
日粮精氨酸水平对生长肉兔氮代谢及肝脏胰岛素样生长因子(IGF-I)基因表达的影响
2011-08-09 11:14
饲料工业 2011年16期
精氨酸作为幼年家畜和家禽的必需氨基酸,具有重要的营养、代谢和免疫功能。近年来,精氨酸已经成为许多科学家的研究热点。越来越多的研究已经表明,肝脏胰岛素样生长因子(IGF-I)在动物生长发育中起着重要的调节作用。IGF既是GH发挥作用的最主要介导因子,又具有与GH无关的独特作用。Wray-Cahen C D[1]研究表明,IGF是动物生长的直接调节物,可以刺激多种细胞的增殖与分化,促进蛋白质的合成和结缔组织及骨髓的产生。了解IGF的作用及其变化规律对于了解动物的生长及某些疾病的发生机理,运用各种(遗传和营养)手段实现对动物生长的调控,从而提高动物的健康和生产水平与效率具有重要的理论和实践意义[2]。
本试验以断奶~2月龄生长肉兔为研究对象,在其他各种营养素满足需要的情况下,探讨添加不同精氨酸水平对氮代谢和肝脏细胞中IGF-I基因表达水平的影响。
1 材料与方法
1.1 试验动物及饲养管理
体重基本一致的100只断奶肉兔。试兔单笼饲养,试验期间日喂食2次,采用常规饲养管理和免疫程序,自然采光、通风。自由采食,自由饮水。
1.2 试验日粮及试验设计
基础日粮(对照组)参照NRC(1977)家兔饲养标准配制而成。试验日粮4组,在基础日粮中以DL-精氨酸盐酸盐的形式添加精氨酸,添加量分别为0.2%、0.4%、0.6%、0.8%,日粮加工成直径约4~6 mm的颗粒饲料,储存在通风干燥避光处备用。基础日粮组成及营养水平见表1。
100只试兔按照体重分为5组,每组20只,公母各半,各组间体重差异不显著。对照组饲喂基础日粮,试验组分别饲喂以上4种日粮,预试期7 d,正式期23 d。
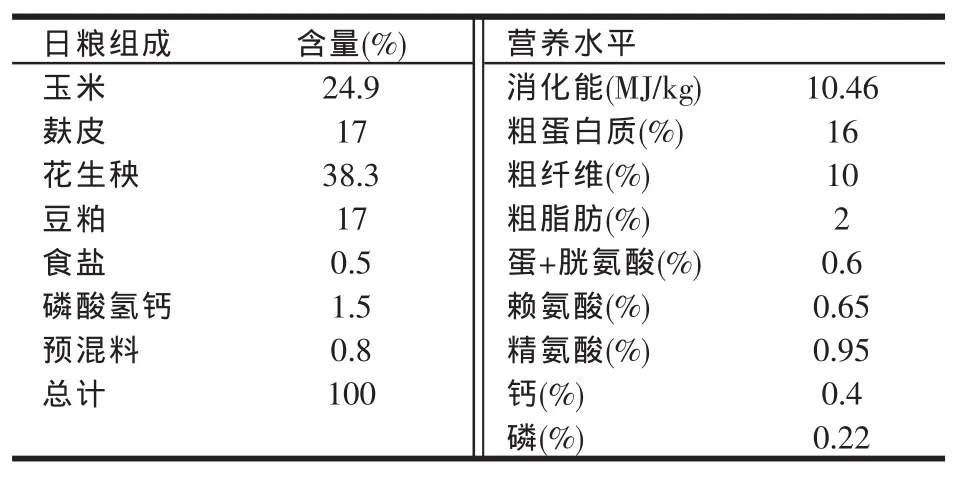
表1 基础日粮组成及营养水平
1.3 氮代谢指标的测定
饲养试验阶段结束前6 d,每组随机抽取6只试验兔转移到经消毒处理的代谢笼内,单笼饲养,并饲喂相应的试验日粮,自由采食和饮水。预试期3 d,后3 d分别收集每只兔全天的粪样和尿样(每天上午8~9点,下午4~5点),4℃密封保存,连收3 d,同时记录每只试验兔每天的采食量、排粪量和排尿量。鲜粪称重后,加10%的硫酸固氮,然后在烘箱中于 65℃下烘干 24 h,称重即粪样的风干重,最后将3 d的风干粪样混合粉碎待测。尿样量取后按一定的比例倒入已准备好的250 ml塑料瓶中,加入5 ml浓硫酸固氮,每天取样比例相同,混匀后冷冻保存备用。样品含氮量的测定依据《饲料分析及饲料检测技术》(张丽英,2002)进行测定。
可消化氮(DN)=食入氮(IN)-粪氮(FN);
沉积氮(RN)=食入氮(IN)-粪氮(FN)-尿氮(UN);
氮表观消化率(DN/IN,%)=DN/IN×100;
食入氮转化为沉积氮的效率(RN/IN,%)=RN/IN×100;
可消化氮利用率(RN/DN,%)=RN/DN×100。
1.4 肝脏样品的采集与制备
在试验开始后的第30 d早晨空腹称重,每组随机抽取6只试验兔,以颈椎错位法将其致死,屠宰,立刻取肝脏相对固定部位样品,放入Eppendorf管中,每管约0.5 g,每组织6管重复,液氮速冻后,-70℃保存备用。
1.5 实验方法
对家兔肝脏细胞中IGF-I基因的表达水平进行了相对定量分析,采用SYBR GreenⅠ荧光染料嵌合法,以家兔肝脏样品提取的Total RNA转录的cDNA作为标准品,分别对管家基因(GAPDH)和目的基因(IGF-I)制作标准曲线。利用标准曲线对各样品管家基因和目的基因分别进行定量,通过管家基因的校正,检测各样品目的基因的相对表达量。
1.5.1 实验试剂
TaKaRa SYBR PrimeScriptTM、RT-PCR Kit(Perfect Real Time)(TakaRa Code.DRR063A);TaKaRa DNaseⅠ (Rnase Free)(TaKaRa Code.D2215);TaKaRa RNAiso Reagent(TaKaRa Code.D312);Easy Dilution(for Real Time PCR)TaKaRa Code.D9160;DNA Marker DL 2000(TaKaRa Code.D501)。
1.5.2 主要实验仪器
TaKaRa PCR Thermal Cycler Dice(TakaRa Code.TP600);TaKaRa PCR Thermal Cycler Dice、Real Time System(TaKaRa Code.TP800)。
1.6 实验步骤
1.6.1 RNA的提取
1.6.1.1 前处理
凡与RNA操作有关的玻璃器皿和剪刀、镊子等器械经常规洗涤烘干后,用锡箔纸包好,180℃烘烤8 h或200℃烘烤2~4 h。塑料制品用0.1%DEPC浸泡处理37℃2 h,或室温处理过夜,再用灭菌双蒸水漂洗数次,高压消毒去除DEPC。(注:操作过程中用到的所有试剂必须用DEPC处理过的水配制)。
1.6.1.2 组织中总RNA的提取(Trizol法)
①取50~100mg组织研磨成粉末(在液氮中操作),转移至1.5 ml离心管中;
②加入1 ml Trizol,用力混匀至清澈,放置在冰上;
③4℃、12 000×g离心10 min,将上清液转移至另一个1.5 ml离心管中;
④加入0.2 ml氯仿,用力摇动15 s,室温放置2~3 min;
⑤4℃、12 000×g离心15 min,将上清液转移至另一个1.5 ml离心管中;
⑥加入0.5 ml异丙醇,混匀,室温放置30 min沉淀RNA;
⑦4 ℃、12 000×g离心 10 min,弃上清;
⑧加入1 ml 75%乙醇(DEPC水配制)洗涤RNA沉淀一次;
⑨4 ℃、7 500×g离心5 min,弃乙醇;
⑩真空干燥,加入 50~70 μl DEPC·H2O 溶解RNA;
⑪分装,-70 ℃保存;
⑫RNA含量的测定 (比色法):将提取的总RNA在紫外分光光度计260 nm和280 nm下测吸光度A值,并计算OD260/OD280的值。此值在1.7~2.0之间说明提取的RNA纯度较高,适宜进行下一步的实验。
1.6.2 引物序列(见表2)

表2 引物序列信息
1.6.3 反应组成及反应条件
1.6.3.1 RT 反应(见表3)

表3 RT反应
1.6.3.2 PCR 反应(见表4)

表4 PCR反应
1.6.4 基因标准曲线的制作
取6个样品的Total RNA(500 ng)反转录获得的cDNA作为标准品,用EASY Dilution按8倍梯度稀释(80、81、82、83、84倍)作为模板,分别制作管家基因和目的基因的标准曲线。管家基因GAPDH标准曲线见图1~图3;目的基因 IGF-I标准曲线见图4~图 6。
1.7 数据处理
用SAS 8.0统计软件中的GLM进行数据的方差分析,用Duncan's法进行数据的多重比较。
2 结果与分析

图1 标准品GAPDH基因标准曲线

图2 标准品GAPDH基因扩增曲线

图3 标准品GAPDH基因熔解曲线

图4 标准品IGF-I基因标准曲线

图5 标准品IGF-I基因扩增曲线

图6 标准品IGF-I基因熔解曲线
2.1 日粮不同精氨酸水平对断奶~2月龄肉兔氮代谢指标的影响(见表5)
由表5可知,日粮中不同水平精氨酸对生长肉兔的尿氮水平影响显著(P值为0.016 7),各添加组的尿氮含量显著低于不添加组(P<0.05)。日粮中不同水平精氨酸对生长肉兔的其它氮代谢指标均无显著影响(P>0.05)。
2.2 产物特异性的确定(见图7、图8)
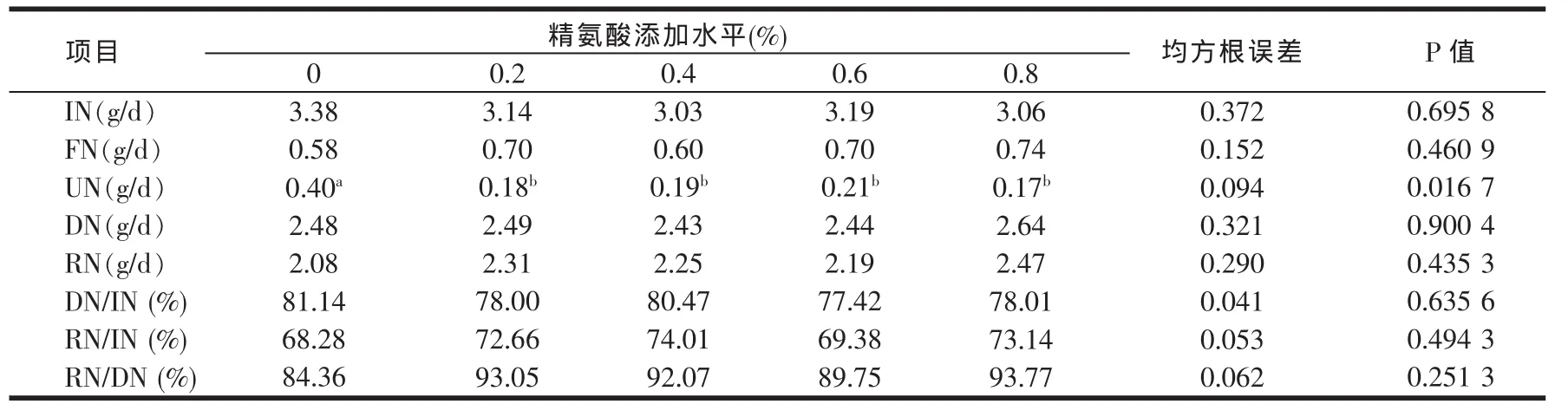
表5 日粮添加精氨酸对断奶~2月龄生长肉兔氮代谢指标的影响

图7 样品IGF-I基因熔解曲线

图8 样品GAPDH基因熔解曲线
由图7和图8样品GAPDH和IGF-I的基因熔解曲线图可以看出,目的基因和管家基因扩增均有单一稳定的峰,说明目的基因和管家基因引物特异性很好,得到了特异性产物。
2.3 扩增曲线(见图9~图10)

图9 样品IGF-I基因扩增曲线

图10 样品GAPDH基因扩增曲线
2.4 精氨酸对断奶~2月龄生长肉兔肝脏IGF-I mRNA表达量的影响(见表6)
由表6可知,日粮不同精氨酸水平显著影响肝脏IGF-I mRNA 表达量(P值为 0.018 2),并且添加精氨酸组IGF-I mRNA表达量明显高于不添加组(P<0.05),各添加组之间差异不显著(P>0.05)。

表6 精氨酸添加水平对断奶~2月龄生长肉兔肝脏IGF-I mRNA表达量的影响
3 讨论与结论
3.1 日粮中添加不同精氨酸水平对断奶~2月龄肉兔氮代谢的影响
肉兔食入饲料的含氮营养物质经体内消化代谢,一部分合成体蛋白沉积体内或被机体所利用,另一部分形成代谢废弃产物随粪、尿排出,构成了家兔的氮平衡。粪N和尿N是食入N的两个损失部分,粪N是经过消化道而没有吸收的部分,这部分N受饲料蛋白含量的影响较大(Spreadkury,1974),尿N是被吸收的氨基酸参加组织代谢,没有被利用合成体蛋白而脱氨后由尿排出,这部分N受饲料氨基酸平衡的影响较大(Ahmed 等,1984)。
本试验中各添加组的尿氮含量显著低于不添加组,说明添加精氨酸能促进尿氮的吸收。但本试验添加精氨酸并未改变氮的沉积和表观消化率等其它氮代谢指标。
3.2 精氨酸对断奶~2月龄生长肉兔肝脏IGF-I mRNA表达量的影响
肝脏是Arg代谢的主要部位,也是循环中IGF-I的主要来源。灌注的肝脏、培养的肝移植物和原代培养肝细胞均可释放IGF-I,肝脏IGF-I mRNA的水平是其他组织的50到100倍。
因食物剥夺或限制引起营养不良时,会抑制肝脏IGF-I的基因表达 (Straus 等,1990;Pell 等,1992;Weller等,1994)[3-5],造成循环 IGF-I水平降低。当蛋白质的数量和质量提高时,小鼠肝脏IGF-I mRNA的含量增加(Miura等,1992)[6]。采食无蛋白日粮或低蛋白日粮的小鼠体重减轻,肝脏IGF-I mRNA表达和循环IGF-I水平降低(Straus等,1990;Vandehaar等,1991)[3,7],同样,低蛋白日粮降低IGF-I mRNA表达和血清IGF-I水平的现象也出现在大动物如绵羊 (Pell等,1992)[4]中。
本试验研究表明,日粮添加精氨酸能提高断奶~2月龄生长肉兔肝脏IGF-I mRNA的表达量,说明精氨酸可能通过生长轴来提高IGF-I mRNA的水平,从而达到促进动物生长的目的。
[1]Wray-Cahen C D,Kerry D E,Evock Clover C M,et al.Redefining by composition:nutrients,hormones,and geese in meat production[J].Ann.Rev.Nutr.,1998,18:63-92.
[2]张克英,陈代文.类胰岛素样生长因子及其营养调控[J].中国畜牧杂志,2002,38(6):42-44.
[3]Straus D S,Takemoto C D.Effect of dietary protein deprivation on insulin-like growth factor(IGF)-I and -II,IGF binding protein-2,and serum albumin gene expression in rat[J].J.Endocrinol.,1990,127:1849-1860.
[4]Pell J M,Bates P C.Differential actions of growth hormone and insulin- like growth factor-I on tissue protein metabolism in dwarf mice[J].J.Endocrinol.,1992,130:1942-1950.
[5]Weller P A,Dauncey M J,Bates P C,et al.Regulation of porcine insulin-like growth factor I and growth hormone receptor mRNA expression by energy status[J].Am.J.Physiol.,1994,266(5):776-785.
[6]Miura Y,Kato H,Noguchi T.Effect of dietary proteins on insulinlike growth factor-I(IGF-I)messenger ribonucleic acid content in rat liver[J].Br.J.Nutr.,1992,67:257-265.
[7]VandeHaar M J,Moats Staats B M,Davenport M L,et al.Reduced serum concentrations of insulin-like growth factor-I(IGF-I)in protein-restricted growing rats are accompanied by reduced IGFImRNA levels in liver and skeletal muscle[J].J.Endocrinol.,1991,130:305-312.
猜你喜欢
今日农业(2022年2期)2022-11-16
现代畜牧科技(2021年2期)2021-03-19
河南畜牧兽医(2020年11期)2020-01-11
科学生活(2019年7期)2020-01-01
养殖与饲料(2019年10期)2019-02-25
山东畜牧兽医(2018年3期)2018-04-26
广东饲料(2016年4期)2016-12-01
中外医疗(2015年11期)2016-01-04
听力学及言语疾病杂志(2015年5期)2015-12-24
中国当代医药(2015年30期)2015-03-01
