tomato SlMYB48 gene
LIU Jiafeng1, GUO Xiaoqing2, WANG Guiqiang3, WANG Hongyun4, ZHU Tong1, CAO Shoujun4, YAO Jiangang4, ZHANG Lili4, ZHANG Ruiqing4, ZHAO Jing1, LI Tao1, 4
(1. College of Life and Science, Yantai University, Yantai 264000, Shandong, China; 2. Yantai Agricultural Technology Extension Center, Yantai 265499, Shandong, China; 3. Agricultural Comprehensive Service Center, Zhangxing Town, Zhaoyuan 265403, Shandong, China; 4. Yantai Academy of Agricultural Sciences, Yantai 264500, Shandong, China)
Abstract: MYB transcription factor is one of the most abundant and widely used members of the plant transcription factor family. In order to mine more information about MYB transcription factor family members of tomato (Solanum lycopersicum) and preliminarily explore its expression pattern and function, this study took tomato Ailsa Craig as the test material. SlMYB48 gene was cloned by RT-PCR, and its bioinformatics, expression and localization were analyzed. The results showed that the open reading frame (ORF) length of tomato SlMYB48 gene was 708 bp, encoding 235 amino acids, and the expression level was the highest in tomato roots, followed by in leaves. The SlMYB48 protein contains a conserved MYB domain, which is an unstable, hydrophilic protein located in the nucleus . SlMYB48 promoter analysis found that it contained a large number of stress response elements. The results of qRT-PCR and RNA-seq database analysis showed that the expression of SlMYB48 gene increased with the extension of treatment time under high salt and biological stress, and decreased under drought stress, suggesting that SlMYB48 gene might be involved in biological and abiotic stress response of tomato. The results of this study provide a reference for further investigation of the function and mechanism of tomato SlMYB48 gene.
Key words: Solanum lycopersicum; SlMYB48; Expression analysis; Location
番茄(Solanum lycopersicum)具有良好的口感、丰富的营养、繁多的品种以及较高的研究价值和经济价值,成为我国乃至世界上栽培最为广泛的蔬菜作物之一。在生物学水平上,番茄也是应用于多种学科领域、进行遗传转化研究的非常重要的模式植物。自然环境中的高盐、干旱、低温等逆境因素严重影响番茄的生长发育和产量,因此了解番茄在抗逆反应过程中的作用机制,进而探寻抗逆基因资源、选育番茄抗逆品种在理论和生产上具有重大意义。
MYB转录因子是植物转录因子家族中数量最多、用途最广的成员之一[1]。MYB转录因子是基于编码蛋白含有一个或多个高度保守的DNA结构域-MYB结构域,含有大约50个氨基酸残基组成的不完全重复的R结构(R1,R2和R3)[2]。根据每个基因中R结构的数量,可分为4个亚类[3]。其中,R1/R2-MYB亚类只包含1个MYB结构域,R2R3-MYB亚类包含2个MYB结构域,R1R2R3-MYB亚类包含3个连续结构域,4R-MYB亚类包含4个MYB结构域[4-6]。
在植物中第一个被发现的MYB类基因是从单子叶植物玉米中克隆出来的ZmMYBC1基因,该基因为色素合成有关基因[7]。随着基因组学和分子生物学的发展,越来越多的MYB家族成员被鉴定。MYB转录因子不仅能对激素以及干旱、高温、高盐等逆境胁迫作出应答[8],而且广泛参与植物初生和次生代谢反应[9],并对植物的生长发育和形态建成[10],具有重要的调控作用。MYB转录因子参与植物的生长发育调控,AtMYB23基因参与拟南芥根表皮发育过程中的正调控途径[11]。植物MYB类转录因子在植物的代谢生理活动中也发挥作用,玉米ZmMYB028和ZmMYB073成员与苯丙烷生物合成负调控相关[12]。已有研究表明,番茄MYB类转录因子在响应干旱胁迫方面发挥着重要作用。在干旱胁迫处理下,SlMYB10和SlMYB14两个基因相对表达量呈升高趋势,将SlMYB14基因沉默后,通过对生理生化指标以及活性氧的测定,表明SlMYB14基因对番茄耐盐性和抗旱性有重要作用[13]。沉默SlMYB71基因后,促进了番茄植株在干旱胁迫下的生长,降低了活性氧含量、相对电导率和丙二醛含量,影响叶绿素积累和胁迫基因的表达,表明SlMYB71基因在非生物胁迫方面起着负调控作用[14]。
笔者前期通过海水栽培番茄的转录组分析,发现随海水浓度增加,SlMYB48表达量升高[15]。为了进一步研究SlMYB48转录因子在番茄生长发育以及抗逆过程中的功能和机制,利用RT-PCR方法克隆了SlMYB48基因,并对其进行生物信息学、启动子响应元件、表达、定位等分析,以期为后续研究番茄SlMYB48基因的功能及作用机制奠定基础。
1 材料与方法
1.1 材料
试验用番茄品种为Ailsa Craig,种子由山东省烟台市农业科学研究院蔬菜花卉研究所保存。试验于2022年10月至2023年7月在山东省烟台市农业科学研究院蔬菜花卉研究所实验室进行。取新鲜的番茄叶片,锡箔纸包裹置于液氮中冷冻,并在-80 ℃保存,用于RNA提取。用于组织表达分析取样的部位及时期:叶为完全展开的功能叶;茎为生长点下端第2节位茎段;根为清洗后的白色根段;花朵为开放当天的花;成熟果为红色的正常健康果实。将番茄种子播种于20 cm×20 cm塑料盆中,栽培基质按草炭、蛭石、珍珠岩体积比2∶1∶1进行配置,在植株5叶1心期时,进行干旱胁迫和高盐胁迫处理,将200 mL10%的聚乙二醇6000 (PEG-6000)溶液、200 mL的NaCl溶液(200 mmol·L-1)分别灌溉番茄植株,在1、6、12、24、48、72 h分别取番茄叶片,液氮冷冻,-80 ℃保存,用于逆境胁迫表达分析。亚细胞定位所用植物材料为本氏烟草(Nicotiana benthamiana L.),将烟草种子播种于7 cm×7 cm穴盘中,光周期设置为光照12 h/黑暗12 h,在温度为(25±2) ℃光照培养箱中培养1个月,用于烟草瞬时转化。
RNA提取试剂盒购于北京华越洋生物科技有限公司;反转录试剂盒购于天根生化科技有限公司;高保真DNA聚合酶购于北京全式金生物技术有限公司;质粒提取试剂盒和DNA回收试剂盒购于艾科瑞生物科技有限公司。其他化学药品购于北京聚合美生物科技有限公司。引物由上海生工生物技术有限公司合成。
1.2 方法
1.2.1 总RNA提取及cDNA第一链的合成 将番茄嫩叶用液氮研磨,按照华越洋RNA提取试剂盒操作说明提取植物组织总RNA,利用1%琼脂糖凝胶电泳检测其完整性,使用天根cDNA第一链合成试剂盒将其反转录获得cDNA,然后置于-20 ℃保存,用于后续试验。
1.2.2 番茄SlMYB48基因的克隆 从NCBI中获得番茄MYB家族基因成员SlMYB48的序列,利用Primer 5工具设计全长引物,引物序列为F:ATGGCACAAGAAGAAATGAGAAGAG;R:TTAGCCAGCAAAGAATGTGTTATCCT,以cDNA为模板,扩增SlMB48全长序列。PCR扩增程序为:95 ℃预变性3 min;95 ℃变性20 s,51 ℃退火20 s,72 ℃延伸22 s,共40个循环,72 ℃延伸5 min。用2%琼脂糖凝胶电泳分离PCR产物,切胶后使用华越洋DNA凝胶回收试剂盒回收目的片段,将扩增产物连接至pMD18-T克隆载体,连接产物转化至大肠杆菌感受态细胞DH5α,挑取经PCR鉴定的阳性克隆,送样测序。
1.2.3 SlMYB48基因生物信息学分析 在NCBI中利用Conservation Domain程序(https://www.ncbi.nlm.nih.gov/ cdd/?term=)进行基因保守结构域分析。在ProtParam网站(https://web.expasy.org/protparam/)計算蛋白的等电点、分子质量和分子式。运用TMHMM Server v. 2.0 (http://www.cbs.dtu.dk/services/TMHMM/)进行蛋白的跨膜域预测;通过SignalP-4.1(https://services.healthtech.dtu.dk/service.php?SignalP-4.1)预测信号肽。利用ExPasy-ProtScale(https://web.expasy.org/protscale/)预测蛋白质亲水疏水性。利用SOPMA网站(https://npsa-prabi.ibcp.fr/cgi-bin/npsa _automat.pl?page=npsa%20_sopma.html)预测蛋白质二级结构。利用Swiss Model(https://swissmodel.expasy.org/)网站进行同源建模预测三级结构。在NCBI网站中利用blastn(https://blast.ncbi.nlm.nih.gov/Blast.cgi? PROGRAM=blastn& PAGE_TYPE=BlastSearch&LINK_LOC=blasthome)将SlMYB48基因与蛋白nr数据库比对。利用DNAMAN对各物种中的MYB的氨基酸序列进行比对。使用MEGA7软件的邻接法构建系统进化树。
1.2.4 SlMYB48表达模式分析 采用CTAB方法提取番茄不同组织部位RNA,并反转录为cDNA。通过Primer Premier 5软件设计荧光定量PCR引物(F:ACGATACGGGTGGACTTGAC;R:ATCCTGGTTGTCCAAGCAAT,扩增片段为227 bp),选取actin为内参基因,引物序列为F:GGAATGGGACAGAAGGAT;R:CAGTCAGGAGAACAGGGT,扩增产物为183 bp,以获得的cDNA为模板,参照iScript? gDNA Clear cDNA Synthesis Kit试剂盒(伯乐生命医学产品上海有限公司)说明书,进行实时荧光定量PCR扩增。采用7500型实时荧光定量PCR仪(ABI,USA)进行分析,PCR反应体系为:2×qPCR Mix 10 μL,正反向引物(10 μmol·L-1)各0.5 μL,cDNA单链模板1 μL,ddH2O 8 μL,共20 μL。PCR反应条件为:95 °C预变性5 min,95 °C变性10 s,60 ℃退火30 s,40个循环,60 °C 延伸 1 min。设置3次生物学重复,使用2-△△Ct法计算基因的相对表达量。数据通过Excel计算并绘制柱状图。
利用番茄基因组数据库(https://solgenomics.net)查询番茄SlMYB48基因相关表达量。在番茄基因组数据库工具栏内进入番茄表达数据库(TED),在表达量一栏下的RNA-seq数据库中输入基因ID:Solyc06g005310进行表达量查询。选取试验编号为D007,该基因在生物胁迫下,受细菌菌株(荧光假单胞菌、恶臭单胞菌和农杆菌)处理的多个表达量作为参考,分析SlMYB48基因在生物胁迫下的表达模式,并将SlMYB48基因数据通过Excel整理、绘制柱状图。
1.2.5 亚细胞定位 使用 Primer Premier 5 软件设计引物,并且在特异性引物上添加BsaI和Eco31I酶切位点及保护碱基(F:cagtCGTCTCacaacatggcacaagaagaaatgag;R:cagtCGTCTCatacagccagcaaagaatgtgttat),按照1.2.2 PCR扩增体系对番茄叶片cDNA进行扩增,反应程序为:95 ℃ 3 min;95 ℃ 20 s,51 ℃ 20 s,72 ℃ 22 s,共40个循环,72 ℃延伸5 min,采用1%琼脂糖凝胶电泳分离PCR产物,切胶回收目的片段。使用BsaI和Eco31I限制性内切酶,对载体和胶回收片段进行双酶切,载体双酶切体系如下:Nuclease-free Water 13 ?L,10×Buffer 2 ?L,BsaI/Eco31I 1 ?L,pBWA(V)HS-ccdb-GLosgfp 4 ?L,共20 ?L,37 ℃反应 1 h。目的基因胶回收片段酶切体系同上。按照PCR反应液纯化试剂盒说明书纯化酶切产物。得到纯化产物后进行连接。连接体系为:Nuclease-free Water 5.5 ?L,10×Buffer 1 ?L,T4DNAligase 1 ?L,纯化产物2.5 ?L,共10 ?L。将重组产物转化到大肠杆菌DH5α细胞,菌液PCR筛选阳性克隆子进行测序,将测序正确的阳性菌落摇菌培养提取质粒。将表达载体转入农杆菌GV3101后,将菌液涂布于卡那霉素抗性YEB 固体培养基上,挑取单克隆放入YEB液体培养基中,在28 ℃下培养2 d,离心去上清液后,用10 mmol·L-1 MgCl2(含120 μmmol·L-1 AS)重悬菌体,调节OD600至0.6左右。从烟草叶片下表皮(背面)压迫注射菌液,弱光培养2 d。制作玻片,在激光共聚焦显微镜下观察并拍照,同时以空载体转化的农杆菌作为对照,重复上述操作。
2 结果与分析
2.1 SlMYB48基因的克隆
以番茄嫩叶cDNA为模板,以全长引物进行PCR扩增,获得基因全长708 bp(图1),编码235个氨基酸。
2.2 生物信息学分析
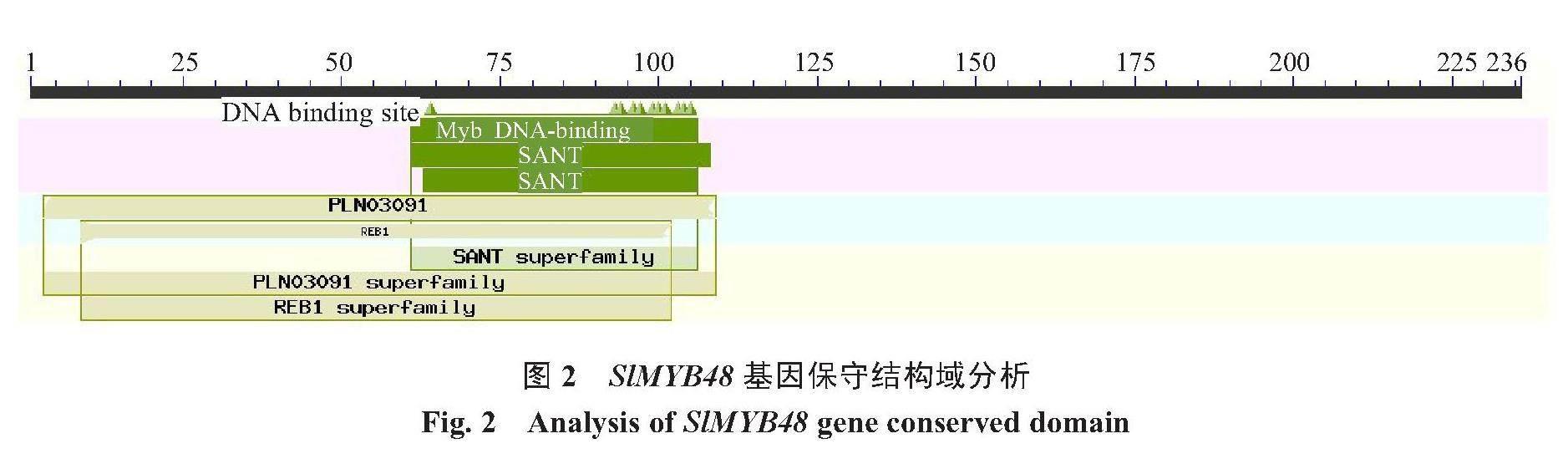
2.2.1 SlMYB48转录因子的保守结构域分析 为了分析SlMYB48蛋白保守结构域,将得到的核苷酸序列翻译成蛋白序列后,在NCBI的功能结构域选项中进行预测。结果表明,SlMYB48基因含有典型的MYB转录因子家族的DNA结构域,还含有PLN03091、REB1和SANT结构域(图2)。
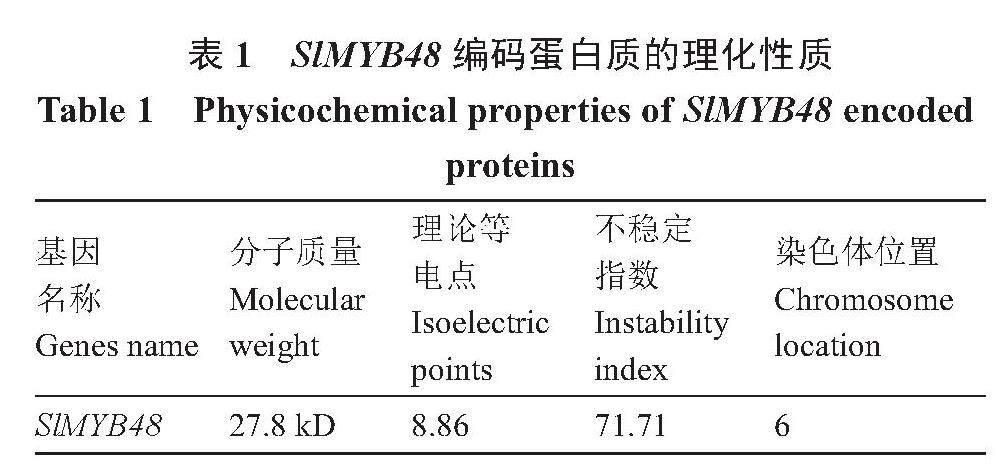
2.2.2 SlMYB48编码蛋白质的理化性质和信号肽以及编码蛋白质产物的亲水性疏水性分析 利用在线软件Protparam分析SlMYB48蛋白质的理化性质,其分子式为C1226H1874N348O365S15,分子质量为27.8 kD,理论等电点pI为8.86。其编码蛋白质的氨基酸组成中丝氨酸(Ser)占比最高,为9.4%,其次是赖氨酸(Lys),为8.1%,天冬酰胺(Asn)为8.1%。蛋白不稳定性指数为71.71,属于不稳定蛋白(表1)。TMHMM在线预测该蛋白的跨膜结构,SlMYB48转录因子所编码的235个氨基酸均不在跨膜区,说明SlMYB48蛋白不含跨膜结构,不属于跨膜蛋白(图3-A)。通过Signal IP4.0软件分析该蛋白信号肽,结果表明SlMYB48蛋白序列无信号肽(图3-B)。
利用ExPasy-ProtScale对SlMYB48蛋白进行亲水性疏水性分析,结果如图4所示,-3.233为最小的负值,1.556为最大的正值,负值的绝对值大于正值的绝对值,表明该蛋白为亲水性蛋白。
2.2.3 SlMYB48蛋白结构分析 运用在线软件SOPMA预测SlMYB48的二级结构,结果显示SlMYB48蛋白中不规则卷曲含量高达49.79%,其次为α-螺旋,占36.17%,延伸链占7.23%,最少的是β-折叠,仅占6.81%(图5-A)。
为了验证笔者对SlMYB48蛋白二级结构预测的结果,利用SWISS-MODEL网站對SlMYB48的蛋白三级结构进行建模(图5-B),从图中可以看出,SlMYB48蛋白结构主要由无规则卷曲和螺旋结构构成,与上述的蛋白二级结构预测结果相符。
2.2.4 蛋白的同源比较及进化分析 通过NCBI中Blast检索,将SlMYB48与蛋白nr数据库比对,分析得知SlMYB48与其他植物的 MYB蛋白有高度的同源性(图6),说明其蛋白的结构和功能都高度保守。结果表明,同源性最高的是潘那利番茄MYB48(XM_015222064.2),同源性达到98.07%;其次是马铃薯MYB48(XM_006366855.2),同源性为96.92%;野生马铃薯MYB48(XM_049519716.1)、窄刀薯MYB48(XM_049556356.1)、辣椒MYB48(XM_016721740.2)这3个MYB基因编码的氨基酸序列同源性也具有较高的同源性,分别为96.72%、96.34、89.39%。
为了进一步分析SlMYB48与其他植物MYB之间的进化层面关系,在上述氨基酸多重比对的基础上,利用MEGA 7软件对栽培番茄(Solanum lycopersicum)、潘那利番茄(Solanum pennellii)、马铃薯(Solanum tuberosum)、野生马铃薯(Solanum verrucosum)、窄刀薯(Solanum stenotomum)、辣椒(Capsicum annuum)、绒毛状烟草(Nicotiana tomentosiformis)、渐狭叶烟草(Nicotiana attenuata)和林烟草(Nicotiana sylvestris)这9种植物的MYB氨基酸序列进行系统进化树分析。通过进化树分析(图7)可以看出,与番茄SlMYB48蛋白亲缘关系最近的是潘那利番茄,被聚为一组,其次是马铃薯,这与氨基酸多重比较的结果一致,不同植物中的MYB蛋白进化关系分析结果基本符合植物分类学地位,其种属特性比较明显,即种属关系越近的亲缘关系越近。
2.2.5 SlMYB48启动子顺式作用元件分析 利用软件Plant CARE对SlMYB48基因启动子序列上的顺式作用元件进行预测分析,如表2所示,SlMYB48启动子中含有大量与逆境胁迫、激素响应和光响应相关的顺式作用元件,包括光响应元件GA-motif、GATA-motif、TCCC-motif、G-Box等,逆境响应元件ABRE、ARE、TCA-element等,表明SlMYB48可能参与相应生长发育与逆境胁迫响应过程。
2.3 SlMYB48基因的表达分析
2.3.1 SlMYB48基因在番茄不同组织中的表达 以番茄肌动蛋白基因 actin为内参,qRT-PCR分析结果表明,SlMYB48 基因在番茄苗的根、茎、叶和花中均有不同程度的表达,其中在根和叶中的表达量较高,在花和茎中表达量极低,在果实中几乎不表达(图8)。
2.3.2 非生物胁迫对SlMYB48基因表达量的影响 对盐胁迫和干旱胁迫下SlMYB48的表达量进行分析,如图9所示。与对照相比,高盐胁迫处理下,6 h后SlMYB48在番茄叶片中表达量迅速上升,并在48 h达到最大值,之后略有下降,但表达量均高于对照。模拟干旱条件下,随处理时间延长,SlMYB48表达量呈先降低后升高的变化趋势,处理1 h,表达量约为对照的一半,12 h后明显低于对照植株。以上结果表明SlMYB48对高盐及干旱均产生胁迫响应。
2.3.3 生物胁迫对番茄SlMYB48基因表达量的影响 番茄叶片在经过细菌菌株(荧光假单胞菌、恶臭假单胞菌和根癌农杆菌)处理后,SlMYB48基因表达量均随处理时间延长上调,其中根癌农杆菌受诱导更明显,SlMYB48表达量在6 h后达到17.23。受荧光假单胞菌诱导较弱,说明SlMYB48可能参与番茄生物胁迫(图10)。
2.4 SlMYB48基因的亚细胞定位
为了明确SlMYB48蛋白在细胞中的作用部位,利用农杆菌介导的瞬时转化法,将培养好的带有SlMYBS48与GFP融合蛋白载体的农杆菌注射到烟草叶片,培养2 d后在显微镜下观察融合蛋白在烟草叶片细胞中的分布情况。以空载体注射烟草的结果作为对照。结果表明,SlMYB48蛋白定位在细胞核中,说明该蛋白主要在细胞核中表达(图11)。
3 讨论与结论
R2R3-MYB蛋白是植物特有的丰富蛋白,在植物的基因组中大约有100多个R2R3-MYB成员,参与植物生长发育、次生代谢调控、生物和非生物胁迫应答[16]。笔者通过RT-PCR技术克隆得到了全长为708 bp的番茄SlMYB48基因的目的片段,位于番茄6号染色体上。通过生物信息学分析得知,SlMYB48是一种不稳定的亲水性蛋白,不含跨膜结构域和信号肽。SlMYB48蛋白编码了235个氨基酸,对其保守结构域分析表明,SlMYB48蛋白含有2个植物MYB转录因子家族的特异性结构域-SANT-myb DNA结合结构域,表明SlMYB48为R2R3型MYB转录因子。
通过与其他植物MYB氨基酸比对分析,发现番茄SlMYB48与其他植物的MYB48氨基酸表现出了较高的同源性,其中与马铃薯和潘那利番茄的同源性较高,表现出较近的进化关系。已有研究表明,StMYB48-like主要参与调控花青素的合成,在植物根的生长中起重要调节作用,同时参与干旱和高盐等非生物胁迫耐受性[17]。有研究表明,在水稻中过表达OsMYB48-1能显著提高水稻的耐旱性和耐盐性[18]。在拟南芥中过表达水稻ZmMYB48,发现ZmMYB48可能是通过ABA信号参与干旱胁迫响应的积极调控因子,表明不同物种的同种基因可能有相似的功能[19],据此推测番茄SlMYB48基因可能与调控番茄植株抗逆特性有关。
笔者对番茄SlMYB48基因启动子序列上的顺式作用元件进行预测分析发现,番茄SlMYB48启动子中含有大量与逆境胁迫、激素响应和光响应相关的顺式作用元件。AtMYB48在拟南芥根中表达量最高,且与野生型相比,在H2O2介导的氧化作用下,AtMYB48敲除后植株的根长显著减少,表明AtMYB48在根发育中具有正调控作用[20]。在笔者的研究中,不同部位表达模式分析表明,番茄SlMYB48基因在番茄根中表達量最高,这与在拟南芥中的研究结果一致,推测SlMYB48基因在番茄中也存在相似的功能,表明SlMYB48基因可能在抵抗逆境胁迫和根的生长发育中发挥重要作用。
猜你喜欢
小猕猴学习画刊(2022年12期)2022-02-06
今日农业(2021年21期)2022-01-12
今日农业(2020年23期)2020-12-15
导航定位与授时(2020年5期)2020-09-23
铁道通信信号(2020年9期)2020-02-06
知识经济·中国直销(2018年3期)2018-04-12
中国中药杂志(2016年20期)2016-11-19
江苏农业科学(2015年5期)2015-10-20
江苏农业科学(2015年9期)2015-10-20
江苏农业科学(2015年8期)2015-09-10
