
不同盐度波动幅度对缢蛏生长和生化组成的影响
2010-12-28 10:23王兴强马甡曹梅阎斌伦
海洋通报 2010年5期
王兴强,马甡,曹梅,阎斌伦
(1. 淮海工学院 江苏省海洋生物技术重点实验室, 江苏 连云港 222005; 2. 中国海洋大学水产学院海水养殖实验室, 山东 青岛 266003)
不同盐度波动幅度对缢蛏生长和生化组成的影响
王兴强1,马甡2,曹梅1,阎斌伦1
(1. 淮海工学院 江苏省海洋生物技术重点实验室, 江苏 连云港 222005; 2. 中国海洋大学水产学院海水养殖实验室, 山东 青岛 266003)
研究了盐度波动幅度(盐度恒定组S0为对照,盐度波动幅度设3、6、9、12和15五个水平,分别用S3、S6、S9、S12和S15表示)对缢蛏生长和生化组成的影响,实验周期90 d。结果表明:随盐度波动幅度的升高,缢蛏存活率、分别以末壳长度和末体质量表示的特定生长率(SGRFL和SGRFQ)呈上升趋势,当到达临界值后,其存活率、SGRFL和SGRFQ随盐度波动幅度的升高而下降;而缢蛏水分和粗蛋白含量呈下降趋势,当到达临界值后,其水分和粗蛋白含量随盐度波动幅度的升高而上升。Turkey’s多重比较发现,S3、S6和S9盐度波动处理组缢蛏的存活率、SGRFL和粗脂肪含量与对照组相比组间差异不显著(P>0.05),而S6和S9盐度波动处理组缢蛏的SGRFQ显著高于对照和其它处理组(P<0.05)。缢蛏存活率、末壳长度、末体质量、SGRFL、SGRFQ、水分含量、粗蛋白含量、粗脂肪含量和灰分含量达到最适值的盐度波动幅度分别为3.24、5.16、5.47、5.00、5.41、6.20、5.32、5.20和6.94。
缢蛏;盐度波动幅度;生长;生化组成
缢蛏(Sinvnovacula constricta(Lamarck))属双壳纲(Bivalvia)、帘蛤目(Veneroida)、灯塔蛤科(Pharellidae)、缢蛏属(Sinvnovacula),俗称蛏、蜻、海蛏或跣等,自然分布于河口或有少量淡水注入的内湾潮间带中、下区软泥滩涂洞穴内[1]。缢蛏营滤食性埋栖生活,可有效改善水质、净化养殖水环境,目前已成为中国沿海池塘综合养殖的主要混养贝类之一[2]。
盐度是海水养殖中重要的环境因子,在养殖生产中由于换水、暴雨和蒸发等原因,在河口附近及集水面积较大的沿海滩涂养殖区,水体盐度常处于波动状态[3-5]。目前,关于不同盐度波动幅度对鱼类和虾类生长和代谢的影响已有报道;一些研究表明,在一定限度内盐度的适宜波动会对水生生物的生长产生积极的促进作用[6-8]。关于盐度变化与贝类生态学响应的研究主要集中在短时间内盐度变化前后贝类的生态学特点,但缺乏盐度节律性连续变化下贝类的生态学反应的报道[9]。缢蛏运动能力相对较弱,因而所受到的盐度变化可能比鱼类、虾类等其它海洋动物所面临的更为剧烈,其对盐度变化的响应规律和适应机制可能也不同。鉴于此,本文初步研究了盐度波动幅度对缢蛏生长和生化组成的影响。
1 材料和方法
1.1 实验配合饲料
实验所用配合饲料配方见表 1,根据配方比例,首先对原料进行预处理,然后配料、混合、粉碎(粒度直径<30 µm)和50 ℃烘干,4 ℃冷藏保存备用。

表 1 饲料配方Tab. 1 Ingredient and proximate composition of the experimental diets
1.2 材料来源
实验缢蛏(壳长度3.2~3.5 cm,体质量1.4~1.9 g)取自连云港市燕尾港缢蛏土池养殖池,缢蛏在实验室玻璃纤维水槽(200×200×100 cm)中暂养1 wk,水温23.5~25.0 ℃,盐度30,暂养期间投喂实验配合饲料(表1)。
1.3 实验设计和管理
实验采用单因子 6水平设计,盐度恒定组S0为对照,盐度波动幅度设3、6、9、12和 15五个水平,分别用S3、S6、S9、S12和S15表示。实验起始盐度均为30,第一次换水即降低盐度,突降幅度分别为3、6、9、12和15,第二次换水时又将盐度升至盐度30,第三次换水与第一次相同,第四次换水与第二次相同,依次类推,S0的盐度始终控制在30。每个处理设5个重复,共30个水族箱。
模拟缢蛏自然生态环境,从自然海区滩涂挖掘泥沙(取自海区滩涂表面10 cm以上部分),经太阳曝晒、粉碎后均匀铺于玻璃水族箱(30×40×50 cm,养殖水体约40 L)中,泥沙厚度大约为10 cm,然后加入砂滤海水大约至35 cm,搅拌均匀,平衡24 h后,再次更换清洁海水,放入健康无损、钻洞能力强的大小相近的实验缢蛏,每个水族箱放 10只,实验前1 d停止投喂,缢蛏初壳长度和初体质量分别见图1和图2。预实验表明,室内养殖缢蛏混养龙须菜(Gracilaria lemaneiformis)有助于缢蛏的存活和生长;因此,每个水族箱2/3水面混养约3 cm厚的龙须菜。每天上午06:00 h投喂实验配合饲料(表 1),下午 18:00 h投喂小新月菱形藻液(Nitzschia. closterium f. minutissima),均过量投饵;投饵2.5 h后用虹吸管从每个水族箱轻轻吸走残饵和排泄物。实验用水由过滤海水加自来水配制,每5 d彻底换水一次,并补充龙须菜和曝晒粉碎的自然海区滩涂泥沙到初始厚度。
实验期间,溶解氧维持在 6.0 mg/L以上,pH 7.7~8.2,光周期14L:10D,水温25.0±0.5 ℃。实验周期为90 d。实验结束时,同样饥饿l d,测量缢蛏末壳长度和称量缢蛏末体质量(图1和图2),然后用解剖刀取出缢蛏软体部,60 ℃烘干至恒重。粗脂肪的测定采用索氏抽提法。样品中粗蛋白的含量首先利用 VarioELⅢ型元素分析仪(Elementar,German)测定氮含量,然后乘 6.25获得。在马弗炉中550 ℃燃烧12 h,获得样品灰分含量。本实验所得粗蛋白、粗脂肪和灰分均以干重计。
1.4 数据处理与计算
特定生长率的计算公式如下:

式中的IL和FL为缢蛏的初壳长度和末壳长度,IQ和FQ为缢蛏的初体质量和末体质量,t为实验周期。
所得数据用单因素方差分析、Turkey’s多重比较以及回归分析进行处理,以P<0.05作为差异显著水平。
2 结 果
2.1 不同盐度波动幅度下缢蛏的生长
结果和变量分析见图 1~图 4。随盐度波动幅度的升高,缢蛏存活率、末壳长度、末体质量、分别以末壳长度和末体质量表示的特定生长率(SGRFL和SGRFQ)呈上升趋势,当到达临界值后,其存活率、末壳长度、末体质量、SGRFL和SGRFQ随盐度波动幅度的升高而下降。其中,S3和S6盐度波动处理组缢蛏的存活率高于对照,而S9、S12和S15盐度波动处理组缢蛏的存活率低于对照;S3、S6和S9盐度波动处理组缢蛏的末壳长度、末体质量、SGRFL和SGRFQ均高于对照,而S12和S15盐度波动处理组缢蛏的末壳长度、末体质量、SGRFL和SGRFQ均低于对照。

图 1 盐度波动幅度(S)对缢蛏末壳长度的影响Fig. 1 Effect of salinity fluctuation on the final shell length of S. constricta
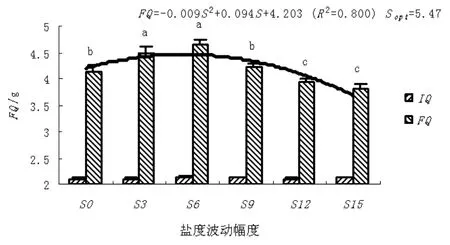
图 2 盐度波动幅度(S)对缢蛏末体质量的影响Fig. 2 Effect of salinity fluctuation on the final quality of S. constricta
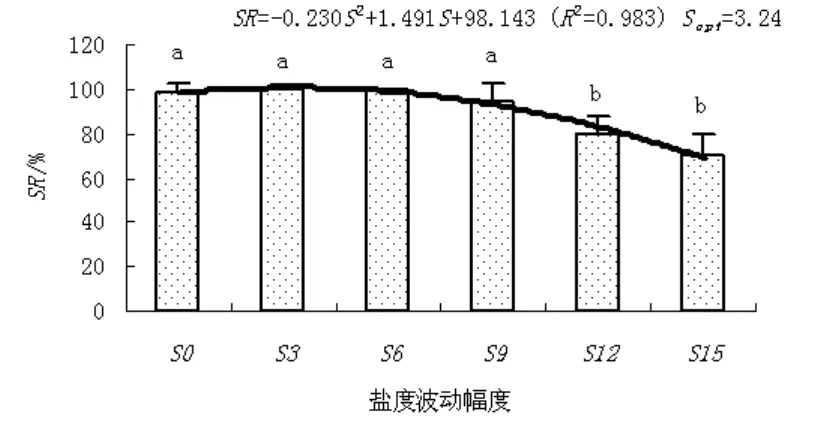
图 3 盐度波动幅度(S)对缢蛏存活率(SR)的影响Fig. 3 Effect of salinity fluctuation on the survival rate of S. constri cta
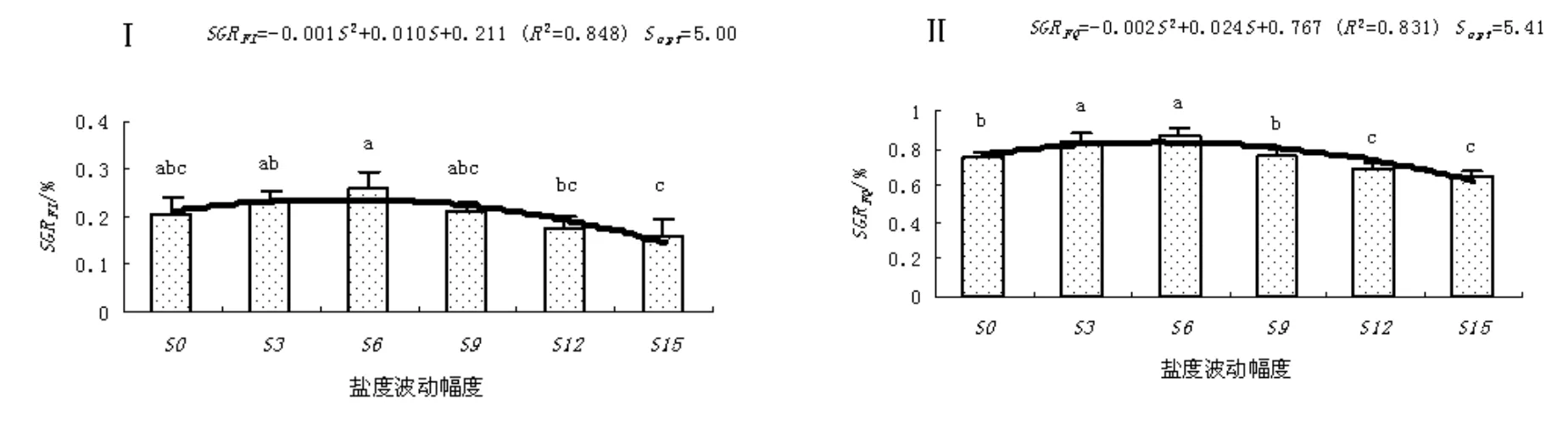
图 4 盐度波动幅度(S)对缢蛏特定生长率的影响Fig. 4 Effect of salinity fluctuation on the specific growth rate of S. constricta
单因素方差分析表明,盐度显著的影响缢蛏的存活率、末壳长度、末体质量、SGRFL和SGRFQ(P<0.05)。Turkey’s多重比较发现,S3、S6和S9盐度波动处理组缢蛏的存活率和SGRFL与对照组相比组间差异不显著(P>0.05),而S6和S9盐度波动处理组缢蛏的SGRFQ显著高于对照和其它处理组(P<0.05)。回归分析表明,缢蛏存活率、末壳长度、末体质量、SGRFL和SGRFQ达到最适值的盐度波动幅度分别为3.24、5.16、5.47、5.00和5.41。
2.2 不同盐度波动幅度下缢蛏的生化组成
结果和变量分析见图5。随盐度波动幅度的的升高,缢蛏水分和粗蛋白含量呈下降趋势,当到达临界值后,其水分和粗蛋白含量随盐度波动幅度的升高而上升。在S0和S3盐度波动幅度下,缢蛏脂肪含量相等;随盐度波动幅度的的升高,缢蛏脂肪含量呈上升趋势,当到达临界值后,其脂肪含量随盐度波动幅度的升高而下降。其中,S3、S6和S9盐度波动处理组缢蛏的水分、粗蛋白和灰分含量均低于对照组,而粗脂肪含量均高于对照组。
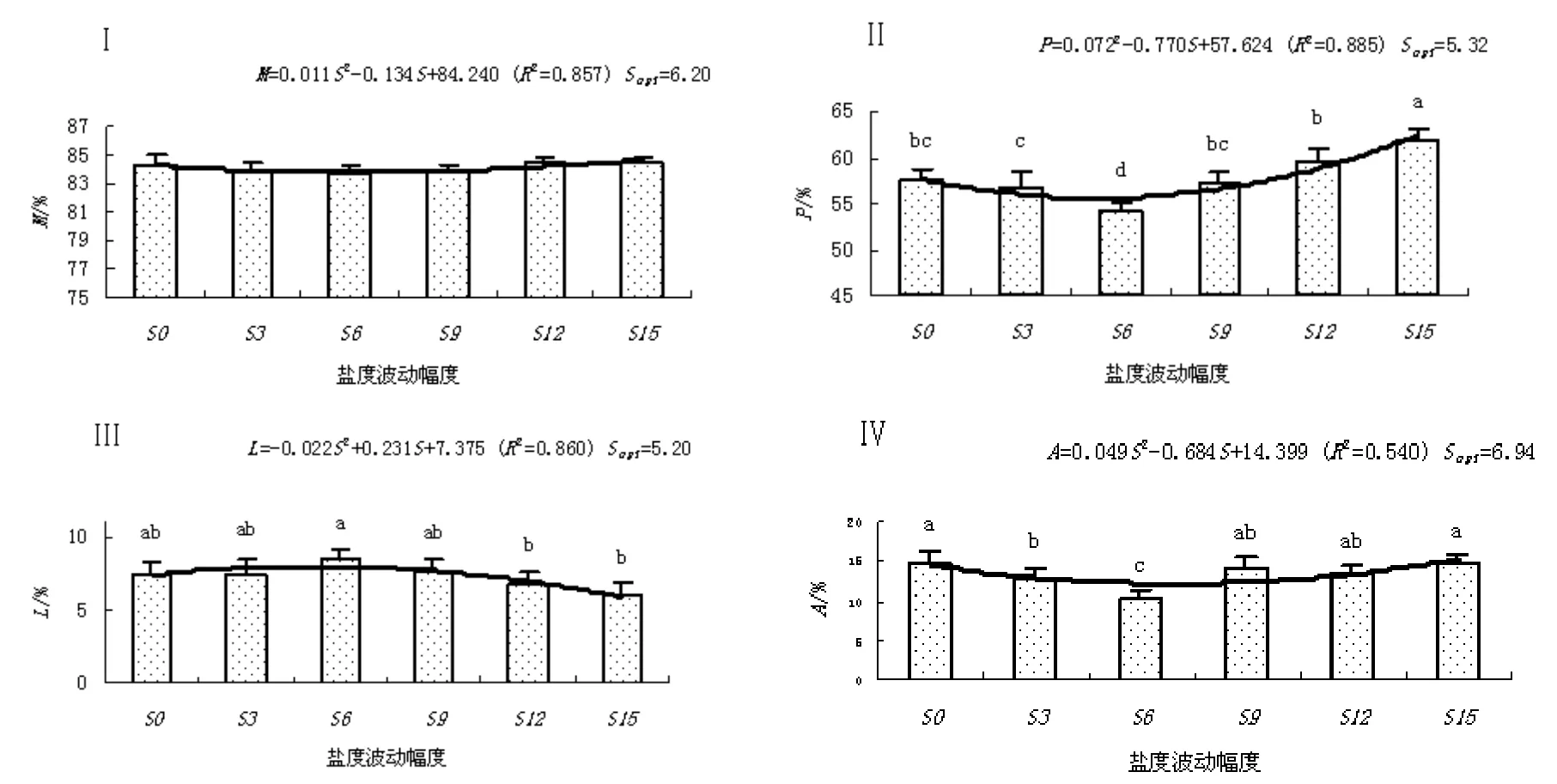
图 5 盐度波动幅度(S)对缢蛏水分含量和生化组成(干质量)的影响Fig. 5 Effect of salinity fluctuation on the moisture content and biochemical composition of S. constricta
单因素方差分析表明,盐度波动幅度显著的影响缢蛏的粗蛋白、粗脂肪和灰分含量(P<0.05)。Turkey’s多重比较发现,S3和S9盐度波动处理组缢蛏的粗蛋白、粗脂肪和灰分含量与对照组相比组间差异不显著(P>0.05),而S3和S6盐度波动处理组缢蛏的粗蛋白和灰分含量显著低于对照(P<0.05)。回归分析表明,缢蛏水分、粗蛋白、粗脂肪和灰分含量达到最适值的盐度波动幅度分别为6.20、5.32、5.20和6.94。
3 讨 论
3.1 不同盐度波动幅度下缢蛏的生长
盐度是影响水生动物生理状态的主要环境因子之一,包括生长、新陈代谢和营养需求等[11-13]。目前,人们对某些恒定盐度下缢蛏生长、代谢和繁殖的状况有所了解,但关于盐度周期性变化对缢蛏的影响研究较少[14-18]。缢蛏主要生活在盐度变化较大的内湾或河口附近,在此环境中,理化因子变化较大,特别是盐度变化更为明显,长期的生存适应已使得缢蛏能长时间耐受较大的盐度波动。从图3可以看出,在0~9盐度波动幅度范围内,缢蛏存活率组差异不显著(P>0.05)。穆迎春等[6]研究发现,在一定的波动幅度内改变盐度可促进中国明对虾(Fenneropenaeus chinensis)进行渗透调节,降低对虾用于代谢上的能量,进而加快对虾的生长。在鱼类的研究中,戴桂珍[8]提出生长加快是鱼类在盐度波动影响下发生的一种普遍性反应,鱼类在盐度以一定幅度波动的情况下所表现出的生长潜力是无论在任何恒定盐度中都发挥不出来的。从图4可以看出,S6和S9盐度波动处理组缢蛏的SGRFQ显著高于对照和其它处理组(P<0.05),这表明,在周期性的改变盐度条件下,适宜的盐度波动幅度可促使缢蛏的生长。同样,在自然条件下,由于降雨和地表径流的影响,近岸潮间带的生物都要经历有节律的短期或长期的盐度变化;尤其生活在潮间带的缢蛏,由于不能移动,每天都要忍受因周期性潮汐作用和水分蒸发所带来的盐度节律性的连续变化。由此可以推测,水生动物的最适盐度是一个盐度区间而不是一个点,而水生动物对盐度波动的反应与适应于环境渗透压的改变有关;当水生动物生活的盐度周期性地偏离其恒定状况下的最适值时,环境渗透压的波动在其承受能力范围之内,并伴随着生理状况和能量代谢的改善,从而生长得到加快、存活率得到提高。
不过,如果盐度波动幅度过大,则不利于缢蛏的存活和生长。从图1至图4可以看出,在S15盐度波动下,缢蛏存活率、末壳长度、末体质量、SGRFL和SGRFQ均最低;Turkey’s多重比较发现,盐度波动幅度超过9时,缢蛏存活率和SGRFQ显著下降(P<0.05)。在生产实践中,已有多起因暴雨天气、径流过大和山洪暴发等使海水盐度骤降而造成缢蛏大规模死亡事件;而且死亡呈现离河口越近,死亡率越高的特点[11]。林笔水等[18]曾报道,大幅度的盐度突变情况下,缢蛏存活率显著降低;即使能存活,在养成中也易感染疾病,并在一定程度上影响生长速度。
3.2 不同盐度波动幅度下缢蛏的生化组成
贝类品质优劣涵盖营养价值、风味和矿物元素等多个方面;其中,水分、蛋白质、脂肪和灰分含量等是鉴定贝类营养价值的重要指标。贝类的生化组成与其生理机能密切相关,而生理机能又与其生活环境密切相关[13]。目前,有关盐度波动对贝类生化组成影响的研究报道甚少。对广盐性的缢蛏而言,在适宜的盐度范围内,缢蛏体内外渗透压平衡,所以只需花较少的能量来维持体内环境的稳定,而盐度超过其适宜范围后,缢蛏则需消耗较多的能量来适应外界环境的盐度变化,导致机体能量物质被过多消耗[19]。本研究中,S3和S9盐度波动处理组缢蛏的粗蛋白、粗脂肪和灰分含量与对照组相比组间差异不显著(P>0.05),而S3和S6盐度波动处理组缢蛏的粗蛋白含量显著低于对照,S15盐度波动处理组缢蛏的粗蛋白含量显著高于对照(P<0.05)。其中原因,可能与盐度波动影响缢蛏渗透调节和代谢耗能,进而影响蛋白质的沉积有关;从本研究可以推测,在不利的盐度条件下,缢蛏首先消耗体蛋白作为能量的补充。
[1] 王如才, 王昭萍, 张建中. 海水贝类养殖学 [M]. 北京: 青岛海洋大学出版社, 1993: 247-252.
[2] 田相利, 李德尚, 董双林, 等. 对虾-罗非鱼-缢蛏封闭式综合养殖的水质研究 [J]. 应用生态学报, 2001, 12(2): 287-292.
[3] PARADO-ESTEPA F D, FERRARIS R P, LADJA J M, et al.Responses of intermolt Penaeus indicus to large fluctuations in environmental salinity [J]. Aquaculture, 1987, 64(3): 175-184.
[4] GAMMELSRØD T. Variation in shrimp abundance on the Sofala Bank, Mozambique, and its relation to the Zambezi River runoff [J].Estuarine, Coastal and Shelf Science, 1992, 35(1): 91-103.
[5] 韦蔓新, 童万平, 何本茂. 台风对工厂化虾池生态系统中 COD的影响 [J]. 湛江海洋大学学报, 2005, 6: 46-50.
[6] 穆迎春, 王芳, 董双林, 等. 不同盐度波动幅度对中国明对虾稚虾蜕皮和生长的影响 [J]. 海洋学报, 2005, 27(2): 122-126.
[7] 丁森, 王芳, 郭彪, 等. 盐度波动对中国对虾稚虾蜕皮、生长和能量收支的影响 [J]. 应用生态学报, 2008, 19(2): 419-423.
[8] 戴桂珍. 盐度波动对幼鱼生长的影响 [J]. 水利渔业, 1993, 4:53-55.
[9] 王吉桥, 靳翠丽, 姜静颖. 不同降盐度方式对海湾扇贝幼贝存活的影响 [J]. 中国水产科学, 2001, 8(3): 35-39.
[10] MAI K, MERCER J P, DONLON J. Comparative studies on the nutrition of two species of abalone, Haliotis tuberculata L and Haliotis discus hannai Ino IV. Optimum dietary protein level for growth [J]. Aquaculture, 1995, 136: 165-180.
[11] 周海嘉, 杨运生, 赵连成. 盐度骤降后缢蛏死亡原因浅析和对策[J]. 苏盐科技, 2004, 3: 18-19.
[12] PARADO-ESTEPA F D, FERRARIS R P, LADJA J M, et al.Responses of intermolt Penaeus indicus to large fluctuations in environmental salinity [J]. Aquaculture, 1987, 64(3): 175-184.
[13] SARAÀ G, ROMANO C, WIDDOWS J, et al. Effect of salinity and temperature on feeding physiology and scope for growth of an invasive species (Brachidontes pharaonis- MOLLUSCA:BIVALVIA) within the Mediterranean sea [J]. Journal of Experimental Marine Biology and Ecology, 2008, 363(1-2):130-136.
[14] 刘泉顺. 几个环境因子对缢蛏土池人工育苗的影响 [J]. 海洋通报, 1984, 3(5): 41-44.
[15] 林笔水, 吴天明. 温度和盐度同缢蛏稚贝存活及生长的关系 [J].水产学报, 1986, 10(1): 41-50.
[16] 吴天明, 林笔水. 几种环境因子对缢蛏苗存活率的影响 [J]. 台湾海峡, 1987, 6(2): 120-126.
[17] 王文雄, 许振祖. 缢蛏稚贝的爬行行为及盐度对其钻穴能力的影响 [J]. 厦门大学学报, 1988, 27(3): 333-337.
[18] 林笔水, 吴天明. 温度与盐度和缢蛏幼体生存、生长及发育的关系 [J]. 水产学报, 1990, 14(3): 171-178.
[19] 沈伟良, 尤仲杰, 施祥元. 不同规格及不同盐度下毛蚶稚贝耗氧率和排氨率的研究 [J]. 海洋水产研究, 2008, 29(2): 53-56.
Effect of salinity fluctuation on the growth and biochemical composition ofSinvnovacula constricta
WANG Xing-qiang1, MA Shen2, CAO Mei1, YAN Bin-lun
(1. Jiangsu Key Laboratory of Marine Biotechnology, Huaihai Institute of Technology, Lianyungang 222005, China;
2. Mariculture Research Laboratory, Fisheries College, Ocean University of China, Qingdao 266003, China)
The study was conducted to determine the effect of salinity fluctuation (S0, the control group exposed to the constant salinities,S3,S6,S9,S12andS15as the representative of salinity fluctuations of 3‰, 6‰, 9‰, 12‰ and 15‰, respectively) on the growth and biochemical composition ofSinvnovacula constricta. The experiment lasted for 90 days. The results showed that the survival and specific growth rate (the finial shell lengthSGRFLand final qualitySGRFQ) ofS. constrictaexhibited increased trends, reaching the maximum value, and thereafter declined, whereas the content of moisture and crude protein exhibited decrease trends, reaching the minimum value, and thereafter rise.Tukey’s test indicated that the survival,SGRFLand the crude lipid content ofS. constrictaatS3,S6andS9were not significantly different as compared with that of the control (P>0.05), whereasSGRFQatS6andS9were significantly higher than that of the control and other treatments (P<0.05). Regression analysis indicated that the optimum salinity fluctuation of the survival, finial shell length, final quality,SGRFL,SGRFQ, moisture content, crude protein content,crude lipid content and ash content were 3.24, 5.16, 5.47, 5.00, 5.41, 6.20, 5.32, 5.20 and 6.94, respectively.
Sinvnovacula constricta; salinity fluctuation; growth; biochemical composition
X174; Q958.11
A
1001-6932(2010)05-0567-05
2009-07-05;
2010-01-16
江苏省自然科学基金项目(BK2006548)
王兴强 ( 1975- ),男,博士,主要从事水生动物生理生态学研究。电子邮箱:xqwangcaomei@yahoo.com.cn
猜你喜欢
股市动态分析(2021年25期)2021-12-30
当代水产(2021年4期)2021-07-20
当代水产(2020年4期)2020-06-16
宇航计测技术(2018年3期)2018-09-08
制造业自动化(2017年2期)2017-03-20
广东海洋大学学报(2015年3期)2015-12-22
华南农业大学学报(2015年5期)2015-12-04
应用海洋学学报(2015年3期)2015-11-22
中国洗涤用品工业(2015年5期)2015-02-28
汉语世界(2015年5期)2015-01-05
