
串珠素在咀嚼肌中的表达及意义
2012-07-31 09:22王彦亮黄志锋周稚辉麻健丰
山东医药 2012年35期
朱 莉,王彦亮,黄志锋,何 帅,周稚辉,麻健丰
(1温州医学院附属口腔医院,浙江温州 325027;2温州医学院药学院;3解放军第118医院)
细胞外机制(ECM)是由纤维性蛋白质、氨基多糖和蛋白聚糖等生物大分子组成的动态网络结构,在维持生物结构的完整性、细胞信号转导和细胞表型、功能的调节中均具有重要作用。串珠素是ECM中主要的蛋白聚糖成分,广泛存在多种组织基底膜和软骨基质中,与其他基质成分紧密结合并自行组成二聚体和其他的大聚合物。大量体外研究发现,串珠素在细胞生长和分化、组织化等方面发挥多种功能,可以通过调节生长因子的结合和活性,影响血管壁细胞、软骨和骨细胞等多种细胞在发育期的增殖、迁移,并影响细胞与基质的黏附,在机体心血管系统和软骨发育,血管、骨骼系统和神经肌接头(NMJ)的生成与功能调控等多种生命过程中均具有重要作用。2010年5月~2011年12月,我们对串珠素基因片段进行克隆、扩增和蛋白表达,以获取串珠素蛋白,并探讨其在咀嚼肌中的表达特点。
1 材料与方法
1.1 材料 探针载体直接转化,扩增,酶切,测序后,酶切片段,插入pET32a,表达,测序,表达全菌蛋白,Wetstern blot 1—测 his,Western blot 2—测抗串珠素,2次均在同一分子量处发现表达。牛串珠素基因(b串珠素)探针为Ray Rodgers教授(Reproductive Medicine Unit,Obstetrics & Gynaecology,Adelaide University,SA,AUSTRALIA,5005)惠赠,全长183 bp,连接于pGEM-T Easy载体。
1.2 实验方法
1.2.1 目的基因扩增和鉴定 用b串珠素-pGEMT Easy质粒转化感受态大肠杆菌DH5α(质粒0.1 μg,DH5α 感受态菌200 μL,冰浴30 min后42 ℃热休克90 s,),涂布于(含 Amp 100 μg/mL)的 LB 平板上,37℃培养过夜。挑阳性菌,将阳性细菌在含Amp的琼脂培养基中划板,挑取单克隆菌落接种于含Amp LB培养液中37℃振荡过夜。扩增后,用碱裂解法提取质粒,酚/氯仿抽提回收质粒,EcoR I+Sal I酶切,琼脂糖凝胶电泳分析鉴定,并送DNA测序。
1.2.2 目的基因克隆和表达 将测序正确的串珠素片段插入到表达载体 pET32a中(DNA片段2.5 μL,载体 2.5 μL,0.1 M MgCl20.5 μL,Sal I 5 μL,16℃,4 h),转化感受态大肠杆菌DH5α,接种于AMP+LB平板。37℃培养过夜。挑取单克隆菌落接种于LB培养液中,37℃振荡培养4 h。回收质粒,酶切,琼脂糖凝胶电泳鉴定。DNA测序鉴定。鉴定正确后,挑选1%转化后的细菌接种在新鲜的LB培养基中,37℃培养至对数生长期,0.2 mM IPTG诱导4 h,收菌,SDS-PAGE分析鉴定。以pET32a空载体转化和未经诱导的细菌设为对照组。
1.2.3 融合蛋白表达检测 采用Western blot法。表达全菌蛋白SDS-PAGE,200 mA低温转膜;5%脱脂奶粉封闭1 h后,鼠抗His抗体(1∶1 000)4℃封闭过夜。PBST(PBS中加入0.1%Tween-20)洗膜3次,每次10 min;羊抗鼠二抗(1∶5 000)封闭1 h;PBST洗膜3次,每次10 min;PBS洗膜3次,每次5 min;发光,显影。阴性对照为pET32a诱导和未诱导菌蛋白。再次行Western blot,一抗采用鼠抗串珠素单抗,检测融合蛋白表达情况。阴性对照仍然设为pET32a诱导和未诱导菌蛋白。
1.2.4 b串珠素抗血清制备 对雄性新西兰家兔进行常规体检,将重组的串珠素核心蛋白500 μg用PBS稀释至1 mL,与等体积的Freund完全佐剂充分乳化后,在家兔皮下注射1 mL,7 d后再次在双后腿淋巴结内注射100 μg抗原以加强免疫。以后每隔10 d进行一次加强免疫,5 W后取家兔全血,4℃过夜,4 000 r/min,4 ℃离心10 min,收集血清,-20 ℃保存。配置0.1 g/L琼脂糖凝胶,行双向免疫扩散试验,测定抗体滴度。
1.2.5 免疫组化实验 将成年大鼠脱颈处死,立即取材面部咀嚼肌,多聚甲醛固定过夜,梯度酒精脱水,石蜡包埋,5 μm石蜡切片。串珠素多克隆抗体工作浓度为1∶100,二抗为羊抗兔IgG(SABC免疫组化试剂盒,博士德公司,武汉),工作浓度为1∶100。切片脱蜡至水,在室温下用H2O2处理30 min,PBS洗5 min×2次,胰蛋白酶消化37℃ 30 min,PBS洗5 min×3次,50 mL/L正常羊血清封闭10 min,PBS洗5 min×3次。加一抗,对照组用正常羊血清代替一抗,4℃过夜。PBS洗5 min×3次,加二抗,37℃30 min。PBS洗5 min×3次,加SABC 37℃ 30 min。PBS洗5 min×3次,DAB显色,苏木素轻度复染。脱水,透明,封片,光镜观察。
2 结果
2.1 串珠素扩增和插入到表达载体后酶切电泳质粒扩增后酶切鉴定,琼脂糖凝胶电泳显示在100~200 bp间有新条带出现(见图1)(DNA marker:SD012)。扩增后所获得的片段送DNA测序(宝泰克公司),测序结果与原序列相同,说明扩增正确。将测序正确的串珠素片段插入到表达载体pET32a中,EcoR I+Sal I酶切后电泳鉴定,在100~200 bp处有稳定电泳条带出现,显示串珠素片段插入到pET32a中的连接反应正确(DNA marker:DGL2000,见图2)。
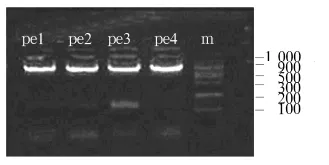
图1 b串珠素扩增后酶切电泳图
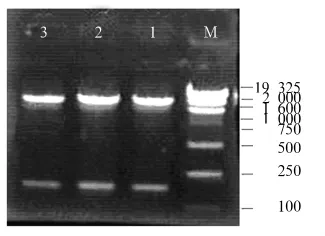
图2 串珠素片段插入到表达载体pET32a后酶切电泳图
2.2 串珠素蛋白表达和分析 SDS-PAGE显示在IPTG诱导的串珠素-pET32a转化菌蛋白有新条带出现,新条带的分子量在35 kD附近,说明IPTG诱导下插入pET32a的串珠素可以进行高效的蛋白表达。对照组分别为未经IPTG诱导的串珠素-pET32a转化菌和用pET32a空载体的转化菌蛋白。Western blot示,未诱导的菌无表达,而融合蛋白的和pET32a转化菌经诱导后均有蛋白表达,其中融合蛋白条带相对分子质量大于pET32a诱导的蛋白条带,说明插入pET32a载体的蛋白成功表达(见图4)。
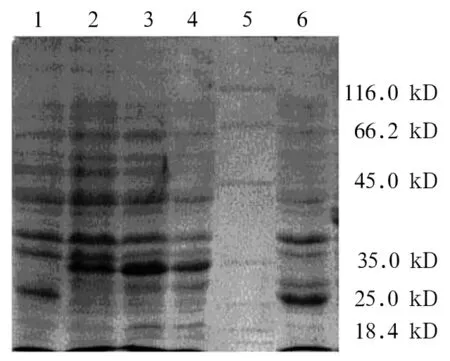
图3 SDS-PAGE分析串珠素表达
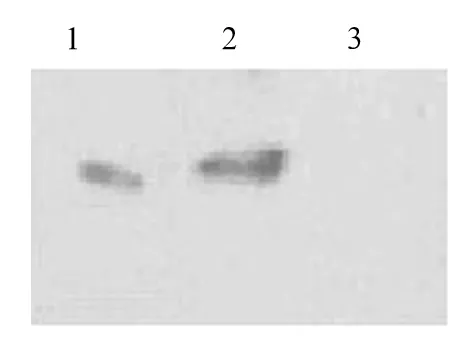
图4 Western blot显示的融合蛋白表达
2.3 免疫组化染色结果 光镜下观察,串珠素在咀嚼肌中表达显著,均匀分布在骨骼肌肌纤维细胞外的基质中,尤其在肌膜处明显聚集呈带状分布(图5)。

图5 串珠素在骨骼肌中的分布
3 讨论
串珠素是ECM中主要的蛋白聚糖之一[1],分子量约为700 kD,由核心蛋白和4条硫酸肝素(HS)侧链组成,与其他基质成分紧密结合并自行组成二聚体和大的聚合物。串珠素主要存在于骨、软骨、血管、牙胚中,介导着细胞增殖分化及细胞信号转导等[2,3]。组织来源不同的串珠素硫肝素侧链不同,不同种属的哺乳动物类串珠素核心蛋白序列上有同源性,不同组织细胞来源的串珠素侧链结构差别较大,侧链的变化对于其生物学活性具有明显的影响。串珠素侧链的HS带有负电,通过其吸水性增加组织的容积,可以结合、释放生长因子,使生长因子暂时失活,并在必要时恢复其活性,不仅具有生长因子储存库的作用,也是细胞黏附分子的重要配体。
骨骼肌再生是一个至今仍未完全被理解的过程,运动神经与骨骼肌间建立突触连接的位置是运动神经与肌细胞相互诱导来确定的,长期以来肌纤维周围的基膜一直被认为是这种调控作用的重要因素,但具体机制了解较少。近年来,随着对NMJ中ACh受体(AChR)复合体研究的深入,揭示出基膜中的多种硫酸乙酰肝素蛋白多糖(HSPG)在此过程中起主导作用。运动神经末梢通过释放的神经递质可诱导突触后膜上信号蛋白的积聚,使AChR复合体定位在突触后膜上,其中串珠素是该过程中最重要的一种信号蛋白,被认为是AChR复合体在运动终板突触后膜上定位的信号分子[4]。串珠素在NMJ处高表达,是NMJ处乙酰胆碱酯酶(AChE)的独特受体分子,在把AChE定位在突触基膜上的过程中发挥主要的作用[5]。AChE复合体在突触后膜的定位是通过串珠素结合AChE和dystroglycan来达到的[6]。体外实验已证实,在运动神经—肌肉共培养模型中,串珠素的存在能够明显促进二者间的相互诱导生长及突触的形成。在NMJ形成后,串珠素还继续对维持突触的稳定发挥作用[7]。同时,串珠素还参与了骨骼肌再生过程中的复杂的调控机制,因其是细胞表面和ECM的主要成分,调控生长因子活性并影响细胞生长和分化[8]。
骨骼肌损伤后,变性的肌纤维释放出多种信号,浸润的炎细胞和重塑的ECM均能影响修复性肌形成的过程。在多种信号存在的情况下,卫星细胞活化、肌前体细胞的增生和分化等一系列过程都需要密切调控。Casar等[9]报道了注射氯化钡诱导产生的小鼠骨骼肌细胞再生过程中观察的HSPG表达情况,发现有几种HSPG出现暂时性上调,包括syndecan-3、glypican、串珠素和syndecan-4。这些HSPG的表达增加与分化过程中早期标志物的表达相一致,并定位于新形成的肌管中。对缺血或毒性损伤,新形成的肌纤维在最初的基膜管中发育,这样可以让NMJ在原来的突触位置形成。这种基膜位置信息的储存依赖于HSPG,如agrin或串珠素。串珠素、glypican、syndecan-3和 probably syndecan-4是伴随新形成的正在分化的肌管能被原位检测到的几种主要HSPG。体外结合试验证实,ECM组蛋白H1能特异性结合串珠素,二者共同存在于再生的骨骼肌中,包括肌管培养基和再生骨骼肌的ECM。组蛋白H1与串珠素结合能显著刺激成肌细胞的增生[10]。
本实验设计通过基因工程方法表达串珠素核心蛋白,并制备特异性多克隆抗体,用以检测串珠素在咀嚼肌等口腔组织中的表达和分布。本研究首先完成了对b串珠素基因的正确克隆和表达,建立了可以表达b串珠素的组织工程菌。pET32a载体上串珠素插入区的上游有His蛋白的表达序列,可作为在Western blot中检测pET32a表达的标记。在以鼠抗His抗体作为一抗进行Western blot检测时,融合蛋白的和pET32a转化菌经诱导后均有蛋白表达,其中融合蛋白条带相对分子质量大于pET32a诱导的蛋白条带;而在以鼠抗串珠素为一抗进行Western blot检测中,只有融合蛋白转化菌经诱导后有蛋白表达,证明串珠素-pET32a转化菌蛋白能表达串珠素,并能与鼠抗串珠素特异性结合。
本实验通过条件诱导方法实现了串珠素核心蛋白的原核表达,而其较复杂的蛋白糖基化过程需要通过真核动物细胞表达来实现。用anti-串珠素单抗进行的Western blot表明原核表达的串珠素核心蛋白具有天然串珠素的免疫原性。因此本实验的方法仅是利用其具有的免疫原性来实现抗体制备,而不能用来产生活性蛋白。免疫沉淀实验证明了用其制备的多克隆血清可对串珠素核心蛋白产生特异性结合。另外,免疫组化实验表明牛与鼠串珠素的免疫原性之间存在交叉,应为哺乳动物间串珠素序列的同源性所致。
串珠素通过参与基底膜成分的结合与组装,对于维持基底膜结构与功能的完整性发挥着极其重要的作用。串珠素广泛表达在咀嚼肌的ECM中,其中在肌膜等ECM的膜性结构中更是集中表达。
[1]Kuo C,Lim S,King NJ,et al.Rhinovirus infection induces extracellular matrix protein deposition in asthmatic and nonasthmatic airway smooth muscle cells[J].Am J Physiol Lung Cell Mol Physiol,2011,300(6):951-957.
[2]Govindraj P,West L,Koob TJ,et al.Isolation and identification of the major heparan sulfate proteoglycans in the developing bovine rib growth plate[J].J Biol Chem,2002,277(22):19461-19469.
[3]Handler M,Yurchenco PD,Iozzo RV.Developmental expression of perlecan during murine embryogenesis[J].Dev Dyn,1997,210(2):130-145.
[4]Mishra M,Naik VV,Kale AD,et al.Perlecan(basement membrane heparan sulfate proteoglycan)and its role in oral malignancies:an overview[J].Indian J Dent Res,2011,22(6):823-826.
[5]Rotundo RL,Rossi SG,Kimbell LM,et al.Targeting acetylcholinesterase to the neuromuscular synapse[J].Chem Biol Interact,2005,(157-158):15-21.
[6]Smirnov SP,Barzaghi P,McKee KK,et al.Conjugation of LG domains of agrins and perlecan to polymerizing laminin-2 promotes acetylcholine receptor clustering[J].J Biol Chem,2005,280(50):41449-41457.
[7]Ohno K.Genetic defects and disorders at the neuromuscular junction[J].Brain Nerve,2011,63(7):669-678.
[8]Bajanca F,Luz M,Raymond K,et al.Integrin alpha6beta1-laminin interactions regulate early myotome formation in the mouse embryo[J].Development,2006,133(9):1635-1644.
[9]Casar JC,Cabello-Verrugio C,Olguin H,et al.Heparan sulfate proteoglycans are increased during skeletal muscle regeneration:requirement of syndecan-3 for successful fiber formation[J].Cell Sci,2004,117(1):73-84.
[10]Henriquez JP,Casar JC,Fuentealba L,et al.Extracellular matrix histone H1 binds to Perlecan,is present in regenerating skeletal muscle and stimulates myoblast proliferation[J].Cell Sci,2002,115(10):2041-2051.
猜你喜欢
石材(2020年10期)2021-01-08
创新作文(1-2年级)(2017年10期)2018-04-16
数学小灵通(1-2年级)(2017年5期)2017-06-05
创新作文(小学版)(2017年28期)2017-03-21
中国运动医学杂志(2016年3期)2016-07-10
中国运动医学杂志(2016年3期)2016-07-10
系统工程与电子技术(2016年2期)2016-04-16
医学研究杂志(2015年5期)2015-06-10
中国光学(2015年1期)2015-06-06
海岸工程(2014年4期)2014-02-27
