
菌根真菌影响森林生态系统碳循环研究进展
2024-05-06 03:12史加勉
生态学报 2024年7期
杨 浩,史加勉,郑 勇,2,*
1 福建师范大学湿润亚热带生态-地理过程教育部重点实验室, 福州 350117 2 福建三明森林生态系统国家野外科学观测研究站, 三明 365002
1 森林生态系统与碳(C)循环
森林是地球上最大、最重要的生态系统之一,覆盖面积超过4000万平方公里,约占地球陆地总面积的30%[1],是全球生物多样性的重要栖息地。森林生态系统提供多种关键的生态功能,如增加碳(C)固持、保护土壤以及提供木材资源等[2—3]。由于森林土壤中含有大量的有机质,因此森林发挥着极其重要的C汇功能。例如,温带和北方森林每公顷土壤平均每年分别接收12.625 Tg C和5.652 Tg C的凋落物[4];而热带和亚热带森林约占世界森林C汇总量的70%[3]。除了调节气候、保持水土、维持生物多样性等常规的生态功能,森林通过消纳大气中的二氧化碳(CO2),进而减缓温室效应,这在人口密度较高的热带和亚热带地区尤为重要[5]。在森林生态系统中,涉及元素周转和有机物分解等生态过程,均与植物及其相关的微生物组(microbiome)有着密不可分的联系。
地下菌根是植物根系与土壤真菌形成的互惠共生体,真菌帮助植物获取养分和水分,而植物为真菌提供碳水化合物[6]。根据形态学差异,菌根可分为七种主要的类型,其中丛枝菌根(arbuscular mycorrhiza, AM)和外生菌根(ectomycorrhiza, EcM)是分布最广泛的两种菌根类型[7]。在森林生态系统中,几乎所有树木其根系都能与土壤真菌形成AM或EcM,进而调节森林土壤C、氮(N)、磷(P)等元素循环过程。研究表明,土壤C储量与菌根真菌类型密切相关。例如,Averill等[8]指出,相较于AM树种为主的森林,EcM和欧石楠菌根(ErM)树种为主的森林每单位N土壤中容纳的C要高70%,主要由于后者可以产生N降解酶系,使得有机N源可以更多地被真菌和宿主吸收利用,相对而言,土壤中起分解作用的腐生性真菌可利用的N源减少,因此土壤有机质分解受限,土壤C更多地被土壤所固持。Jo等[9]的研究发现,AM树种多度增加时,总体上加速了土壤养分的周转,进而影响森林生产力、生态系统C和养分保持,即AM植物主导的森林其养分循环加速。也有研究通过田间和盆栽实验,发现植物在接种AM真菌后产生的球囊霉素相关土壤蛋白(GRSP)可以提高土壤团聚体稳定性参数[10—11]。Terrer等[12]发现,大气CO2浓度升高(eCO2)时,植物和土壤在C贮存方面存在权衡关系:当植物地上生物量在eCO2条件下增加时,土壤有机C储量却会下降。值得注意的是,与AM森林相比,EcM森林在eCO2的影响之下,植物对N吸收更多,土壤有机质(SOM)分解作用更强,导致土壤释放更多的C,反倒不利于土壤C固持。因此综合来看,AM森林和EcM森林究竟谁可以更多地固持C,目前尚无定论。
为了深入理解和梳理森林中菌根真菌及其在C循环功能方面的研究进展,本文概述了温带(或北方)森林以及热带和亚热带森林中菌根真菌在土壤C的稳定、分解、积累方面的影响、作用机制及其对全球变化的响应,并对该研究领域可能存在的主要问题与今后的研究重点进行展望。
2 温带/北方森林中的菌根真菌与C循环
温带森林和北方(寒带)森林生态系统主要分布在亚洲北部、欧洲大部及北美洲北部。在全球范围内,北方森林覆盖了约11%的陆地表面积[13],其中土壤的C储量达到整个森林C储量的16%[14]。地上植物凋落物的质量和分解速率被认为是SOM长期积累的决定因素。大量光合固定的C直接作用于地下根系及其相关微生物[15],可能对土壤C的吸存产生重要影响[16—17]。因此从菌根真菌角度去探究地下C的去向及其固存速率的影响因素,对于预测温带森林和北方森林生态系统中土壤C储量在应对森林管理策略改变及全球气候变化时如何响应至关重要。
2.1 温带/北方森林生态系统菌根真菌对C贮存的影响
温带/北方森林是以EcM树种为主要植被的森林生态系统(图1),但其地上生物量C总量仅占森林生态系统总生物量的21%,表明EcM植被对地上生物量C的贡献相对较小。相反,地下C储量与EcM植物生物量的比例呈正相关。Phillips等[18]认为这些地区之所以拥有大量地下C储量主要是由于树木根系向菌根真菌提供了大量的C,进一步强调了菌根分解过程相对缓慢,森林地下C储量占主导。也就是,菌根共生体占温带(北方)森林地下C的很大一部分。森林中的优势植物物种将相当大比例的地下C分配给菌根共生体,这些共生体代表了SOM总库中大量的C输入[19—21]。事实上,北方森林腐殖质层中积累的C至少有一半来自根系输入,而不是地上植物凋落物输入[22]。土壤真菌可以促进宿主植物通过光合作用固定更多的大气CO2。随后在植物的根际,一部分C转移至菌根真菌,用于菌根真菌的生长和菌丝等组织发育,一旦这些菌丝死亡,其组织中的C可迅速被其它土壤微生物所分解,或者保存在土壤中多年,甚至数十年[23]。菌根相关的C保留在土壤中的时间越长,对土壤C封存的可能贡献就越大。长期的森林演替和随之而来的腐殖质积累与菌根真菌群落的系统发育及形态变化是一致的。与根相关的子囊菌门真菌通常能促进来自菌丝体的有机C和N的生化稳定,某些形成线状菌根的真菌在演替早期的多度较高,这意味着菌根真菌能从新近产生的真菌菌丝和旧的腐殖质中都能有效地回收N和C[24]。基于上述研究结果,我们可以发现,菌根真菌在北方或温带森林生态系统C汇方面的确起着至关重要的作用,而全球气候变化对真菌群落有着复杂的影响,从而将影响全球C元素的生物地球化学循环[25]。
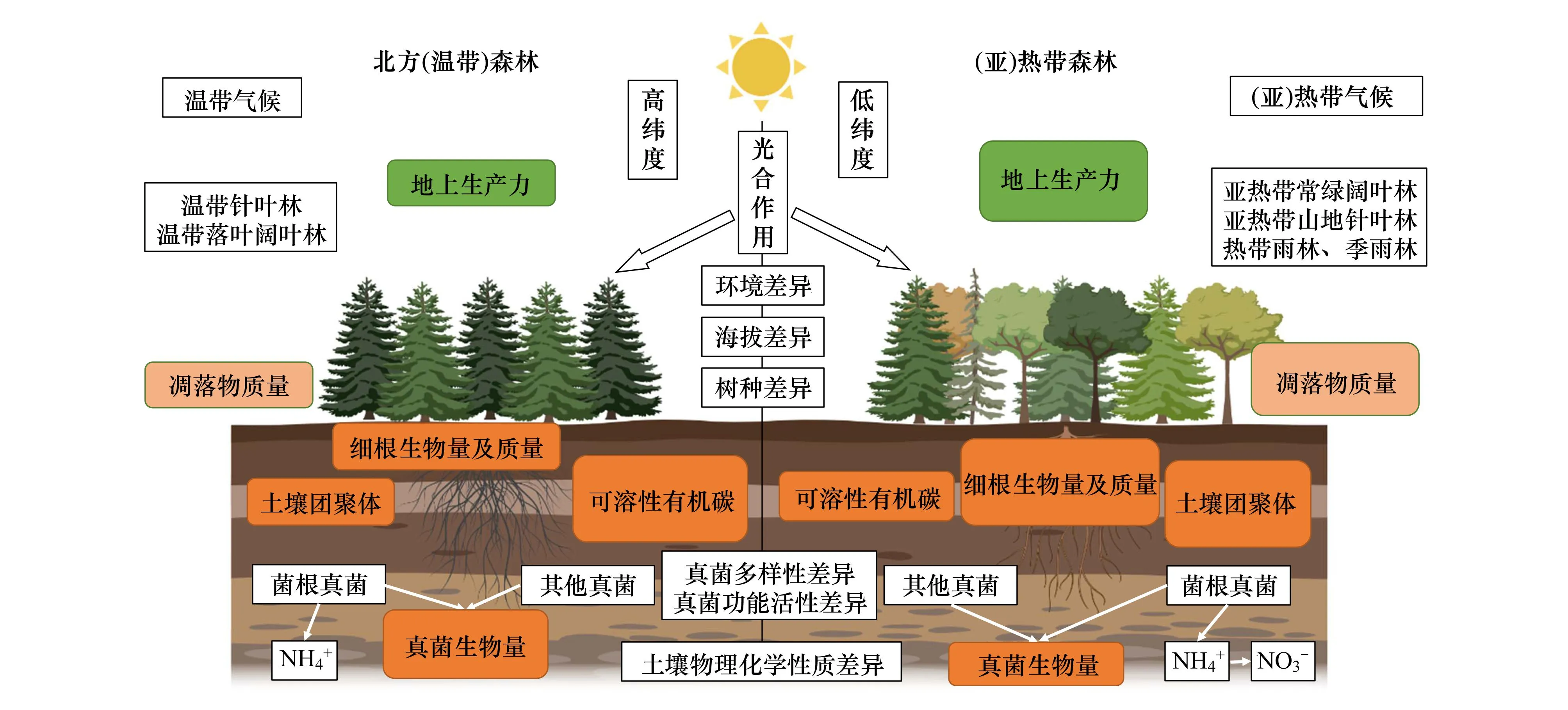
图1 不同类型森林生态系统中菌根真菌及其他微生物介导的土壤C循环的差异Fig.1 Differences in soil C cycling mediated by fungi across different forest ecosystems图中彩色矩形的大小代表其相应参数的大小或质量的高低,不同菌根真菌关于N循环的差异同时会造成其C循环的部分差异
2.2 全球变化对温带/北方森林生态系统菌根真菌和C库的影响
揭示全球变化因子对森林生态系统C存贮或分解的影响及相关机制至关重要。近年来,已经陆续有研究表明大气CO2富集[26—27]、N沉降[28—29]、土地利用变化[30—31]和温室效应[32—33]以及干旱[34—35]等气候变化因子,是潜在影响森林生态系统C稳定或分解的重要因素。例如,随着大气CO2浓度的升高,菌根真菌可能会从土壤中活体或已死亡的菌丝生物量中吸收更多的C,从而对化石燃料燃烧和森林砍伐造成的大气CO2浓度上升起到了缓解作用。另一方面,大气N沉降可能增加真菌组织的周转速率,并消除CO2对菌丝生物量的影响从而导致菌丝中的C储量下降。
AM真菌和EcM真菌的群落组成随着CO2或N的增加而变化,菌根的生长速度和菌丝质量不同,且不同的植物物种组合可能会对菌根生物量、存续时间或分解速率产生综合的影响(图1)[36]。Hanson等[37]认为,在全球变化背景下,较高的真菌多样性可能是维持森林生态系统稳定的必要条件。氮富集显著影响森林土壤中真菌的多样性及其C汇功能,例如向北方森林生态系统施加过量N时,地下C通量显著减少[38]。Phillips等[18]研究发现,随着大气CO2浓度升高,来自根系的C其分解速率加快,相当程度上抵消了地下C输入的增加。Clemmensen等[22]研究发现北方森林生态系统长期演替时,菌根的N循环受到负面影响,从而导致植被养分受限和组成发生变化[39—40]。在全球变化背景下,增温导致的森林土壤变暖会对土壤呼吸产生促进作用,研究发现增温条件下EcM真菌对土壤呼吸有显著影响[41],且增温导致的植物对EcM真菌C分配的变化可能会改变EcM真菌与自由生活的分解者之间竞争相互作用的强度[22,42]。例如,Morrison等[43]发现土壤增温增加了凋落物中木质素的相对多度比例达23%,提高了木质素分解酶的活性,抑制了纤维素分解酶的活性,且发现其中外生菌根的多度是对照的三倍。Jörgensen等[44]针对北方森林的研究表明,施N增加了真菌生物量,促进了土壤C固持,但减少了EcM真菌的数量,一定程度上破坏了该生态系统中有机质的营养循环。
3 热带/亚热带森林中的菌根真菌与C循环
热带和亚热带森林是全球森林生态系统中除了温带、北方森林之外的另一重要组成部分。热带森林分布在地球温暖、潮湿的赤道地区,占据了一条宽阔的区域带[45]。该区域的植物主要以壳斗科、樟科、竹林与针叶林(如马尾松、杉木林)为主(图1)。在全球尺度上,热带和温带森林的C储量密度是相当的(242 vs. 239 Mg C hm-2)。相对于温带或北方森林,热带/亚热带森林以生物量形式存储的C更多(达56%),而仅有32%储存在森林土壤中[3]。
3.1 (亚)热带森林生态系统菌根真菌对C库的影响
与温带(北方)森林类似,在热带和亚热带森林生态系统,菌根同样是影响C稳定或分解的重要因素。热带森林主要以AM树种为主(图1),热带森林地上生物量为162 GT,占44%,显著高于温带/北方森林的21%,表明AM植被对热带森林地上生物量C的贡献相对较大[25]。Rillig等(2004)的研究表明,AM真菌通过菌丝产生的糖蛋白对SOM的贡献不容被忽视,表明那些非源于凋落物的菌根真菌相关的C对土壤有机C库和N库的贡献甚至可能超过土壤微生物生物量的贡献[46]。Bereau等[47]的研究发现,在水分充足条件下,菌根定殖可以提高宿主植物净光合作用、生长速率和P的吸收,而当土壤湿度到达某一阈值,菌根共生降低了双柱苏木(Dicoryniaguianensis)植物幼苗的C的吸收,这可能与植物和菌根真菌在低光照强度下的C分配竞争有关。另有研究发现,AM真菌的根外菌丝是湿润热带森林土壤CO2的主要释放源,表明AM真菌根外菌丝是C从热带森林树木到大气的重要途径,通过它们的高呼吸速率将植物来源的C迅速释放到大气[48]。在热带森林中,尽管AM真菌其菌丝本身对有机质矿化的促进作用有限,但它们可作为重要通道将C传递给其他土壤微生物[49]。近期的研究发现,在亚热带落叶和常绿阔叶林中,AM树种比EcM树种具有更强的降解可溶性有机碳(DOC)的能力,但AM和EcM两种常绿针叶树其凋落物中DOC的生物降解速率无显著差异[50]。总而言之,研究菌根真菌与植物之间的相互作用,厘清菌根介导C元素固定或分解的机理,是目前生态学较为热门和必要的研究课题。
3.2 全球变化对热带/亚热带森林中菌根真菌及其C库的影响
全球变化对热带、亚热带森林中的菌根真菌的影响不尽相同。例如,在热带森林生态系统,对于土壤真菌群落的影响,降水可能是比植物多样性或土壤养分更重要的因素,土壤中菌根真菌类群数量随年均降水量的增加而显著增加[51]。此外从全球尺度来看,大气N沉降或施氮肥可能会降低EcM真菌的生物多样性,改变EcM真菌的群落组成[52]。Vincent和Declerck[53]研究了全球变化对EcM真菌可能产生的影响,认为eCO2可能会增加树木的C固存,最终导致EcM真菌的菌丝生物量、群落组成发生变化,最终对eCO2产生正反馈。EcM真菌对增温或全球变暖的响应相对微弱,而且是以种水平产生特异性差异的,其可能是通过改变养分循环,从而影响整个生态系统[54]。比较而言,EcM森林对全球气候变化的适应能力更强,而AM森林可能对全球气候变化相对更为敏感[55]。例如,Maitra等[56]发现在亚热带森林中,虽然干旱不会对土壤和根系中AM真菌OTU丰富度有显著影响,但是干旱会显著降低AM真菌根外菌丝密度、孢子密度和根侵染率。同样,热带森林的AM真菌对N输入的响应都表现为根侵染率、孢子密度和根外菌丝长度的强烈下降[57]。全球气候变化背景下,AM和EcM真菌群落的响应变化,会进一步影响森林C循环相关功能。Averill等[28]预测:①生态系统的无机N输入将有利于AM相关的树种,而不利于EcM相关的树种;②无机氮输入导致的森林中EcM真菌的下降将减少森林生物群落的地下C储存能力。Treseder[58]提倡在地球系统模型中更详细地纳入AM真菌,以改进我们对全球气候变化背景下的森林C储量的预测。总而言之,全球变化对于森林C循环有着复杂的影响,这就需要我们对其中真菌介导的森林C汇功能进行机理分析。
4 菌根真菌驱动森林生态系统C循环功能的机制
4.1 菌根真菌与森林地下C的关系
凋落物的分解与森林生态系统C汇潜力有着直接的关系。早期研究认为,凋落物的分解速率一般是由气候、基质质量及其化学性质所决定[59—60]。后来研究发现,微生物群落组成也是影响凋落物分解的重要因素[61—63]。例如,基于“养分竞争假说”的加吉尔效应(Gadgil effect)认为,土壤中的菌根真菌与其他微生物都需要可利用养分以供应自身的生长和繁殖,两者的竞争关系导致了其他微生物的N限制,从而抑制了SOM的分解。Smith和Wan[64]应用资源比率理论(Resource-ratio theory)评估了真菌对不同形式有机N的竞争是否会影响凋落物的分解,结果发现EcM真菌相比于自由腐生真菌,在分解那些难分解凋落物时存在养分竞争方面的优势,只有在那些难分解有机物占主导的森林中,才会产生加吉尔效应[65—66]。虽然EcM真菌并不总是限制凋落物的分解,但EcM对N的吸收往往增加凋落物的碳氮比(C/N),这就间接地增大了凋落物的分解难度。因此,在EcM植物占主导的森林生态系统中,由于土壤C/N相对更高,加吉尔效应对土壤有机质分解的抑制作用可能比先前预测的更大。
SOM的形成与凋落物分解亦息息相关。Cotrufo等[67]提出的“微生物效率-基质稳定(microbial efficiency-matrix stabilization)假说”认为,不稳定的植物成分能被微生物更有效地利用,因此它们是微生物残体C的主要来源,这些通过微生物分解的产物通过促进聚集作用、与矿物土壤基质的强化学键结合作用等,成为稳定型SOM的主要前体。此外,SOM的形成还与凋落物质量、C饱和状态等有直接的联系[68]。不同树种及其菌根类型对于土壤C循环的影响存在显著差异。例如,有别于EcM真菌,AM真菌无法直接利用土壤中的有机养分,而是偏向于利用“无机”营养模式,具有更快的有机质周转速率[69]。另有“缓慢分解(slow decay)假说”指出,EcM真菌系统中的营养循环相对封闭,凋落物分解速率更慢,因而植物源有机C在土壤中的积累相对容易[70—71]。上述的“资源比率理论”和“微生物效率-基质稳定”两种假说都存在于森林生态系统中,它们共同说明了有机C在表层土壤中的分解速率,但对深层土壤中有机C的储存可能有着不同的影响[72]。
地下植物C输入主要通过根系-真菌途径进入土壤。真菌是主要的分解者以及森林生态系统中微生物源SOM的主要贡献者之一[73]。EcM真菌是地下植物C的关键影响因素,高达35%的净初级产量通过宿主根系分配给EcM[74]。地上植物凋落物被认为是EcM的重要营养来源,特别是N元素影响它们的多度和群落组成[23,75]。Whalen等[76]的研究指出,根系相比于凋落物对土壤中真菌群落生物量、活性以及群落组成等影响更大。去除根系后,土壤微生物总生物量、土壤C储量显著减少,并且显著降低了土壤真菌群落中特定类群的相对多度,例如EcM真菌中的Russula以及病原菌和腐生菌中的Ascomycota,同时根系去除会降低真菌的多度和群落构建的速度。
4.2 不同菌根类型树种对于土壤C循环的影响机制
一般认为,森林土壤总C主要来自地上植物凋落物输入和地下根际输入,地上和地下两种输入途径对土壤C的贡献因森林生态系统类型而异。研究发现,北方森林表土腐殖质层累积的C主要来自地上植物凋落物输入以及腐生菌的分解作用,而更深层土累积的C来自于菌根真菌的贡献[22]。然而,在温带森林生态系统中新近的研究发现,森林C汇功能还依赖于地上植被类型与地下的真菌类型。例如,研究表明处在不同演替阶段的森林生态系统其C汇潜力与森林土壤真菌多样性之间密切相关[24]。在森林演替早期,地下EcM真菌占优势,土壤固持的C较少。及至森林演替后期,地上植被变化且伴随地下ErM真菌逐渐成为主导类群,其产生的黑化菌丝难以被分解,促进了腐殖质的形成,土壤C固持功能变得更强。此外,菌根真菌在SOM的形成与稳定过程中起重要作用[74]。一方面,菌根真菌可以通过直接的酶分解、化学氧化、以及供C方式促进其自身的异养生长,从而加速SOM的降解;另一方面,菌根真菌会与自由生长的腐生微生物发生资源竞争关系,从而潜在地抑制SOM的分解,导致更多土壤C的累积[74]。
菌根真菌在陆地生态系统中对土壤C循环的影响主要有以下三种方式:①通过导管(conduit)形式把植物光合作用固定的产物转运到土壤,从而增加植物-土壤途径的C输入[49,77—78];②菌根共生体本身生物量对土壤中有机C的贡献及其分泌土壤蛋白物质所发挥的C汇功能[79—80];③通过激发效应或控制土壤养分有效地调控土壤有机C的矿化[23,61,81]。AM和EcM无论是在形态上还是在生理功能上都存在明显差异,因此菌根介导的森林土壤C循环很大程度上取决于(优势)树木的菌根类型和森林土壤中菌根真菌的群落组成[8,82]。树种菌根类型对于森林土壤C的影响主要有如下几个方面。
首先,不同菌根类型的树种在对宿主植物光合产物分配方面存在差异。EcM植物通常可分配更多的碳水化合物给菌根真菌[83],使得EcM植物比AM植物通过菌根释放到土壤中的C更多。AM和EcM两种共生体在生产力[84—87]、菌根形成[88]以及细根结构[89—90]上都存在差异,这些差别影响到它们在地上生产力和地下C汇功能的贡献度。
其次,不同菌根真菌产生的根外菌丝的数量及其功能存在差别。菌根真菌的根外菌丝是土壤中重要的C源,通常认为EcM真菌产生的根外菌丝生物量要比AM真菌高出一个数量级[82],但是AM真菌具有更加丰富的宿主植物物种[91],可以形成不易分解的有机C,产生球囊霉素相关土壤蛋白[46]等,使得AM真菌生物量同样是土壤有机C输入的重要组分。EcM宿主凋落物比AM宿主凋落物的分解速率慢1倍以上[86—87],在以EcM树种为优势树种的森林中会存在较多半分解的凋落物残体[8,69],其土壤表层会有更多凋落物C积累[92]。Lin等[93]通过盆栽试验,评价了活根和菌根真菌菌丝对其自身凋落物分解的相对影响,发现菌根真菌菌丝能显著降低凋落物质量损失,但以菌根类型对不同树种进行分类时,EcM树种和AM菌树种的活根和菌根真菌对凋落物分解的影响差异不显著。
第三,不同菌根类型树种影响土壤微生物的代谢活性。土壤中与微生物C和N转化相关的胞外酶如β-1, 4-葡糖苷酶(BG)、β-N-乙酰-氨基葡糖苷酶(NAG)活性以及微生物代谢熵(qCO2,即单位微生物生物量的呼吸速率)在不同菌根类型森林生态系统中存在明显差异。例如,温带和亚热带AM树种占主导的森林生态系统其土壤BG酶活性显著高于EcM树种占主导的森林[94—95],而不同菌根类型森林土壤NAG酶活性显著不同[96]。单位微生物生物量下土壤BG和NAG酶活性表现为EcM森林显著高于AM森林[97],说明在EcM占主导的森林生态系统,微生物对土壤胞外酶的投资更大,并且微生物对于合成获取N的胞外酶的投资大于C相关胞外酶[95,97]。类似地,经微生物生物量标准化的qCO2表现为EcM森林显著高于AM森林[95,98],表明在EcM森林,土壤微生物的C利用效率更低[99—100]。因此,不同菌根树种主导的森林生态系统中,土壤微生物功能活性存在差异,会影响土壤养分状况以及共生树种的生长,随后土壤微生物和地上植物共同影响和驱动森林土壤C循环。
5 存在的问题及未来研究展望
菌根真菌作为森林生态系统地下最重要的微生物功能类群组分,是森林生态系统土壤C循环和贮存的关键影响因子。随着分子生物学技术的发展,人们已经对森林生态系统真菌组开展了较多的基础性研究,明确了真菌多样性和群落组成对于维系森林生态系统地上和地下有机物分解、C转化、C平衡方面的关键作用。然而,菌根真菌对森林C汇功能特别是土壤C库变化的影响机制和内在作用机理还缺乏系统深入的认知,建议未来的研究应重视如下几个方面:
应加强基于野外长期控制实验的关于菌根类型对森林C汇功能实验研究。例如,作为世界上最大的生物多样性实验网络平台,TreeDivNet(https://treedivnet.ugent.be/)目前汇聚了全球29个实验,植树总数超过120万株;该平台致力于通过长期定位试验,研究树种多样性(包括菌根类型)与森林生态系统功能之间的关系。我们于2021年在福建上杭建立了“树种菌根类型与森林生态过程长期试验平台”,拟开展亚热带森林典型AM和EcM树种组合如何影响土壤生物多样性及森林C汇功能方面的研究。
其次需要进一步发展菌根真菌多样性的研究方法与技术。例如在二代测序技术的基础上,有待三代测序技术其测序精度提高后,更广泛地应用于森林生态系统菌根真菌多样性的检测。三代测序在读长方面的优势,无疑将显著提高菌根真菌物种分子鉴定方面的准确性,有助于甄别参与森林生态系统C循环功能的真菌物种或类群。然而在实践中,仍然需要重视菌根真菌的形态学研究,如菌丝、孢子、侵染强度等性状往往是响应环境因子的敏感指标,因此在采用分子生物学研究方法的同时,同样需要重视发展形态学研究方法与技术。
全球变化对菌根真菌及森林土壤C汇功能的影响研究有待加强。空间尺度方面,全球变化对于森林生态系统及其菌根真菌的影响较为复杂,结果往往会因研究地点、森林类型、树种特性等不同而差异明显。时间尺度上,由于全球变化可视作一种长期性的环境压力,森林生态系统中树木、菌根真菌及其功能会存在一种逐渐的适应性或耐受力,因此短期响应与反馈可能不足以准确体现全球变化对于森林生态系统的影响。总之应加强不同时空尺度下,森林生态系统中菌根真菌多样性及其生态功能对全球变化的响应与反馈研究。
猜你喜欢
陕西农业科学(2022年2期)2022-03-28
浙江农业学报(2021年5期)2021-05-31
园林科技(2020年2期)2020-01-18
上海农业学报(2017年4期)2017-04-10
广东饲料(2016年5期)2016-12-01
亚热带资源与环境学报(2015年4期)2015-01-22
东北师大学报(自然科学版)(2014年1期)2014-02-27
亚热带资源与环境学报(2014年3期)2014-01-23
环境与可持续发展(2013年6期)2013-03-11
植物营养与肥料学报(2012年5期)2012-10-26
