
巨噬细胞迁移抑制因子调控人胚胎干细胞增殖的相关性分析
2024-05-14 17:30冯晓丽陈薪旭郑晓晗
健康之家 2024年2期
关键词:增殖
冯晓丽 陈薪旭 郑晓晗
摘要:目的 明确巨噬细胞迁移抑制因子在胚胎干细胞上的相互作用受体,阐明MIF维持胚胎干细胞增殖存活的作用机制。方法:培养人胚胎干细胞,运用流式细胞仪、免疫荧光共聚焦显微镜、Western blot等检测方法检测HESCs相关受体CD74、CD44、CXCR2、CXCR4、CXCR7 表达情况,同时在胚胎干细胞中加入MIF、ISO后,CCK-8 法检测人胚胎干细胞活性。结果 流式细胞仪、免疫荧光共聚焦显微镜、Western blot等检测方法表明,人胚胎干细胞中的相关因子均有表达,其中CXCR2、CXCR7和MIF的表达水平较高,而CD74、CD44、CXCR4表达水平较低,但比较并无显著差异,P>0.05。在胚胎干细胞中加入MIF、ISO后,CCK-8 法检测显示,与空白组相比,对照组和实验组的吸光度值和细胞活性均明显下降,P<0.05;与对照组相比,实验组的吸光度值和细胞活性明显更低,P<0.05。结论 MIF在人胚胎干细胞中可能通过与CXCR2、CXCR7、CD74、CD44、CXCR4等相关因子结合调控人胚胎干细胞的增殖活性。
关键词:巨噬细胞迁移抑制因子;人胚胎干细胞;增殖;细胞活性
目前研究发现,人胚胎干细胞(hESCs)可分泌表达MIF,并在维持hESCs增殖存活方面具有重要作用[1~2]。MIF促进hESCs增殖存活的相关功能機制仍不完全明确,相关研究认为MIF可能是通过结合CXCR2和CXCR7相关受体,进而激活下游PI3-Akt信号通路来维持ESCs的增殖和存活[3~4]。为此,本研究拟通过添加不同浓度梯度的MIF、MIF抑制剂ISO观察ESCs的增殖存活状况,检测ESCs 中MIF的自分泌情况、相关受体表达,以及不同条件下细胞增殖情况,阐明MIF维持ESCs增殖存活的相关机制。
1资料与方法
1.1 临床资料
检验方法:包括酶联免疫吸附测定法、CCK-8法、免疫细胞化学染色、流式细胞仪分析、蛋白质印迹法、免疫共沉淀等方法。
细胞培养:胚胎干细胞(H9)在添加 mTeSRTM1培养基中,置于37 ℃、5% CO2和饱和湿度的培养箱中常规培养,取对数生长期且状态良好的细胞用于后续实验。贴壁细胞消化,收集细胞悬液,离心收取细胞上清用于后续检测。
1.2 方法
(1)流式细胞仪检测H9细胞CD74、CD44、CXCR2、CXCR4、CXCR7表达情况:将六孔细胞培养板中的H9细胞制成单细胞悬液。取100 ?L细胞悬液至离心管中,并分别在管中加入5 ?LPE标记的CD74、CD44、CXCR2、CXCR4、CXCR7对应抗体,4 ℃反应30 min;加入2 mL PBS离心去上清,重复2次;再加入500 ?LPBS,通过流式细胞仪器进行检测。
(2)免疫荧光共聚焦显微镜检测H9细胞MIF、CD74、CD44、CXCR2、CXCR4、CXCR7表达情况:取对数生长期的H9细胞。长满后,加入750 ?L/9.6 cm2的Accutase进行消化;接种于细胞玻片的24孔细胞培养板内,待细胞贴壁,用 4%的多聚甲醛室温下固定20 min;用 PBS清洗3次后,同时用10%驴血清(Biorad)和 0.1%Triton X-100(Sigma)的PBS在室温下封闭和渗透30 min。孵育一抗4 ℃过夜。用PBS洗3次,每次5 min。避光孵育二抗1 h后,再用PBS洗3次,每次5 min。检测MIF、CD74、CD44、CXCR2、CXCR4、CXCR7表达情况。本阶段所用试剂如下。
一抗:鼠抗人MIF(1:100); 兔抗人CD74(1:200);兔抗人CD44(1:100);兔抗人CXCR2(1:200);兔抗人CXCR4(1:200);兔抗人CXCR7(1:200)二抗:山羊抗鼠(1:500), 山羊抗兔(1:500);样品用抗褪色荧光固定介质(Dako)固定。使用蔡司 LSM 510 共焦激光扫描显微镜(Carl-Zeiss 显微成像)获取图像。
(3)Western blot检测H9细胞MIF、CD74、CD44、CXCR2、CXCR4、CXCR7表达情况:胚胎干细胞(H9)在添加 mTeSR?1培养基中,置于37 ℃、5% CO2和饱和湿度的培养箱中常规培养。长满后,加入750 ?L/9.6 cm2的Accutase进行消化;细胞悬液离心,收集细胞沉淀;加入细胞裂解液和蛋白酶抑制剂裂解细胞,离心提取总蛋白;BCA 法测定蛋白浓度,加样后采用80 V 30 min,120 V 60min程序进行电泳,再通过湿转法将蛋白转移到PVDF膜上。转膜完成后,PVDF膜用封闭液室温孵育2 h。再将PVDF膜与特异性一抗在4 ℃下过夜孵育,TBST洗5次。PVDF膜置于特异性二抗中室温摇床孵育2 h,TBST洗5次,滴加ECL发光液进行蛋白曝光。
(4)加入MIF、CCK-8 法检测细胞活性:取对数生长期的H9细胞。长满后,加入750 ?L/9.6 cm2的Accutase进行消化, 接种于96孔细胞培养板,共分3组,分别为空白组、对照组、实验组,每组重复3个复孔。空白组加入包被培养器皿的BD MatrigelTM 和500 ?LmTeSRTM1培养基,对照组加入包被培养器皿的BD MatrigelTM 和100 ?LmTeSRTM1培养基以及实验组等量细胞,实验组加入包被培养器皿的BD MatrigelTM和100 ?LmTeSRTM1培养基以及适量细胞。再分梯度浓度加入hMIF,各组在37 ℃、5% CO2和饱和湿度的培养箱中常规培养,分别在24 h、48 h、72 h每孔细胞加入10 ?L CCK-8溶液,共孵1~4 h后在酶标仪450 nm处读取每孔细胞的吸光度值,测定细胞活力。
(5)加入ISO、CCK-8 法检测细胞活性:操作步骤同(4)。
1.3 统计学分析
研究所得数据均采用SPSS27.0 统计软件统计处理,数据以(±s)表示。所有数据均采用 GraphPad Prism软件进行显著性检验,采用 Bonferroni 事后检验的单因素方差分析。P<0.05为差异性显著。
2结果
2.1 不同检测方法下人胚胎干细胞相关因子的表达水平
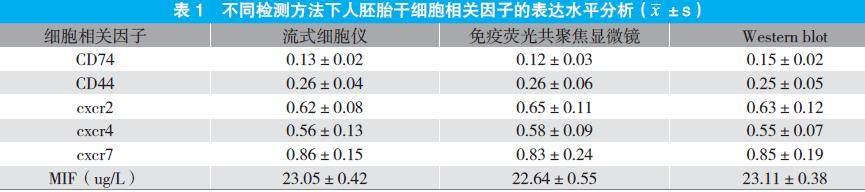
流式细胞仪、免疫荧光共聚焦显微镜、Western blot等检测方法表明,人胚胎干细胞中的相关因子均有表达,其中CXCR2、CXCR7和MIF的表达水平较高,而CD74、CD44、CXCR4表达水平较低,但比较并无显著差异,P>0.05。见表1。
2.2 胚胎干细胞活性检测
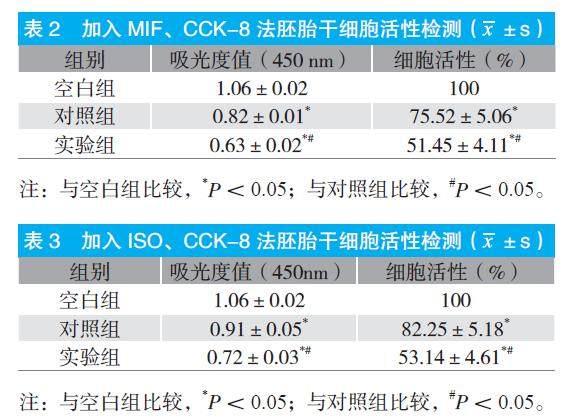
在胚胎干细胞中加入MIF、ISO后,CCK-8 法检测显示,与空白组相比,对照组和实验组的吸光度值和细胞活性均明显下降,P<0.05;与对照组相比,实验组吸光度值和细胞活性明显更低,P<0.05。见表2~3。
3讨论
本研究发现,流式细胞仪、免疫荧光共聚焦显微镜、Western blot等检测方法表明,人胚胎干细胞中的相关因子均有表達,其中CXCR2、CXCR7和MIF的表达水平较高,而CD74、CD44、CXCR4表达水平较低,但比较并无显著差异,P>0.05。这些因子可与MIF结合,其中CD74在MIF受体复合物中起着关键作用,因为CD74敲除或应用抗CD74抗体可消除表达CD44、CXCR2、CXCR4或CXCR7的细胞中的MIF效应[5]。但有研究表明,MIF可通过CXCR7激活PI3K-AKT信号通路。这说明CD74在介导MIF信号中的作用中并非完全不可或缺。CD74不能单独介导MIF信号,但是可以协助增强CD44、CXCR2或CXCR4的表达效应[6]。因此,CD74在MIF信号中起着中枢作用,可以引导组织受体并帮助形成受体复合物。MIF可以直接结合CXCR2和CXCR7受体触发相关效应。MIF首先与CXCR2结合,通过与G蛋白偶联激活ERK1/2和AKT信号通路。CXCR7不与任何类型的G蛋白偶联,而是与β-arrestin2结合进而激活ERK1/2信号通路,但也有研究显示可以同样激活AKT信号通路[7~8]。此外,MIF 最重要的作用之一是具有促进细胞增殖和抑制细胞凋亡的能力。ISO-1经常作为MIF的标准抑制剂,可抑制MIF的互变异构酶活性,也可阻止 MIF 与其表面受体的结合,从而阻断MIF诱导的信号级联[9~10]。本研究在胚胎干细胞中加入MIF、ISO后,CCK-8法检测显示,与空白组相比,对照组和实验组的吸光度值和细胞活性均明显下降,P<0.05;与对照组相比,实验组的吸光度值和细胞活性明显更低,P<0.05。表明抑制 MIF 将会显著抑制细胞的增殖活性。
综上所述,CXCR2、CXCR7、CD74、CD44、CXCR4等MIF相关受体在人胚胎干细胞中均有所表达,表明巨噬细胞迁移抑制因子在调控人胚胎干细胞增殖中具有重要作用。
参考文献
[1]李江,夏鹏,曹必伟,等.独活对巨噬细胞迁移抑制因子抑制作用的有效成分虚拟筛选研究[J].中国中医药信息杂志,2019,26(5):105-108.
[2]SPARKES A,De BAETSELIER P,ROELANTS K,et al.The non-mammalian MIF superfamily[J].Immunobiology,2017,222(3): 473-482.
[3]杜向荣,宋守君,薛海波,等.中国汉族人中巨噬细胞移动抑制因子-173G/C基因多态性与免疫相关性疾病易感性的关系:Meta分析[J].中华微生物学和免疫学杂志,2018,38(8):582-590.
[4]李玉锦,王杰,王亚妮,等.巨噬细胞移动抑制因子调控细胞衰老的研究进展[J].中国生物工程杂志,2023,43(2):120-129.
[5]郑晓晗,冯晓丽,胡兰,等.巨噬细胞迁移抑制因子促进人胚胎干细胞分化为腹侧中脑多巴胺能神经祖细胞[J].中国组织工程研究,2023,27(33):5348-5356.
[6]魏艳召,郑晓晗,高仕君,等.人胚胎干细胞自分泌巨噬细胞迁移抑制因子及受体表达[J].中国组织工程研究,2023,27(1):34-41.
[7]肖农,曹雪涛,于益芝,等.重组腺病毒介导的TNF基因转染对巨噬细胞功能影响[J].中国肿瘤生物治疗杂志,1995(4):295-296.
[8]陈天琰,张磊,高建一,等.人胚胎干细胞源神经干细胞微囊泡对巨噬细胞极化的影响[J].江苏大学学报(医学版),2020,30(6):515-519,525.
[9]元力,潘旭,边国慧,等.不同体外诱导分化来源巨噬细胞表型分子分析及极化功能比较[J].国际输血及血液学杂志,2018,41(2):121-132.
[10]韦添元.以CRISPRa诱导人胚胎干细胞分化为巨噬细胞的研究[D].哈尔滨:哈尔滨工业大学,2022.
猜你喜欢
中国当代医药(2016年32期)2017-02-23
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
山东农业科学(2016年12期)2017-01-21
中国现代医生(2016年27期)2016-12-21
中国医药导报(2016年25期)2016-11-30
科技视界(2016年5期)2016-02-22
中国现代医生(2015年21期)2015-08-07
