
东、西方蜜蜂对胡蜂气味辨识比较研究
2012-11-28 09:58汪正威曲玉凤胡宗文
中国蜂业 2012年1期
汪正威 曲玉凤 杨 爽 胡宗文 谭 垦
(云南农业大学东方蜜蜂研究所,昆明 650201)
嗅觉是蜜蜂等昆虫的主要感知系统,蜜蜂不仅能通过灵敏的嗅觉来辨别和采集不同种类的花蜜和花粉,还能通过体表信息素的气味来辨认巢伴和非巢伴。蜜蜂的嗅觉在学习行为、亲属辨认、工蜂监督和防御行为等行为上起着重要作用[1]。
蜜蜂是虫媒花植物理想授粉者,植物花通过散发出芳香的气味来吸引蜜蜂等授粉者。自从von Frisch首次发现蜜蜂经过几次往返于食源和蜂巢之间,能形成规律性采集,并能招引同伴去采集的现象[2]。研究者就开始集中研究蜜蜂对食物的嗅觉联想学习,在Kuwabara报道用颜色作为条件刺激下有PER的启发下[3],Takeda第一次用嗅觉PER条件学习测试蜜蜂的嗅觉辨别能力,将一种气味和糖水奖励相配合训练之后,再通过给予其他不同种气味但不配合糖水奖励刺激,来观察蜜蜂对不同气味之间的辨别和选择[4-5]。之后,1983年Bitterman等人研究蜜蜂通过联想学习一个条件刺激(气味)和非条件刺激的联系,使得该实验模型成为了之后研究蜜蜂行为学、神经学的经典试验模型[6]。
在黑暗的蜂箱当中,蜂王在各个蜂脾之间寻找空巢房产卵时,周围的工蜂会围绕蜂王,通过触角轻触蜂王来获取蜂王信息素,并能将该信息传递给巢内的其他工蜂,而蜂王爬过的蜂脾也同样能传递蜂王信息素存在的信息[7-9]。工蜂通过嗅觉感知蜂王信息素的存在来感知蜂王的存在。同样,工蜂还能依据不同化学气味来辨识与自己亲缘关系较近的工蜂,因为气味是受遗传控制,亲缘关系越近的工蜂,气味越相似[10]。
基于嗅觉信号的非巢伴辨识是社会性昆虫防御的基础[11-13]。在缺蜜季节,往往会发生盗蜂现象,在被盗群巢门口经常能见到守卫蜂的防御,对归巢蜂进行辨认防止偷盗[14]。也有研究称蜜蜂能够辨别出食物上是否存在捕食者——蜘蛛,如果存在被捕食的危险,蜜蜂则会放弃对该食源的采集[15]。尽管对蜜蜂的这种行为的本质原因不是很清楚,这就吸引了更多的研究者对蜜蜂防御性行为进行研究。
东、西方蜜蜂是我国现目前广泛饲养的两大蜂种,近来对东、西方蜜蜂的研究也逐渐深入。但是,对二者之间的学习记忆行为、防御行为的研究相对较少。在云南山区,东、西方蜜蜂总是生活在胡蜂的袭扰之下,东方蜜蜂作为本地蜂种与外来引入蜂种——西方蜜蜂对胡蜂存在不同的防御策略[16],二者是否对胡蜂气味存在不同的辨识能力呢?为了研究东、西方蜜蜂对胡蜂的不同防御行为,本研究通过研究二者对胡蜂气味的辨识能力,以期为其防御行为提供理论参考。
1 材料方法
1.1 Y形管试验
蜂群:实验所用东、西方蜜蜂蜂群均取自饲养于云南农业大学东方蜜蜂研究所院内实验蜂场内的健康蜂群。分别选取东方蜜蜂和西方蜜蜂蜂群各三群,群势较强,在实验过程中能有大量蜜蜂出巢采集的蜂群。
共同实验操作和共用Y形管:Y形管分支长15cm,一个入口,两个出口。分别在两个出口处接两个透明小盒子,用于盛装气味同时也尽量避免外界环境的影响。实验过程中,首先,用试管在巢门口捕获出巢采集的采集蜂10~15只,用棉球塞上带回实验点。将蜜蜂从入口处依次放入,蜜蜂在Y形管的分支处会根据前方的气味而做出选择。根据蜜蜂对不同气味的选择出口作详细记录。每次实验交替的时候,都调换两个出口的位置来中和之前蜜蜂爬行过程中留下的气味或相关信息素的作用。重复5次实验,之后交换两种气味的位置,以减少实验用蜜蜂存在位置偏好。
空白对照,Y形管的两个出口处都接没有放入任何气味的瓶子。观察蜜蜂在Y形管中是否存在位置偏好性。
实验组,Y形管的一个出口接气味,另一个出口仍接空瓶,观察蜜蜂在只有一种气味的Y形管中的选择情况。
在Y形管实验中,我们所用的胡蜂气味——每次选用了活体胡蜂用于气味辨识实验。对胡蜂剪翅去腿处理,同时还保持胡蜂的生存活性。
1.2 数据统计
采用Microsoft Office Excel 2003进行数据整理和数据处理,绘制图表。用卡方检验蜜蜂在Y形管出口选择上的差异,分析东、西方蜜蜂对胡蜂气味的辨识能力差异显著性。
2 结果与分析
2.1 空白对照
为了解蜜蜂对Y形管左右位置是否存在偏向性,设置了空白对照组,Y形管的左右两个出口都没有气味,观察蜜蜂的偏向性。
2.1.1 东方蜜蜂的Y形管空白实验
东方蜜蜂三群(n=248)用于空白实验,其中,从左边离开的有171只,占69.0%;从右边离开的有77只,占31.0%,与假设的50% vs 50% 相比,差异极显著(χ2=18.478, P<0.001),说明东方蜜蜂具有明显的向左偏向性。
2.1.2 西方蜜蜂的Y形管空白实验
西方蜜蜂三群(n=215)用于空白实验,其中,从左边离开的有124只,从右边离开的有91只,与假设的50%相比:差异不显著(χ2=2.553,P=0.110),说明西方蜜蜂没有明显的左右的偏向性。
2.1.3 东、西方蜜蜂左右偏向性比较
东方蜜蜂和西方蜜蜂二者之间比较发现,东方蜜蜂较西方蜜蜂有更强的“左”偏向,西方蜜蜂没有偏向性(χ2=6.335, P=0.012)。
2.2 对胡蜂气味的辨识比较
当Y形管其中的一个出口处放置处理过的胡蜂,观察蜜蜂在Y形管的选择,对其中的胡蜂的辨识和趋避性。
2.2.1 东方蜜蜂对胡蜂气味的辨识
当胡蜂放置在Y形管的左边出口处,选用东方蜜蜂三群(n=482)用于试验,其中,180只蜜蜂选择从含有胡蜂气味的左边出口离开,仅占37.34%,而相对更多(62.66%)的蜜蜂从右边空白出口离开。与空白对照比较,二者差异极显著(χ2=20.554, P<0.001)。东方蜜蜂对胡蜂气味存在明显趋避性。

表1 东、西方蜜蜂对胡蜂趋避性结果
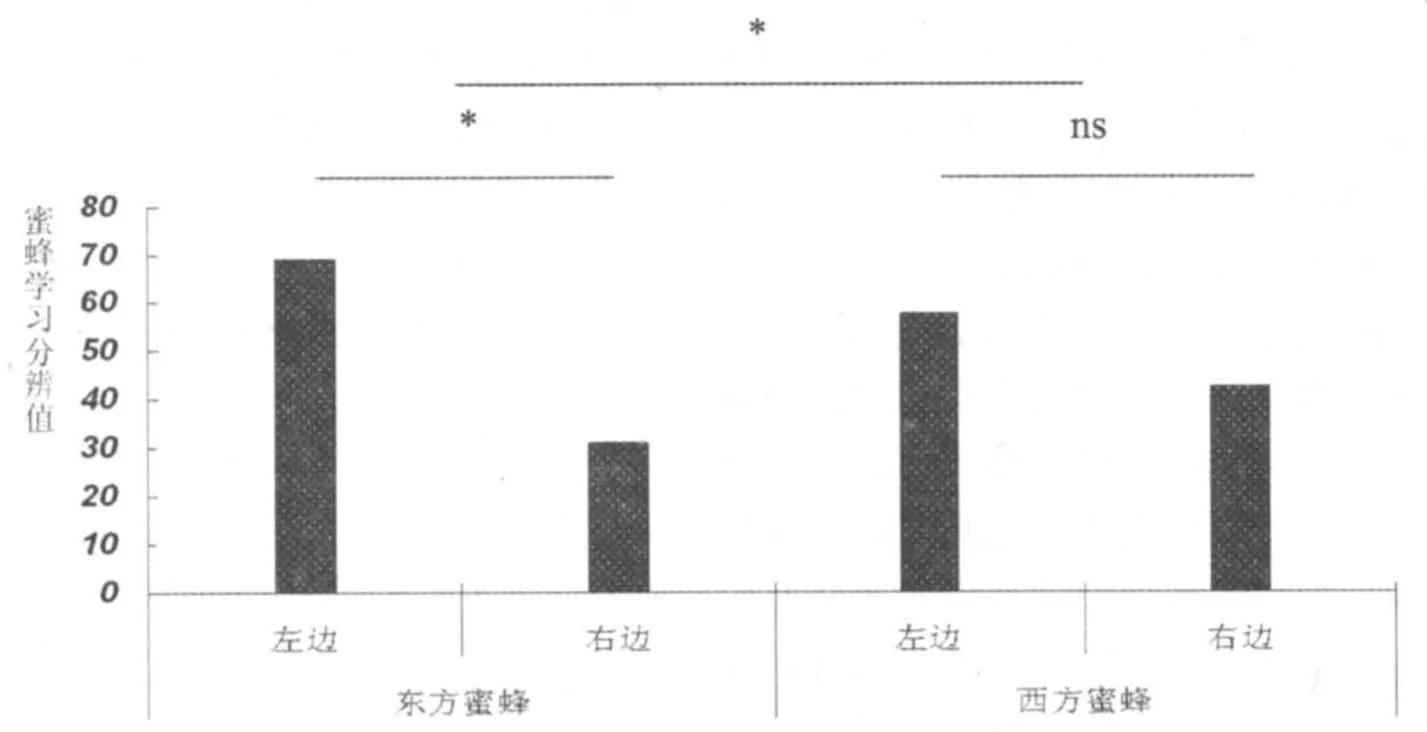
图1 东、西方蜜蜂偏向性结果*为差异显著P<0.05,ns 为差异不显著。

图2 东、西方蜜蜂对胡蜂气味的辨识比较
当胡蜂放置在Y形管的右边出口处,选用东方蜜蜂三群(n=463)用于试验,其中,也仅有101只选择从含有胡蜂气味的右边出口离开,占21.81%,而78.19%从左边空白出口离开。与空白对照比较,二者差异不显著(χ2=2.079, P=0.149)。东方蜜蜂对胡蜂趋避与其本身的明显左偏向性二者差异不显著。说明东方蜜蜂对胡蜂气味存在明显的趋避性。
2.2.2 西方蜜蜂对胡蜂气味的辨识
当胡蜂放置在Y形管的左边出口处,选用西方蜜蜂三群(n=379)用于试验,其中,153只蜜蜂选择从含有胡蜂气味的左边出口离开,占40.37%,59.63%的蜜蜂从右边空白出口离开。与空白对照比较,二者差异显著(χ2=6.483, P=0.011),西方蜜蜂对胡蜂气味存在趋避性。
当胡蜂放置在Y形管的右边出口处,选用西方蜜蜂三群(n=349)用于试验,其中,也仅有102只选择从含有胡蜂气味的右边出口离开,占29.23%,而70.77%从左边空白出口离开。与空白对照比较,二者差异不显著(χ2=3.690, P=0.055)。西方蜜蜂对胡蜂气味的趋避未达到显著水平。
2.2.3 东、西方蜜蜂对胡蜂气味的辨识比较
将Y形管左右出口的数据平均,消除左右方向偏向性的影响后。比较东、西方蜜蜂对胡蜂气味的辨识能力。数据如表.示,东、西方蜜蜂对胡蜂均有较强的趋避性。其中,东方蜜蜂对胡蜂的趋避率比西方蜜蜂高(70.26%>64.97%),尽管二者在统计上差异不显著(χ2=0.570, P=0.450)
3 讨论
通过比较东、西方蜜蜂对胡蜂气味的辨识和趋避性,我们发现:(1)东方蜜蜂具有较强的左偏向性,而西方蜜蜂方向偏向性不强;(2)东、西方蜜蜂对胡蜂气味都有较好的辨识能力,均会产生趋避性。我们的结论也从另一方面给前人的研究提供了类似的证据。谭垦等研究发现当有胡蜂试图在东、西方蜜蜂巢门口伺机捕食时,巢门口的守卫蜂数量会增加,说明东、西方蜜蜂都能判断有胡蜂接近。就东、西方蜜蜂对气味之间的辨别比较研究较少,前人的研究多集中在西方蜜蜂对气味的辨识和学习上,而对东、西方蜜蜂之间就感官和学习能力上的比较仅查到通过比较东方蜜蜂和西方蜜蜂之间对颜色的辨别能力,结果表明东方蜜蜂的识别能力更强[16-17]。
东方蜜蜂作为本土饲养的蜂种,与其捕食者——胡蜂之间二者协同进化,所以东方蜜蜂守卫蜂在防御胡蜂捕食时,形成了其特有的防御方式,其对胡蜂气味也相对敏感。而西蜂蜜蜂自1896年才由于良好的生产性能和对环境的较强适应力[18],逐渐在我国广泛推广。尽管二者存在很多相似,但不同种间由于长期的进化,还是存在一些细微差异,例如:体色、个体大小、采食半径等。有报道称如十字花科、唇形科的某些种类植物只能由东方蜜蜂采集[18],东方蜜蜂对胡蜂捕食进化出了较明显的防御方式,首先通过召集大量的守卫蜂,在巢门口通过抖动身体来防御胡蜂,之后还能通过结团将胡蜂热死其中;而西方蜜蜂则多为通过召集守卫蜂堵塞巢门口,或者是单独对胡蜂进行攻击撕咬等较简单的防御方式[17]。东方蜜蜂比西方蜜蜂对胡蜂气味更敏感,可能与二者的生活环境有很大相关性,二者与环境之间的进化稍微有些差异。
影响蜜蜂对气味辨识的影响因子很多,其中气味的浓度会影响蜜蜂的辨识能力,气味种类 会影响蜜蜂的辨识能力。研究发现蜜蜂能够通过多种气味中的一种主要成分的味道辨别并对该气味进行学习和记忆。本研究中用到的是活体胡蜂用于比较东、西方蜜蜂对胡蜂气味之间的辨识差异性,成分和浓度现目前不是很明确,有待今后进一步的研究。
东、西方蜜蜂是我国当前饲养的两种主要的蜜蜂,本研究中通过比较二者在Y形管中对胡蜂气味的辨识能力,发现东方蜜蜂比西方蜜蜂对胡蜂更敏感,但二者没有统计学上的差异。其中,东、西方蜜蜂对胡蜂气味均有趋避性这一现象为研究蜜蜂防御性行为提供了参考。
[1]Breed M D.Nestmate recognition in honey bees[1].Animal Behaviour,1983,31, 86-91.
[2]Von Frisch K.Tanzsprache und Orientierung der Bienen,1965,Springer-Verlag.
[3]Kuwabara M.Bildung des bedingten Reflexes von Pavlovs Typus bei der Honigbiene, Apis mellifica[J].J Fac Sci Hokkaido Univ Ser VI Zool, 1957, 13, 458-464.
[4]Takeda K.Classical conditioned response in the honey bee[J].Journal of Insect Physiology, 1961, 6, 168-179.
[5]Reith M, Baumann G, Cla en-bockhoff R,Speck T.New insights into the functional morphology of the lever mechanism of Salvia pratensis(Lamiaceae)[J].Annals of botany, 2007, 100, 393.
[6]Vareschi E.Duftunterscheidung bei der Honigbiene Einzelzell-Ableitungen und Verhaltensreaktionen[J].Journal of Comparative Physiology A: Neuroethology, Sensory, Neural, and Behavioral Physiology, 1971, 75, 143-173.
[7]Butler C.The method and importance of the recognition by a colony of honeybees (A.mellifera) of the presence of its queen[J].Transactions of the Royal Entomological Society of London, 1954,105, 11-29.
[8]Sakagami S F.The false-queen: fourth adjustive response in dequeened honeybee colonies[J].Behaviour, 1958, 13, 280-296.
[9]Menzel R.Associative learning in honey bees[J].Apidologie, 1993,24,157-157
[10]Getz W M,Page JR R E.Chemosensory Kin-communication Systems and Kin Recognition in Honey Bees[J].Ethology, 1991,87, 298-315.
[11]Dani F R, Jones G R, Corsi S, et al.Nestmate recognition cues in the honey bee: differential importance of cuticular alkanes and alkenes[J].Chemical senses, 2005, 30, 477.
[12]Heiling A,Herberstein M.Predator-Cprey coevolution: Australian native bees avoid their spider predators.Proceedings of the Royal Society of London[J].Series B: Biological Sciences, 2004, 271, S196.
[13]Li J, Tan K,He S.Hornet predation risk: apperception and response by honey bees[J].Journal of apicultural research, 2008, 47, 174-175.
[14]Pahl M, Tautz J, Zhang S.Honeybee cognition[J].Animal Behaviour:Evolution and Mechanisms, 2010, 87-120.
[15]Reith M, Baumann G, Cla en-bockhoff R,et al.New insights into the functional morphology of the lever mechanism of Salvia pratensis(Lamiaceae)[J].Annals of botany, 2007, 100, 393.
[16]Tan K, Hepburn H, Radloff S, et al.Heat-balling wasps by honeybees[J].Naturwissenschaften, 2005, 92, 492-495.
[17]Tan K, Radloff S, Li J, et al.Bee-hawking by the wasp, Vespa velutina, on the honeybees Apis cerana and A[J].mellifera.Naturwissenschaften, 2007, 94, 469-472.
[18]杨冠煌.论引入西方蜜蜂对中蜂的危害及生态影响[J].昆虫学报,2005, 48, 401-406.
猜你喜欢
心理学报(2022年1期)2022-01-21
第二课堂(课外活动版)(2021年11期)2021-01-18
当代陕西(2020年23期)2021-01-07
中国蜂业(2020年7期)2020-08-21
当代陕西(2019年12期)2019-07-12
意林·少年版(2019年5期)2019-04-18
中国化肥信息(2019年1期)2019-04-03
中国蜂业(2017年10期)2017-10-21
意林·少年版(2017年5期)2017-04-13
意林·少年版(2014年13期)2014-08-27
