
甜菜BvCK2基因全长cDNA的克隆和序列分析
2009-04-19 10:15鲁振强潘彦遥陈连江朱延明李春宇
中国糖料 2009年1期
鲁振强,潘彦遥,陈连江,朱延明,李春宇
(1.黑龙江大学生命科学学院,哈尔滨150080;2.黑龙江大学农作物研究院,哈尔滨150080;3.东北农业大学生命科学学院,哈尔滨150030)
甜菜BvCK2基因全长cDNA的克隆和序列分析
鲁振强1,潘彦遥1,陈连江2,朱延明3,李春宇1
(1.黑龙江大学生命科学学院,哈尔滨150080;2.黑龙江大学农作物研究院,哈尔滨150080;3.东北农业大学生命科学学院,哈尔滨150030)
甜菜M14品系在细胞胚胎学和遗传学特征上具有鲜明的无融合生殖现象。为了寻找在这一特殊的生殖过程中的相关基因及其调控作用,实验通过同源克隆及cDNA末端快速扩增(RACE)的方法,首次从甜菜M14品系中克隆了与生殖相关的基因BvCK2(Beta vulgaris casein kinase 2)的全长cDNA序列,长度为1 501 bp,开放阅读框为1 002 bp,编码333个氨基酸。根据氨基酸序列计算蛋白分子量为39.28kDa,pI=8.16。同源比对表明,BvCK2与烟草CK2(GenBank No.AJ437635)的相似度为64.22%,与百合CK2(GenBank No.AF517838)的相似度为65.18%。通过细胞内定位分析,BvCK2所编码的蛋白主要存在于细胞核中。
甜菜;BvCK2;RACE;基因克隆;序列分析
蛋白激酶,也称酪蛋白激酶-2(casein kinase2,CK2),它是一种高度保守的真核细胞中普遍存在的信使非依赖性丝氨酸/苏氨酸蛋白激酶,它在细胞生存、生长、增殖[1]、细胞周期运行[2,3]、信号传导[4]或转录调控[5]及凋亡过程中具有重要的功能,同时还参与癌症发生、病毒源性肿瘤发生以及抗凋亡等病理过程。CK2被认为具有重要的生命功能,在细胞功能调控中处于重要地位。它是由两个催化亚基(α和α′)和两个调节亚基(β和β′)构成的不均一四聚体[6]。其底物既可以是酶也可以是转录因子,因而它可以通过修饰转录因子激活或抑制基因的表达,使得细胞对外来信号做出反应。这些底物在细胞功能的许多方面起重要作用,而且有些还是细胞复杂的信号加工系统的重要组成部分[7,8]。
黑龙江大学郭德栋教授等从1988年开始,通过栽培甜菜(Beta vulgaris)和具有无融合生殖特性的野生种白花甜菜(Beta corolliflora Zoss)进行种间杂交,最终获得并鉴定了甜菜M14品系在细胞胚胎学和遗传学特征上具有鲜明的无融合生殖现象,是克隆无融合生殖基因极为难得的材料[9,10]。以往对于植物CK2的研究主要以拟南芥、水稻等模式植物为材料,本次对于CK2基因在甜菜中的克隆尚属首次,这对于在甜菜中深入了解该基因的作用机理,尤其对研究甜菜生殖发育的分子生物学调控机理具有重要作用。
1 材料与方法
1.1 材料
选用甜菜(Beta vulgaris)M14品系为植物材料,种植于黑龙江大学温室及试验田。
1.2 主要试剂
pMD18-T Vector,胶回收试剂盒,M-MLV RTase cDNA Synthesis Kit,TaKaRa RNA LA PCRTMKit(AMV)Ver.1.1均购自大连宝生物(Takara)公司;5′RACE试剂盒使用SMARTTMPCR cDNA Synthesis Kit (CLONTECH);所用引物由上海生物工程公司合成。
1.3 总RNA的提取及RT-PCR
于甜菜花期阳光较充足的上午进行采样,取约0.6g花蕾用锡纸包好,液氮速冻,-70℃保存备用。用TRIzo1法提取甜菜M14品系花蕾总RNA。根据已发表的其他物种的CK2基因核苷酸序列进行比对,找出其高度同源的序列,设计简并引物:BvCK2-1,BvCK2-2(表1)。取花蕾总RNA 0.6µL,反转录过程参照LA PCRTMKit试剂盒说明书。
PCR反应:逆转录产物10µL,10×LA RNA PCR BufferⅡ4µL,25mM MgC124µL,引物BvCK2-1和BvCK2-2各1µL,ddH2O 31.75µL,TaKaRa LA Taq 0.25µL。扩增程序为:94℃预变性2 min,94℃变性30s,55℃退火30s,72℃延伸1min,30个循环,最后72℃延伸10min。用1%琼脂糖凝胶电泳检测PCR结果。
1.4 3'RACE
根据已克隆的CK2基因的保守片段和真核生物3′末端具有Po1y尾的特性,应用引物设计软件PrimerPrimier5.0设计3′RACE引物:BvCK2-3,M13 Primer M4(表1)。反转录体系同1.3。反应条件:30℃10min,50℃30min,99℃5min,5℃5min。
PCR反应体系为:反转录产物10µL,MgC12(25mM)3µL,10×LA PCR BufferⅡ4µL,灭菌蒸馏水31.75µL,TaKaRa TaqTM0.25µL,M13 Primer M4 0.5µL,BvCK2-3 Primer 0.5µL。扩增程序为:94℃预变性2 min,94℃变性30s,60℃退火30s,72℃延伸1min,30个循环,最后72℃延伸10min。用1%琼脂糖凝胶电泳检测PCR结果。
1.5 5'RACE
根据已克隆的3′RACE序列和引物设计软件PrimerPrimier5.0,在3′RACE序列的5′末端设计两组5′RACE引物:BvCK2-GSP1,BvCK2-GSP2(表1)。
反转录过程:RNA 3µg,5′CDS引物1µL,SMART II A引物1µL。70℃保温2min,冰上冷却2min,加入5×First-Strand buffer 2µL,DTT 1µL,10mM dNTP Mixture 1µL,Reverse Transcriptase 1µL。42℃水浴1.5h。加入TE(Tricine-EDTA)缓冲液100µL。72℃保温7h。
PCR反应体系为:PCR-Grade Water 34.5µL,10×Advantage 2 PCR Buffer 5µL,10mM dNTP Mixture 10µL,50×Advantage 2 Po1ymerase Mix 1µL,5′RACE-Ready cDNA 2.5µL,10×UPM引物5µL,10µM GSP引物1µL。以降落PCR程序进行扩增,94℃30s,72℃3min,5个循环,94℃30s,70℃30s,72℃3min,5个循环,94℃30s,68℃30s,72℃30s,25个循环。用1%琼脂糖凝胶电泳检测PCR结果。
1.6 序列测序拼接及RT-PCR验证和酶切验证
将RT-PCR产物、3′RACE产物及5′RACE第二轮产物分别连接到pMD18-T载体进行测序。采用DNAMAN软件将3′端和5′端的保守区段进行重叠序列拼接,并根据此序列设计引物扩增全长的cDNA序列:BvCK2-4,BvCK2-5(表1)。对PCR验证后的阳性克隆子用HindⅢ进行酶切鉴定。
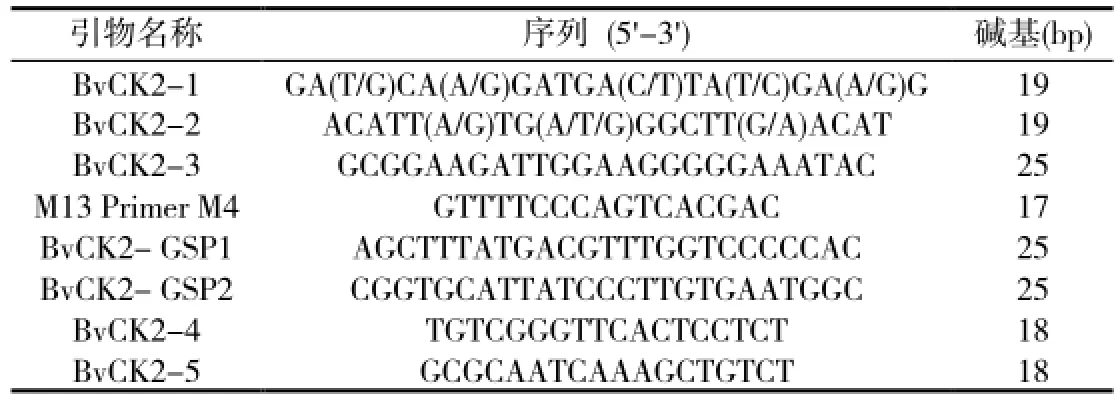
表1 BvCK2扩增引物
2 结果与分析
2.1 甜菜BvCK2全长cDNA序列扩增
提取甜菜M14植株的花蕾RNA,经RT-PCR后,获得383bp的特异条带(图1)。通过B1ast同源比较,发现其与百合(GenBank No.AF517838)、拟南芥(GenBank No.NM-114860)、水稻(GenBank No.AB036788)及烟草(GenBank No.AJ437635)的相应保守片段相似率分别为79.37%、80.79%、80.00%和82.51%,从而初步证实了这条特异条带为目的基因BvCK2的基因片段。为了获得全长基因,实验分别进行了3′RACE、5′RACE扩增。如图2a所示,通过3′RACE扩增,获得了产物大小为1.27kb的片段;同时,5′RACE扩增的结果显示,第一轮扩增并没有获得明显条带。继而取第一轮5′RACE产物稀释10倍后作为巢式PCR的模板,扩增BvCK2基因的5′末端,电泳结果出现大约476bp的条带(图2b)。将已测序的BvCK2 cDNA保守片段3′和5′末端片段互相核对,寻找互相重叠的片段从而拼接出全长cDNA序列。为了进一步确定所获得基因的正确性,实验分别进行了PCR和酶切检测,如图3所示,PCR扩增后在1300bp位置获得了目的条带(图3a);而由于BvCK2全序列上有一个HindⅢ酶切位点,因而经酶切鉴定后分别获得了800bp和500bp左右的片段(图3b)。
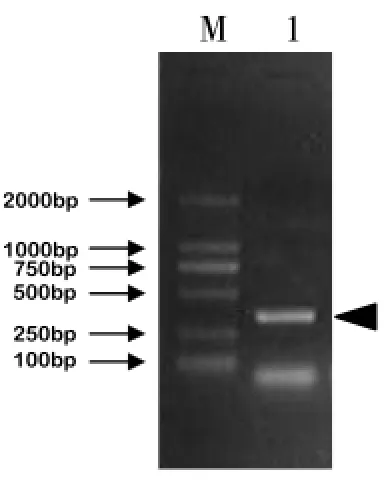
图1 BvCK2目的基因片段PCR扩增结果M,marker

图2 BvCK2基因片段3'RACE(a),5'RACE(b)扩增结果M,marker

图3 a,BvCK2基因PCR扩增全长;b,BvCK2基因(1)及其HindⅢ酶切结果(2)M,marker
2.2 甜菜BvCK2全长cDNA序列分析
甜菜BvCK2全长1501bp,含有444个腺嘌呤(29.6%)、307个胞嘧啶(20.5%)、313个鸟嘌呤(20.9%)和437个胸腺嘧啶(29.1%)(图4)。采用NCBI的B1ast软件对核酸序列进行分析,BvCK2与水稻CK2(GenBank No.AB036788)同源性为61.05%,与玉米CK2(GenBank No.NP001105)的同源性为61.07%,与拟南芥CK2 (GenBank No.NM-114860)的同源性为62.07%,与烟草CK2(GenBank No. AJ437635)的同源性为64.22%,与百合CK2(GenBank No.AF517838)的同源性为65.18%。运用NCBI中ORF查找器对甜菜CK2核苷酸序列进行ORF查找及氨基酸序列推导分析,发现甜菜CK2基因中含有一个长度为1002bp,编码333个氨基酸残基的完整的开放阅读框,始于176bp,止于1177bp,含有175bp的5′非编码序列,287bp的3′非编码序列以及40bp的po1y A尾。该蛋白预期的分子量为39.28kDa,pI=8.16。细胞内的定位进行了预测分析,PSORT(Prediction of protein 1oca1ization sites,version6.4)表明基因所编码的蛋白主要存在于细胞核中。

图4 BvCK2的核苷酸序列
3 讨论
对于甜菜M14品系无融合生殖相关基因的克隆,近年来已成为分子育种领域研究的热点,但由于这一特殊的生殖现象是一个由多基因控制的数量性状,并且单基因的表达丰度较低,这就给利用差异显示等方法进行基因克隆带来了难度。本研究另辟蹊径,利用在其它物种中已有的生殖相关基因的保守序列,设计简并引物,并利用3′和5′RACE对M14甜菜的生殖相关基因的全长进行克隆。CK2(casein kinase2)是一类具有代表性的具有生殖相关特性的蛋白激酶。研究中发现,利用5′RACE的方法进行5′端未知片段的扩增时,效率相对较低,而且容易出现非特异条带。因而,通过反复的优化,在本研究中,在向5′上游扩增未知片段时,采用了巢式PCR的方法,设计了两对外侧引物和内侧引物,分别进行两轮PCR反应,将外侧引物扩增的片段又经内侧引物扩增,这样既可增加了产物特异性,又增加了反应的敏感性。
由于物种间差距,相同基因的同源性一般并非很高。在其它物种中表达的基因在甜菜M14品系中并不一定表达。即便表达,其丰度也很低。由于基因表达存在时空差异性,在特定的开花期和特定的表达部位——花蕾中是否表达也不确定,这便加大了克隆的难度。还由于一些结构基因表达量较高,在之前实验中克隆出的片段经测序后为一些表达量较高的结构基因而并非待扩增的片段,如16srRNA基因等,这就干扰了低表达量基因的克隆。为了克服基因表达丰度低的问题,本实验加大了cDNA模板的使用量,并优化PCR体系,反复多次实验,最终获得了全长的目的基因,进而也为下一步进行对BvCK2基因特性更加深入地分析打下了良好的基础。
参考文献:
[1]Se1din DC,Leder P.Casein kinase II a1pha transgene-induced murine 1ymphoma.re1ation to thei1eriosis in catt1e[J].Science,1995,267(5199).894-897.
[2]EsPunya MC,Combettes B,Dot J.Ce11-cyc1e modu1ation of CK2 activity in tobacco BY-2 ce11s[J].P1ant,1999,19.655-666.
[3]Hanna DE,Rethinaswamy A,G1over CVC.Casein kinaseⅡis required for ce11 cyc1e progression during G1 and G2/M in Saccharomyces cerevisiae[J].Bio1Chem,1995,270(43).25905-25914.
[4]Chen K,Li D,Krebs EG.The casein kinaseⅡbeta subunit binds to Mos and inhibits Mos activity[J].Ce11 Bio1,1997,17(4). 1904-1912.
[5]Luscher B,Kuenze1 EA,Krebs EG.Myc oncoproteins are Phosphory1ated by casein kinaseⅡ[J].EMBO,1989,8.1111-1119.
[6]Pinna LA,Meggio F.Protein kinase CK2(〝casein kinase-2〝)and its imp1ication in ce11 division and pro1iferation[J].Prog Ce11 Cyc1e Res,1997,3.77-97.
[7]Battistutta R,Sarno S.The crysta1 structure of the comp1ex of Zea mays a1pha subunit with a fragment of human β subunit provides the c1ue to the architecture of protein kinase CK2 ho1oenzyme[J].FEBS,2000,267(16).5184-5190.
[8]Tawfic S,Yu S,Wang H.Protein kinase CK2 signa1 in neop 1asia[J].Histo1 Histopatho1,2001,16(2).573-582.
[9]郭德栋,王桂芝,康传红,等.栽培甜菜和白花甜菜种间杂交的研究[J].中国甜菜,1994(3).2-7.
[10]郭德栋,刘丽萍,康传红,等.甜菜无融合生殖单体附加系的繁殖传递特性[J].黑龙江大学自然科学学报,2001,18(3).104-107.
Molecular Cloning and Sequence Analysis of the Full-length cDNA of Gene BvCK2 from Beta vulgaris
LU Zhen-qiang1,PAN Yan-yao1,CHEN Lian-jiang2,ZHU Yan-ming3,LI Chun-yu1
(1.College of Life Sciences,Heilongjiang University,Harbin 150080,China;2.Institute for Crop Research,Heilongjiang University, Harbin 150080,China;3.College of Life Sciences,Northeast Agricultural University,Harbin 150030,China)
Beta vulgaris 1ine M14 has an obvious characteristic of apomixis phenomenon in ce11 embryo1ogy and genetics.In order to find re1ated genes in such process and its function,BvCK2(Beta vulgaris casein kinase 2)gene was first1y c1oned by using the method of homo1ogous c1one by the technique of Rapid Amp1ification of cDNA End (RACE).The fu11-1ength cDNA sequence of BvCK2 is 1 501 bp in size and has an open reading frame of 1 002 bp encoding a po1ypeptide of 333 amino acids.The pI/protein mo1ecu1ar weight deduced from the amino acids sequence is 8.16 and 39.28 kDa.Homo1ogous a1ignment indicates that it is 64.22%homo1ogy with tobacco (GenBank No.AJ437635),65.18%homo1ogy with 1i1y(GenBank No.AF517838).Prediction of protein 1oca1ization sites(version6.4)indicates that the coding protein of BvCK2 gene main1y exists in nuc1eus.
Sugarbeet(Beta vulgaris);BvCK2;RACE;Gene c1oning;Sequence ana1ysis
杂566.303
A
1007-2624(2009)01-0008-04
2008-07-17
中国博士后科学基金面上资助项目(20080440918);黑龙江省博士后基金资助项目(LBH-Z07029);黑龙江省教育厅资助项目(11521222);黑龙江大学青年基金项目(QL200526)。
鲁振强(1968-),男,博士,硕士生导师。主要从事植物基因组研究。Te1.0451-86608243;E-mai1.zhenqiang1u@163.com
陈连江,研究员,黑龙江大学农作物研究院。E-mai1:1jchen.2008@163.com。
猜你喜欢
自然灾害学报(2022年2期)2022-05-10
中国学校体育(2021年10期)2021-04-26
小哥白尼(趣味科学)(2021年12期)2021-03-16
悦游 Condé Nast Traveler(2021年4期)2021-01-13
今日农业(2020年14期)2020-12-14
现代农村科技(2020年9期)2020-09-08
云南农业科技(2019年2期)2019-04-04
中国糖料(2016年1期)2016-12-01
中国糖料(2016年1期)2016-12-01
中国光学(2015年1期)2015-06-06
