
U251和U87细胞的Ezrin蛋白生物信息学分析
2013-09-21 12:59刘乃杰梁华新秦治刚孙利波房晓萱赵丛海朱庆三
中国老年学杂志 2013年7期
刘乃杰 梁华新 秦治刚 孙利波 房晓萱 赵丛海 朱庆三
(吉林大学中日联谊医院,吉林 长春 130033)
应用蛋白质组学和免疫组化检测发现,在不同来源的转移性肿瘤中,Ezrin蛋白高表达〔1~3〕。相对于良性上皮组织膜顶端的表达,胞浆高表达的Ezrin与人类乳腺癌去分化、侵袭和不良预后有关〔4〕。应用shRNA和Ezrin负调控区突变抑制 Ezrin功能,可阻止侵袭和早期转移〔5~7〕。研究发现Ezrin基因过表达与儿童横纹肌肉瘤和骨肉瘤临床分期相关,移植瘤模型证实减少横纹肌肉瘤或骨肉瘤细胞系Ezrin的表达可导致肺转移减少〔8〕。神经胶质瘤因其浸润性生长,使手术及放化疗效果均不理想。Tynninen等〔9〕研究发现Ezrin同样在神经胶质瘤高表达,且与神经胶质瘤的进展相关,与组织学细胞类型和WHO肿瘤等级相关。但不同肿瘤等级、不同浸润性神经胶质瘤Ezrin表达量不同的机制不清楚。本文对侵袭性不同的胶质瘤细胞系U87和U251的Ezrin蛋白序列进行生物信息学分析,拟寻找浸润性生长的机制,为神经胶质瘤的治疗寻求新的途径。
1 材料与方法
1.1 主要试剂 组织RNA提取试剂盒、小量质粒提取试剂盒,北京博迈德科技发展有限公司;M-MLV反转录酶,Promega公司;Ribonuclease Inhibitor、dNTP Mixture、Oligo d(T)15Primers、Random Primers、TaKaRa LA Taq®Hot Start Version、pMD®18-T vector、DL2000 DNA Marker、DL10000 DNA Marker、限制性内切酶XhoⅠ和EcoRⅠ,宝生物工程(大连)有限公司;DEPC,Sigma公司。U251、U87细胞购自中国科学院细胞库。应用Primer Premier 5.0软件设计Ezrin基因的引物:EF216:5'-CTCGAGCTATGCCGAAACCAATCAATGTCCG-3',ER1976:5'-GAATTCTTACAGGGCCTCGAACTCGTCGATG-3',由上海生工生物工程技术服务有限公司合成。
1.2 基因序列及蛋白结构分析工具 Basic Local Alignment Search Tool(nucleotide blast及 blastx),EBI>Tools>Protein,SWISS-MODEL,SWISS-PDB Viewer。
1.3 方法
1.3.1 Transwell检测U251和U87细胞的迁移侵袭能力 取100 μl Matrix gel加至 Transwell小室中,置 37℃,待其凝固。0.25%胰酶消化收集细胞,计数,吸取100 μl(含1×105个细胞)加至Matrix gel表面,于37℃继续培养4~6 h,结晶紫染色,计数穿过膜底面的细胞。
1.3.2 Ezrin基因的钓取、克隆及鉴定 0.25%胰酶消化,收集细胞1×107,用组织RNA提取试剂盒提取细胞RNA,定量后,反转录合成 cDNA。反应体系:10 × buffer 2 μl,dNTPs(2.5 mmol/L)4 μl,RNase Inhibitor(40 U)0.5 μl,Oligo d(T)15Primers 1 μl,Random Primers 1 μl,RNA 1 μg,M-MLV 1 μl,补加free-RNase H2O至20 μl。反应条件:37℃ 1 h。取上述合成的cDNA PCR扩增Ezrin基因,配制25 μl反应体系:10倍缓冲液2.5 μl,dNTPs(2.5 mmol/L)4 μl,引物 EF216(10 pmol/L)、ER1976(10 pmol/L)各 1 μl,TaKaRa LA Taq®Hot Start Version 0.5 μl,cDNA 2 μl,ddH2O 16 μl。按下列条件进行反应:94℃30 s,58℃ 2 min,72℃ 30 s,35 个循环,72℃ 5 min。1% 琼脂糖凝胶电泳鉴定PCR产物并回收,进行TA克隆。37℃过夜培养后,挑取单菌落摇菌,小量提取质粒,XhoⅠ和EcoRⅠ双酶切鉴定。
1.3.3 Ezrin基因测序及序列分析 选取阳性质粒测序,由上海生工生物工程技术服务有限公司完成。应用Basic Local A-lignment Search Tool(nucleotide blast)分析所测得的序列,判断其与GenBank登陆的序列的相似性。再应用Local Alignment Search Tool(blastx)分析所测序列氨基酸变异情况。
1.3.4 U87和U251细胞Ezrin蛋白结构分析及预测 应用EBI>Tools>Protein和 SWISS-MODEL在线分析 U87和 U251细胞Ezrin蛋白功能、结构域、空间结构差异;应用SWISS-PDB Viewer分析变异氨基酸的位置。
2 结果
2.1 Transwell检测U251和U87细胞的迁移侵袭能力 Transwell检测结果可见,穿过膜底面的U87细胞为(35.5±6.89)个,明显高于U251细胞〔(13.3±3.01)个〕(P<0.001)。表明U87的迁移、浸润能力强。见图1。
2.2 U251和U87细胞系Ezrin基因的钓取、克隆及鉴定 1%琼脂糖凝胶电泳显示,RT-PCR扩增产物为1 700 bp,与Ezrin基因的大小相符(图2)。切胶回收该PCR产物,TA克隆。应用XhoⅠ和EcoRⅠ双酶切,电泳显示在图3a泳道10和12、图3b泳道2在1 700 bp处有一DNA条带,表明其质粒为阳性克隆载体。
2.3 Ezrin基因测序及序列分析 选取图3中的阳性质粒测序,应用Basic Local Alignment Search Tool(nucleotide blast)分析显示,所测得的U251和U87细胞Ezrin基因序列与GenBank登陆序列的相似性为99%,U251细胞Ezrin基因序列中有一个位点突变,U87细胞有5个位点。应用 Local Alignment Search Tool(blastx)分析Ezrin氨基酸,U251细胞Ezrin第407位氨基酸变异,U87细胞Ezrin在第66、258、265和577位变异。

图1 Transwell检测U251和U87细胞的迁移侵袭

图2 RT-PCR扩增U251和U87细胞系Ezrin基因
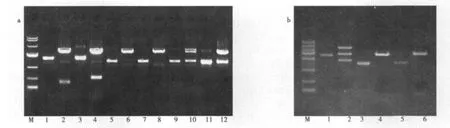
图3 XhoⅠ和EcoRⅠ酶切鉴定Ezrin基因的TA克隆
2.4 U87和 U251细胞 Ezrin蛋白功能、结构分析 应用EBI>Tools>Protein在线分析 U87和 U251细胞 Ezrin蛋白功能,与U87细胞 Ezrin匹配的蛋白有1 503个,与 U251细胞Ezrin匹配的蛋白却有362个。Ezrin的结构分析显示,蛋白质结构数据库PDB中U87细胞Ezrin的数量多于U251细胞,而蛋白质结构等级分类显示的结构域的结构分级和分层分级相同。U 87细胞Ezrin的结构域有45个,而U251细胞的Ezrin只有5个。
2.5 SWISS-MODEL建模分析变异氨基酸的位置 应用SWISS-MODEL分析,U87细胞Ezrin的3D空间结构有8处与U251细胞不同,QMEAN4评分U87细胞Ezrin低于U251细胞(表1)。SWISS-PDB Viewer显示U87细胞 Ezrin第258、265位氨基酸相对位于空间结构的内侧。

表1 Ezrin蛋白QMEAN4评分
3 讨论
Ezrin是ERM家族成员之一。ERM蛋白质是高度保守的〔10〕,从进化上,不同种类动物ERM都有着相同的蛋白质结构,由氨基末端FERM、富含α-螺旋区和含有F-肌动蛋白结合位点(30个氨基酸)的羧基末端组成,这些结构域的同源可达约76%。生化研究表明,ERM蛋白通过N-末端和C-末端约100个氨基酸称为N-/C-ERMAD的结构域相互作用,屏蔽其与肌动蛋白和膜的结合位点,对ERM蛋白进行负调控。X-射线显示FERM结构域由包括一个与羧基末端结合的折叠的PTB/PH/EVH1的三叶草状排列的F1、F2和F3,掩蔽一个大的FERM结构〔11〕。Li等〔10〕研究的最新数据表明丰富的α中央螺旋结构与FERM的F1和F2的连接区的相互作用对屏蔽结合位点,特别是与磷酸肌醇结合的位点具有重要作用。表明任何ERM蛋白的调控因子结合α-螺旋区可能会导致构象的变化,因此,在激活过程中也会出现。ERM蛋白的激活是由信号传导严格调控的。
越来越多的证据证明ERM蛋白在肿瘤进展中的作用。基因和蛋白质表达分析显示Ezrin在各种人类和啮齿动物肿瘤中的表达与良性的组织对比显著增加〔5,12~15〕。Ezrin在胞质表达量增加与人类乳腺癌去分化、侵袭性和不良预后有关〔3〕。并且在同一肿瘤的不同临床分期,Ezrin的表达量显著不同。大量研究证实Ezrin在肿瘤组织的激活、表达,参与肿瘤细胞迁移侵袭,是受信号分子调控的〔16~18〕。本研究推测Ezrin蛋白在肿瘤组织中变异影响其自身结构,从而导致细胞迁移侵袭能力不同。因此,本研究对具有不同侵袭能力的脑胶质瘤细胞U251和U87的Ezrin蛋白功能和结构进行生物信息学分析来加以证明。应用Basic Local Alignment Search Tool(nucleotide blast)分析显示,U251和U87细胞Ezrin基因序列与GenBank登陆序列的相似性为99%,U251细胞Ezrin基因序列中有一个位点突变,U87细胞的有5个位点,均导致相应的氨基酸变异,即U251细胞Ezrin第407位氨基酸变异,U87细胞Ezrin第66、258、265和577位变异。本研究表明U87细胞Ezrin氨基酸变异多于U251细胞,导致其功能、结构域及空间结构不同,从而影响U87、U251细胞的迁移和浸润能力不同。
1 Cui Y,Wu J,Zong M,et al.Proteomic profiling in pancreatic cancer with and without lymph node metastasis〔J〕.Int J Cancer,2009;124(7):1614-21.
2 Belbin TJ,Singh B,Smith RV,et al.Molecular profiling of tumor progression in head and neck cancer〔J〕.Arch Otolaryngol Head Neck Surg,2005;131(1):10-8.
3 Sarrió D,Rodríguez-Pinilla SM,Dotor A,et al.Abnormal ezrin localization is associated with clinicopathological features in invasive breast carcinomas〔J〕.Breast Cancer Res Treat,2006;98(1):71-9.
4 Baumgartner M,Sillman AL,Blackwood EM,et al.The Nckinteracting kinase phosphorylates ERM proteins for formation of lamellipodium by growth factors〔J〕.Proc Natl Acad Sci USA,2006;103(36):13391-6.
5 Yu Y,Khan J,Khanna C,et al.Expression profiling identifies the cytoskeletal organizer ezrin and the developmental homeoprotein Six-1 as key metastatic regulators〔J〕.Nat Med,2004;10(2):175-81.
6 Khanna C,Wan X,Bose S,et al.The membrane-cytoskeleton linker ezrin is necessary for osteosarcoma metastasis〔J〕.Nat Med,2004;10(2):182-6.
7 Elliott BE,Meens JA,SenGupta SK,et al.The membrane cytoskeleton crosslinker ezrin is required for metastasis of breast carcinoma cells〔J〕.Breast Cancer Res,2005;7(3):R365-73.
8 Niqqli V,Rossy J.Ezrin/radixin/moesin:Versatile controllers of signaling molecules and of the cortical cytoskeleton〔J〕.Int J Biochem Cell Biol,2008;40(3):344-9.
9 Tynninen O,Carpén O,Jääskeläinen J,et al.Ezrin expression in tissue microarray of primary and recurrent gliomas〔J〕.Neuropathol Appl Neurobiol,2004;30(5):472-7.
10 Li Q,Nance MR,Kulikauskas R,et al.Self-masking in an intact ERM-merlin protein:an active role for the central alpha-helical domain〔J〕.J Mol Biol,2007;365(5):1446-59.
11 Arpin M,Chirivino D,Naba A,et al.Emerging role for ERM proteins in cell adhesion and migration〔J〕.Cell Adh Migr,2011;5(2):199-206.
12 Bruce B,Khanna G,Ren L,et al.Expression of the cytoskeleton linker protein ezrin in human cancers〔J〕.Clin Exp Metastasis 2007;24(2):69-78.
13 Song J,Fadiel A,Edusa V,et al.Estradiol-induced ezrin over-expression in ovarian cancer:a new signaling domain for estrogen〔J〕.Cancer Lett,2005;220(1):57-65.
14K¨obel M,Langhammer T,Hüttelmaier S,et al.Ezrin expression is related to poor prognosis in FIGO stage I endometrioid carcinomas〔J〕.Mod Pathol,2006;19(4):581-7.
15 Elzagheid A,Korkeila E,Bendardaf R,et al.Intense cytoplasmic ezrin immunoreactivity predicts poor survival in colorectal cancer〔J〕.Hum Pathol,2008;39(12):1737-43.
16 Belkina NV,Liu Y,Hao JJ,et al.LOK is a major ERM kinase in resting lymphocys cytoskeletal rearrangement through ERM phosphorylation〔J〕.Proc Natl Acad Sci USA,2009;106(12):4707-12.
17 ten Klooster JP,Jansen M,Yuan J,et al.Mst4 and Ezrin induce brush borders downstream of the Lkb1/Strad/Mo25 polarization complex〔J〕.Dev Cell,2009;16(4):551-62.
18 Chirivino D,Del Maestro L,Formstecher E,et al.The ERM proteins interact with the class C-Vps/HOPS complex to regulate the maturation of endosomes〔J〕.Mol Biol Cell,2011;22(3):375-85.
猜你喜欢
中国现代医生(2022年19期)2022-08-25
湖北农业科学(2022年11期)2022-07-18
实用肿瘤学杂志(2020年4期)2020-12-08
趣味(数学)(2020年4期)2020-07-27
支部建设(2020年15期)2020-07-08
中国组织化学与细胞化学杂志(2017年1期)2017-06-15
百科知识(2015年18期)2015-09-10
中国当代医药(2015年9期)2015-03-01
中国老年学杂志(2015年9期)2015-01-31
肿瘤预防与治疗(2014年4期)2014-11-24
