
水分对番茄不同叶龄叶片光合作用的影响
2013-12-08 12:39陈凯利李建明贺会强胡晓辉姚勇哲孙三杰
生态学报 2013年16期
陈凯利,李建明,贺会强,胡晓辉,姚勇哲,孙三杰
(西北农林科技大学园艺学院, 杨凌 712100)
水分对番茄不同叶龄叶片光合作用的影响
陈凯利,李建明*,贺会强,胡晓辉,姚勇哲,孙三杰
(西北农林科技大学园艺学院, 杨凌 712100)
以番茄品种“金棚1号”为材料,采用盆栽方式,按照蒸腾蒸发量(ET)的50%、75%、100%和125%作为补充灌溉量研究了不同水分下番茄结果期叶片气体交换特性和光响应特征参数随叶龄的变化。结果表明:番茄叶片随着叶龄的增加,净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)逐渐降低,水分利用效率(WUE)呈先上升后下降趋势;叶龄为18 d 和29 d的叶片最大净光合速率(Pmax)随灌溉量的增加均先增加后降低,分别在75%ET和100%ET处理达到最大值。叶龄为38 d和47 d的叶片Pmax均以125%ET处理最大。表观量子效率(α)随叶龄的增大也先升高后下降,在叶龄为38 d 时最大;番茄叶片的光饱和点(LSP)随叶龄的加大而减小。不同水分处理下不同叶龄叶片的光响应特征参数为:叶片在叶龄为18 d时,Pmax为20.64—26.73 μmol·m-2·s-1,α为0.0518—0.0556;叶龄为29 d时,Pmax为11.00—24.24 μmol·m-2·s-1,α为0.0522—0.0594;叶龄为38 d 时,Pmax为11.77—18.18 μmol·m-2·s-1,α为0.0619—0.0693;叶龄为47 d时,Pmax为9.09—18.17 μmol·m-2·s-1,α为0.0538—0.0606。随叶龄加大,增加补充灌溉量有利于延缓叶片光合能力的降低。气孔限制是水分影响番茄叶片光合作用的主要因素,气孔限制与非气孔限制因素是番茄叶片Pn随叶龄变化的原因。
番茄;叶龄;水分;光响应特性;气体交换参数
在干旱和半干旱区,水分是作物光合作用的限制性因子[1]。水分胁迫后的植物光合速率下降[2- 3],水分胁迫下气孔因素和非气孔因素共同引起作物光合效率降低[4]。光响应模型是评价和模拟植物光合作用的物质生产以及生态系统生产能力的基础[5],常见的有直角双曲线模型[6]、非直角双曲线模型[7]、指数模型[8- 9]以及由这些模型发展起来的新模型[10]。由光响应曲线得到的最大光合速率(Pmax)和表观量子效率(α)是光合模型中两个重要的参数,是描述植物光合作用特征的重要生理生态特征参数,它因植物的种类、品种、生长条件和自身代谢状况的差异而不同[5]。
叶龄是叶片生命活动长短的标尺。因此,叶龄直接影响植物的光合作用,其大小影响叶片的光响应特征参数[11]。此外,水分不足会加速叶片的衰老,使叶片的生活期缩短。不同水分条件下相同叶龄的叶片光合性能也会呈现出差异性。因此,测定不同叶龄叶片在不同水分状况下的光响应特征参数为建立植物有关叶龄和水分的光合作用模型提供理论参数,也为植物在不同水分条件下光响应特征参数随叶龄变化的趋势提供理论依据。
目前国内番茄光合作用对水分的响应研究大多集中在气体交换参数[12- 18]和其日变化方面[13- 14,18],不同叶龄叶片光合作用的光响应特性对水分的响应鲜有报道。为此,本试验以番茄“金棚1号”为材料,研究不同补充灌溉量条件下塑料大棚番茄光合作用随叶龄的变化,为水分和叶龄有关的光合模型的建立提供理论参数。
1 材料与方法
1.1 试验材料与试验设计
试验于2011年3月至6月在陕西省杨凌区西北农林科技大学园艺学院塑料大棚中进行。供试材料为番茄品种“金棚1号”。试验采用盆栽方式进行,定植塑料花盆高48 cm,直径40 cm。定植基质为土壤和牛粪的混合物,定植前先将土壤和牛粪分别过筛,再按体积比1∶1配比,混合均匀后装入盆中,每盆16 kg。基质饱和重量含水量为40%。于3月24日番茄幼苗5叶1心时定植,每盆定植1株,共36盆,浇饱和水缓苗后常规栽培管理,于5月8日番茄进入结果期开始水分处理。
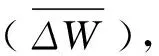
1.2 测定项目
1.2.1 基质重量含水量的测定
于5月8日开始每隔4 d在距离土壤顶端10 cm的土层取样,置于铝盒中,采用烘干法测定基质重量含水量,其公式为:基质重量含水量=(基质鲜质量-基质干质量)/基质干质量。
1.2.2 叶片光合作用光响应曲线的测定
试验分别于5月12日、5月23日、6月1日、6月10日测定4月24日标记的初展开叶片,即于水分处理4 d 后开始连续监测叶龄为18、29、38、47 d的叶片光响应曲线。使用LI-6400XT便携式光合仪(美国LI-COR公司),采用Li-6400-2B光源,设定气体流速为500 μmol/s,控制叶温25 ℃,设置光合有效辐射强度(PAR,μmol·m-2·s-1)梯度为2 000,1 700,1 400,1 100,800,500,200,150,100,50,20,0 μmol·m-2·s-1)。测定前用1 400μmol·m-2·s-1的PAR对叶片进行光诱导,使叶片活化。采用“Light-curve”程序自动测定,每个光强下稳定120 s。仪器自动记录每个光强下净光合速率(Pn,μmol·m-2·s-1)、蒸腾速率(Tr,mmol·m-2·s-1)、气孔导度(Gs,mol·m-2·s-1)和胞间CO2浓度(Ci,μmol·mol-1)等光合指标。番茄叶片水分利用效率(WUE,μmol/mmol)和气孔限制值(Ls)分别通过公式WUE=Pn/Tr和Ls=1-Ci/Ca[9]得到,其中Ca为空气CO2浓度(μmol/mol)。
1.3 数据处理
采用Microsoft Excel 2003软件对数据进行处理和绘图。
Peat[8]认为利用指数模型描述番茄叶片光合作用对光强的响应较好,因此本试验依据Bassman等[9]的指数模型拟合光响应曲线方程:
(1)
式中,Pmax为最大光合速率(μmol· m-2·s-1),α为表观量子效率,C0为一度量弱光下净光合速率趋近于0的指标。
按下式计算光补偿点(LCP):
LCP=Pmax×ln(C0)/α
(2)
假定Pn(净光合速率)达到Pmax的99%时,PAR为光饱和点(LSP),则有:
LSP=Pmax×ln(100C0)/α
(3)
运用DPS分析软件(Marquardt算法)拟合光响应曲线得到番茄叶片的Pmax、α、LCP、LSP等光响应特征参数。
2 结果分析
2.1 补充灌溉量对基质重量含水量的影响
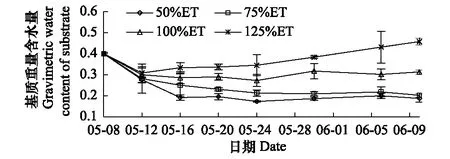
图1 不同补充灌溉量对基质重量含水量的影响 Fig.1 Effects of different supplementary irrigation amount on gravimetric water content of substrate
补充灌溉量的多少直接影响基质重量含水量的高低,基质重量含水量随补充灌溉量的增加而增加(图1)。随着水分处理时间的延长,各处理基质重量含水量都先快速下降,然后在一定的范围内波动;不同补充灌溉量下各处理基质水分含量的差异较为明显;其中125%ET处理在处理后期基质重量含水量升高,并使基质水分处于过饱和状态。
2.2番茄不同叶龄叶片气体交换参数对水分和光照的响应
2.2.1水分对番茄不同叶龄叶片净光合速率的影响
不同补充灌溉量条件下,番茄结果期不同叶龄叶片Pn随着PAR的增大的变化趋势一致,且随PAR的增强发生明显改变(图2)。总体来说,不同水分处理不同叶龄叶片Pn均在PAR增强初始阶段(PARlt;200 μmol·m-2·s-1,Pn呈线性增大;在PARlt;800 μmol·m-2·s-1)时,Pn随PAR的升高而迅速增大;当PAR继续增强时,Pn增大速度减缓或下降。在叶龄为18 d 时,在不同PAR条件下,Pn总是75%ETgt;100%ETgt;125%ETgt;50%ET,100%ET和125%ET处理Pn一直较为接近,而当PARgt;500 μmol·m-2·s-1时,50%ET处理与其他处理差异比较明显。与叶龄为18 d 的叶片Pn相比,番茄片叶龄为29 d 时,各水分处理Pn降低。在不同PAR条件下,Pn总是100%ETgt;125%ETgt;75%ETgt;50%ET,当PARgt;500 μmol·m-2·s-1时,各水分处理下的Pn的差异显著,50%ET处理在高光强(PARgt;1400 μmol·m-2·s-1)下Pn出现下降的趋势。当叶龄增至 38 d时,番茄叶片Pn进一步降低。当叶龄为 38 d 时,当PARlt;1100 μmol·m-2·s-1时,Pn总是125%ETgt;100%ETgt;75%ETgt;50%ET,当PARgt;1100 μmol·m-2·s-1时,75%ET处理出现光抑制现象,且下降速率比50%ET处理快。但一直以125%ET处理Pn最高。当叶龄增至 47 d 时,各水分处理Pn再次减小。在不同PAR条件下,Pn总是125%ETgt;100%ETgt;75%ETgt;50%ET。由此可知,随着番茄叶片叶龄的增加,Pn出现最大值的灌溉量不同。当叶龄增大时,需要较大的灌溉量来获得较高的Pn。

图2 水分对番茄不同叶龄叶片净光合速率光响应过程的影响Fig.2 Effect of water on light response of net photosynthetic rate for different leaf age tomato leaves
2.2.2 水分对番茄不同叶龄叶片气孔导度的影响
不同水分条件下不同叶龄番茄叶片的Gs均在PARlt;200 μmol·m-2·s-1时呈直线上升,随后缓慢升高(图3)。不同叶龄叶片在4个水分处理下Gs随PAR变化的趋势与Pn相似,这是因为气孔是植物叶片与外界进行气体交换的主要通道,O2、CO2、H2O都是通过气孔扩散,其闭合程度直接影响Pn和Tr,间接影响WUE。

图3 水分对番茄不同叶龄叶片气孔导度光响应过程的影响Fig.3 Effect of water on light response of stomatal conductance for different leaf age tomato leaves
2.2.3 水分对番茄不同叶龄叶片蒸腾速率的影响
图4表明不同叶龄番茄叶片的4个水分处理的Tr随PAR的升高变化的趋势与Pn和Gs随PAR的变化极为相似。番茄叶片叶龄为18 d 时,50%ET处理叶片的Tr明显低于其他3个处理;当叶片叶龄为29 d 时,各水分处理下Tr降低。不同PAR下,不同水分处理的Tr差异明显,总是100%ETgt;125%ETgt;75%ETgt;50%ET。当叶片叶龄增至38 d 时,Tr再次降低。番茄叶片叶龄达到47 d时,Tr降至最低。
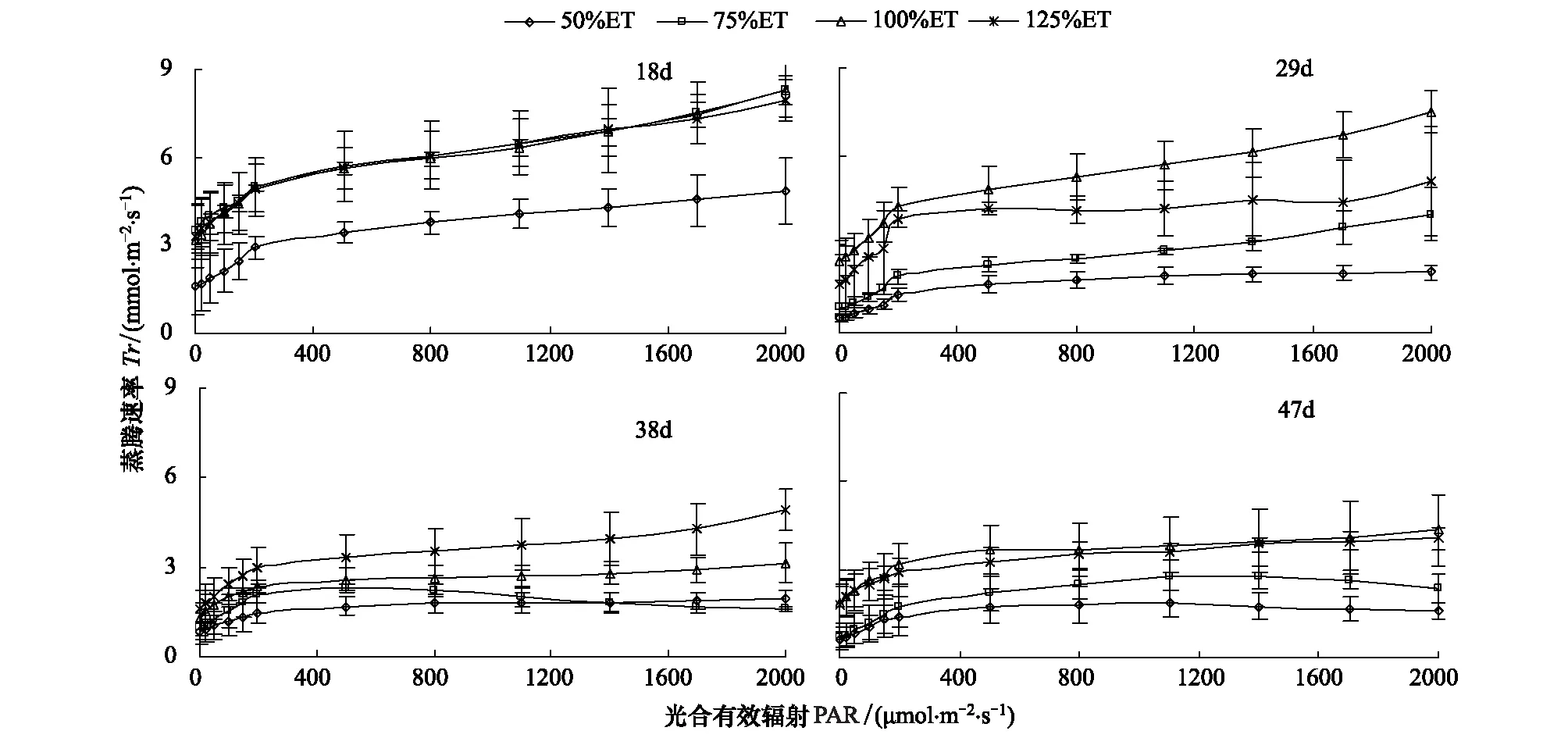
图4 水分对番茄不同叶龄叶片蒸腾速率光响应过程的影响Fig.4 Effect of water on light response of transpiration rate for different leaf age tomato leaves
2.2.4 水分对番茄不同叶龄叶片水分利用效率的影响
由图5可知,不同叶龄番茄叶片的WUE在PARlt;500 μmol·m-2·s-1时,4个水分处理下WUE均随PAR的增强而急剧上升;当PARgt;800 μmol·m-2·s-1,WUE随PAR的增强逐渐减小。50%ET处理WUE较其他3个处理高,且其他3个水分处理间的WUE相近;当番茄叶片叶龄增加到29 d时,各水分处理WUE均提高;当叶龄达到38 d时,除125%ET处理WUE不再增加,其他3个处理WUE均继续增加。最后当叶龄为47 d时,各水分处理WUE下降,且相差不大。总之,除125%ET处理WUE随叶龄的变化不大,其他3个处理WUE均随叶龄的增大而呈先增加后降低的趋势。
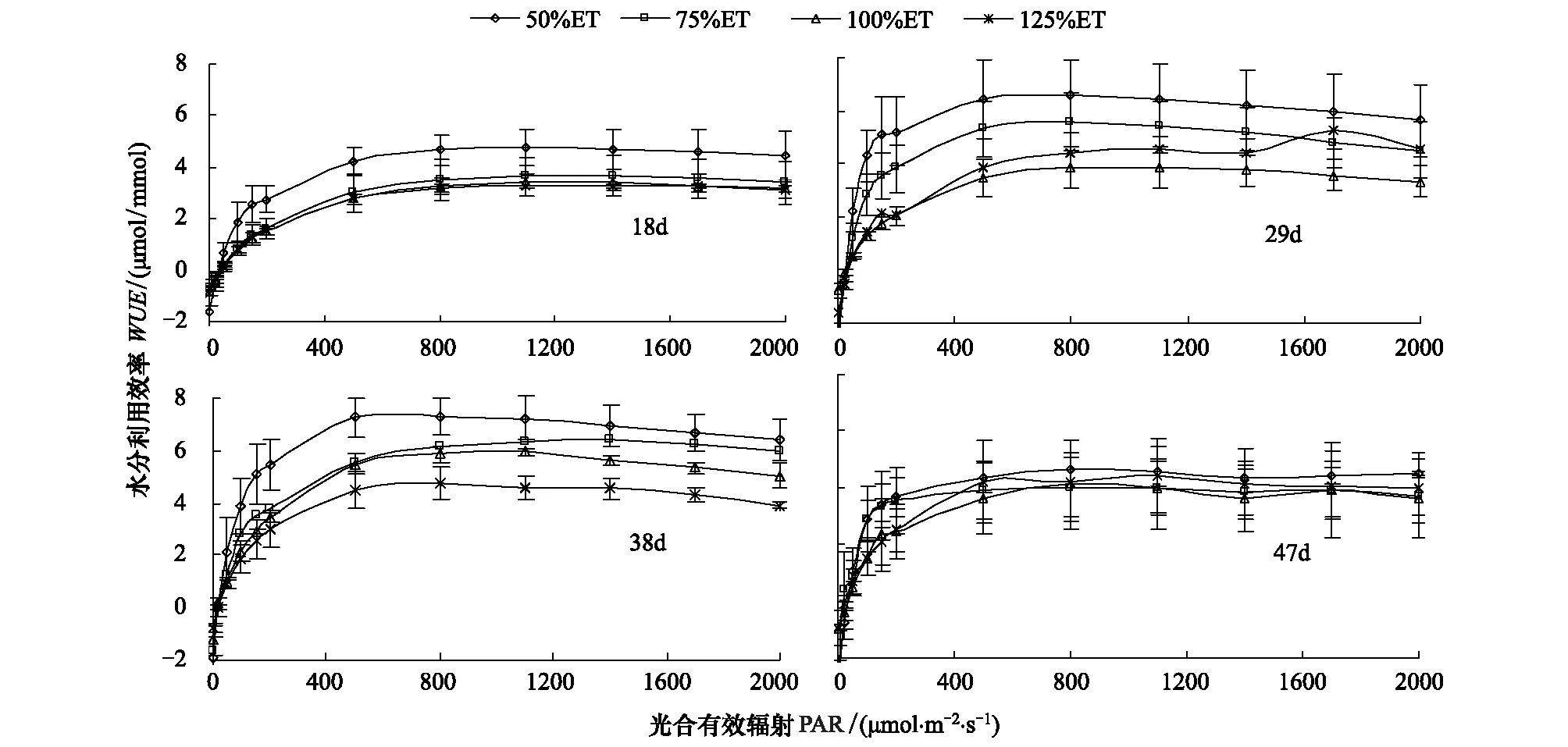
图5 水分对番茄不同叶龄叶片水分利用效率光响应过程的影响Fig.5 Effect of water on light response of water use efficiency for different leaf age tomato leaves
2.2.5水分对番茄不同叶龄叶片胞间CO2浓度的影响
由图6可以看出,不同叶龄叶片在50%ET处理下Ci总小于其他处理。番茄叶片叶龄为18 d时,50%ET处理Ci较其他3个处理低,且其他3个水分处理间的Ci相近;当番茄叶片叶龄增加到29 d时,100%ET和125%ET处理Ci变化不大,50%ET和75%ET处理Ci均减小,且各水分处理间差异明显;当叶龄达到38 d时,50%ET、75%ET和125%ET处理Ci又轻微上升。当叶龄从38 d增加到47 d时,各水分处理Ci变化不大。总体来说,番茄叶片Ci在叶龄从18 d到47 d的过程中,50%E和75%ET处理Ci先降低后升高,其他2个处理变化不明显。

图6 水分对番茄不同叶龄叶片胞间CO2浓度光响应过程的影响Fig.6 Effect of water on light response of intercellular CO2 concentration for different leaf age tomato leaves
2.2.6 水分对番茄不同叶龄叶片气孔限制值的影响
各水分处理Ls随PAR的变化趋势与WUE随PAR的变化趋势相似(图7)。在PARlt;200 μmol·m-2·s-1时,各处理Ls呈直线上升,其中50%ET处理,Ls增幅最大;PAR在200—500 μmol·m-2·s-1时,各水分处理Ls缓慢增大;当PARgt;500 μmol·m-2·s-1时,各水分处理Ls趋于平缓,不再增大。不同叶龄叶片的Ls均以50%ET处理的最高。总之,50%ET和75%ET处理Ls随叶龄增大先升高后降低,在叶龄为29 d时均最大;100%ET和125%ET处理Ls随叶龄增大,Ls略有升高。在叶龄为47 d时,各水分处理Ls相差不大。
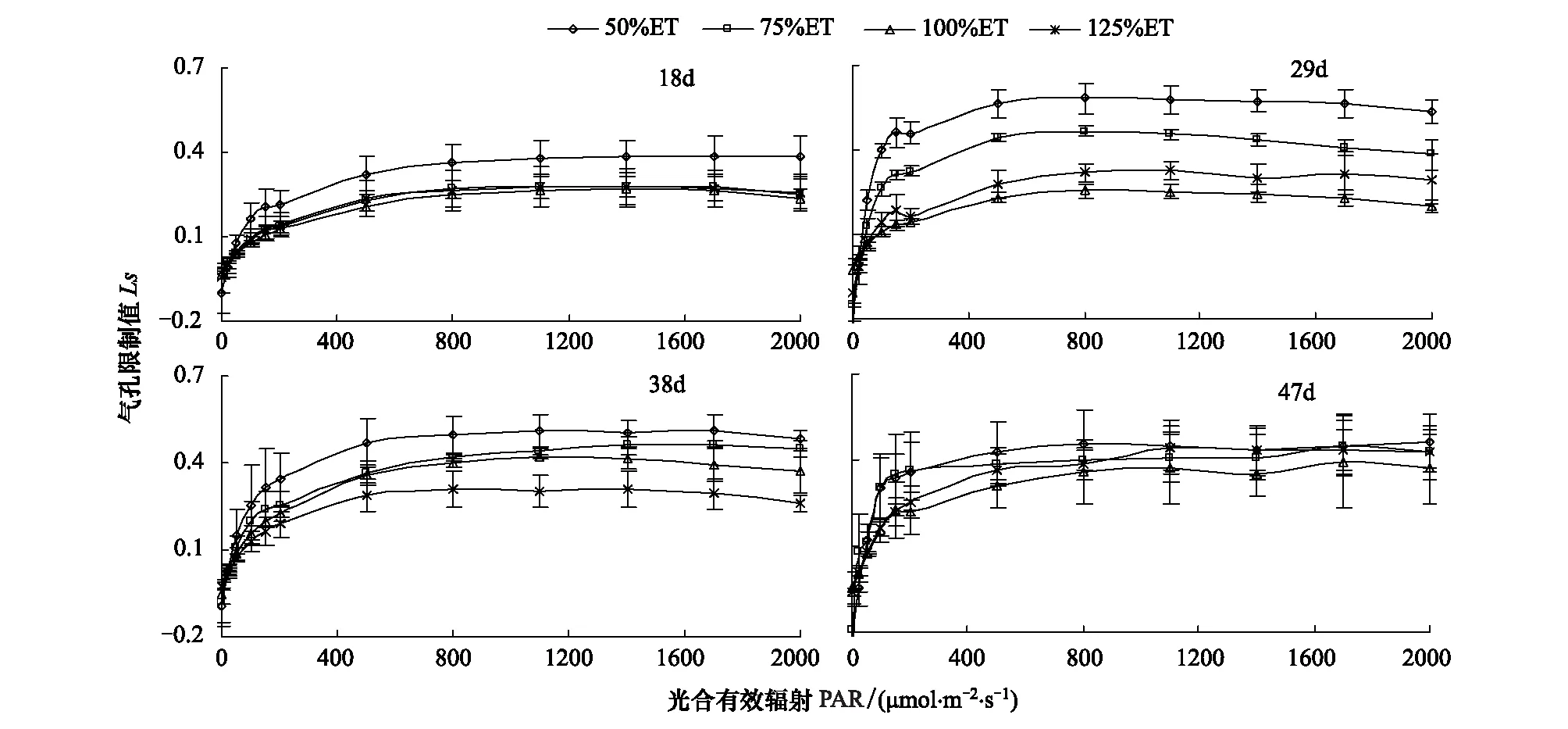
图7 水分对番茄不同叶龄叶片气孔限制值光响应过程的影响Fig.7 Effect of water on light response of stomatal limitation value for different leaf age tomato leaves
2.3 水分对番茄不同叶龄叶片光响应特征参数的影响
最大光合速率(Pmax)、表观量子效率(α)、暗呼吸速率((Rd)、光补偿点(LCP)以及光饱和点(LSP)是由光响应曲线所适合的光合模型计算得到。水分和叶龄对结果期番茄叶片的光响应特征参数有明显影响(表1)。Pmax是度量叶片光合能力的指标。本研究中,Pmax随叶龄的增大而降低。当叶片叶龄为18 d 时,Pmax随灌溉量的增加先升高而降低,以75%ET处理最高(26.73 μmol·m-2·s-1),50%ET处理最低(20.64 μmol·m-2·s-1),与75%ET处理差异显著;随着叶龄增加,番茄叶片在叶龄为29 d时随灌溉量的增加先增大而减小,Pmax最高值(24.24 μmol·m-2·s-1)出现在100%ET处理,与75%ET(16.31μmol·m-2·s-1)和50%ET(11.00 μmol·m-2·s-1)达到显著性差异。当番茄叶片叶龄进一步增加到38 d,除50%ET处理Pmax出现轻微的增加外,其他处理的均进一步减小。此时,番茄叶片的Pmax以125%ET处理达到最大值(18.18 μmol·m-2·s-1)。当番茄叶片叶龄为47 d 时,各水分处理Pmax继续减小,但随灌溉量增加而增加,以125%ET处理为最大值(18.17 μmol·m-2·s-1)。随着叶龄的增加,50%ET和75%ET处理的Pmax在叶龄增大初期(18—29 d)过程中下降较快,各处理在后期(38—47 d)下降均比较缓慢。
α是反映了作物在弱光下对光照的利用效率。从表1可以看出,水分对不同叶龄番茄叶片的α影响不明显。50%ET和75%ET处理α在叶龄从18 d 到29 d 的过程中呈现出降低的趋势,在以后叶龄增加的过程中α先在叶片叶龄为38 d 时达到最大值,然后降低。100%ET和125%ET处理随叶龄的增加总体表现为先增加后降低的趋势,并且也在叶片叶龄为38 d 时达到最大值。水分和叶龄对Rd和LCP影响无规律。LSP随水分和叶龄的变化与Pmax随之的变化极为相似,这可能是LSP与Pmax有密切关系。番茄叶片的LSP随叶龄的增大而降低。当叶片叶龄为18 d 和 29 d时,LSP随灌溉量的增加先升高后降低,均在100%ET时最大(分别为2317.12 μmol·m-2·s-1和1957.53 μmol·m-2·s-1),当叶龄为38 d 和47 d 时,LSP随灌溉量的增加而增加,并都在125%ET处理达到最高值(分别为1375.39 μmol·m-2·s-1和1333.56 μmol·m-2·s-1)。随着叶龄的增加,各水分处理的LSP在叶龄为18—38 d期间下降较快,在38—47 d期间变化较小。

表1 水分对番茄不同叶龄叶片光响应特征参数的影响
同一叶龄同列数据后标不同小写字母者表示差异显著(Plt;0.05)
3 结论与讨论
水是植物进行光合作用所必不可少的,除了水是进行光合作用的原料外,水分在维持叶片水势、调节气孔等方面起着更重要的作用。许多关于番茄的研究[12- 18]表明,水分直接影响番茄在光合作用过程中的气体交换参数。土壤水分降低使番茄的Pn、Gs、Tr、Ci降低,WUE增加。本研究中,番茄叶片Pn、Gs、Tr、Ci随灌溉量的降低而降低,WUE却提高,与前人研究一致[12- 18]。根据Farqhar 和 Sharkey[21]的理论,只有当Ci降低和Ls增大时,Pn的降低主要是由气孔导度的降低引起,限制因素为气孔因素。反之,如果Pn的下降伴随着Ci的提高,光合作用的主要限制因素则是非气孔因素[1]。本研究中,随着灌溉量的减少,番茄叶片的Ci均降低,并伴随着Ls的升高,这表明番茄叶片在低灌溉量下Pn降低主要是由气孔限制因素引起的。水分胁迫使叶片气孔关闭,Gs降低导致进入叶片内的CO2浓度减少,光合作用的供应功能降低(Ci减少),光合作用的底物变少使植株净光合速率降低[22]。
有研究表明Gs是引起Pn随叶龄变化的一个因子,Pn和Gs随叶龄增大同步下降[23- 24]。Kitajima等[25]认为Pn与叶龄呈负相关关系。Field和Mooney[26]以及霍宏和王传宽[27]认为光合能力和Gs随着叶龄的增加而下降,WUE不受叶龄变化的影响。本研究中,番茄叶片Pn和Gs随着叶龄的增大而同步减小,与前人研究一致。除125%ET处理WUE随叶龄的变化不大,其他3个处理WUE均随叶龄的增大而呈现增加后降低的趋势。
Peri等[28]认为Pn和Gs先迅速升高并在叶片完全展开后达到最大值,然后随着叶片的衰老而逐渐降低。艾希珍等[29]和王玲平等[30]也认为叶片叶龄较小时,气孔与非气孔因素同时影响叶片的Pn;而叶片衰老时,叶肉因素即非气孔限制因素影响更多。本研究中,50%ET和75%ET处理下,叶片在叶龄从18 d到29 d的过程中,Ci降低,Ls升高;说明此阶段气孔限制因素引起Pn下降;当叶龄为29—47 d,Ci逐渐增大,Ls降低;说明此过程中非气孔限制因素引起Pn的下降;这表明在50%ET和75%ET处理下,气孔限制与非气孔限制因素是番茄叶片Pn随叶龄变化的原因。100%ET和125%ET处理Ci随叶龄变化不大,Ls略有升高,引起番茄叶片Pn随叶龄变化的原因还有待进一步研究。
Pmax、α、Rd和LSP及LCP是表征植物光合作用的重要特征参数。大量研究表明植物光响应特征参数对水分的响应有明显的阈值,过高或过低的土壤水分都会使Pmax和α及LSP减小[31- 38]。本研究中,番茄不同叶龄叶片Pmax最适的补充灌溉量不同,且对水分的响应有明显的阈值。叶龄为18 d 的番茄叶片Pmax在75%ET处理下最高,叶龄为29 d的番茄叶片Pmax在100%ET处理下最高,叶龄为38 d和47 d的番茄叶片Pmax在125%ET处理下最高;在叶龄为18 d 时Pmax为20.64—26.73 μmol·m-2·s-1;叶龄为29 d 时,Pmax为11.00—24.24 μmol·m-2·s-1;叶龄为38 d 时,Pmax为11.77—18.18 μmol·m-2·s-1;叶龄为47 d时,Pmax为9.09—18.17 μmol·m-2·s-1。本研究表明,叶龄的增大会降低番茄叶片的Pmax,这与Peat[8]在番茄、霍宏和王传宽[28]在红松、张小全和徐德应[39]在杉木上的研究一致。另一方面,水分胁迫会加速番茄叶片Pmax的降低。因此,随着叶片叶龄的增大,较高补充灌溉量有利于番茄叶片光合作用的进行。
Field和Mooney[26]认为α并不因叶龄的变大而受到影响。艾希珍等[29]认为黄瓜叶片、王玲平等[30]认为萝卜叶片的α均随叶龄的增大先升高后降低,最大值分别出现在叶片伸出后的 20—40 d 和25 d。本研究中,α随叶龄的增加先升高后降低,各水分处理下均在叶龄为38 d时达到最大值。不同水分处理下番茄叶片叶龄为18 d 时α为0.0518—0.0556;叶龄为29 d 时,α为0.0522—0.0594;叶龄为38 d 时,α为0.0619—0.0693;叶龄为47 d时;α为0.0538—0.0606。
植物LSP较高表明对强光的利用率也较高。本研究中不同叶龄番茄叶片在125%ET处理下LSP较高,而在较低的灌溉量(50%ET)条件下,LSP总是表现为最低。不同水分处理下番茄叶片的LSP随叶龄的加大而减小。
总之,不同叶龄叶片进行光合作用时所需的适宜的基质含水量不同。叶龄较小,Pn在基质含水量较低时,就可达到极大值;当叶龄逐渐增大,要获得较大的Pn就必须提高基质含水量。本研究中,进一步提高补充灌溉量是否可以提高番茄植株的光合能力还有待进一步研究。
[1] Xu D Q. Photosynthesis Efficiency. Shanghai: Shanghai Science and Technology Press, 2002.
[2] Wei A L, Wang Z M, Zhai Z M, Gong Y S. Effect of soil drought on C4 photosynthesis enzyme activities of flag leaf and ear in wheat. Scientia Agricultura Sinica, 2003, 36(5): 508- 512.
[3] Jeyaramraja P R, Meenakshi S N, Joshi S D. Water deficit induced oxidative damage in tea (Camelliasinensis) plants. Plant Physiol, 2005, 162: 413- 419.
[4] Winter K, Schramm M J. Analysis of stomatal and non-stomatal components in the environmental control of CO2exchanges in leaves of Welwitschia mirabilis. Plant Physiol, 1986, 82(1): 173- 178.
[5] Yu G R, Wang Q F. Eco-physiology of plant photosynthesis, transpiration, and water use. Beijing: Science and Technology Press, 2010: 190, 191.
[6] Johnson and Thornley. I R Johnson, J H M Thornley. Vegetative crop growth model incorporating leaf area expansion and senescence, and applied to grass. Plant, Cell and Environment, 1983, 6: 721- 729.
[7] Herrick J D, Thomas R B. Effects of CO2enrichment on the photosynthetic light response of sun and shade leaves of canopy sweetgum trees (Liguidambarstyraciflua) in a forest ecosystem. Tree physiology, 1999, 19: 779- 786.
[8] Peat W E. Relationships between photosynthesis and light intensity in the tomato. Annals of Botany, 1970, 34: 319- 328.
[9] Bassman J B, Zwier J C. Gas exchange characteristics ofPopulustrichocarpa,PopulusdeltoidesandPopulustrichocarpa×Populusdeltoidesclone. Tree Physiology, 1991, 8: 145- 149.
[10] Ye Z P. A new model of light-response of photosynthesis and its application. Journal of Biomathematics, 2008, 23(4): 710- 716.
[11] Shi W M, Chen Q Y, Qiao X J. Preliminary study on the model for the photosynthesis rate of cucumber leaves at different ages and determination of parameters in sunlight greenhouse. Transactions of the Chinese Society of Agricultural Engineering, 2005, 21(5): 113- 118.
[12] Guo Y B, Feng H, Wu P T. Effects of water stress on physiological characteristic and water production efficiency of tomato. Journa1 of Irrigation and Drainage, 2008,27(3): 52- 55.
[13] Zou T S. The study of photosynthetic and transpiration daily variation of tomato leaf under different soil moisture condition. China Vegetable, 1989, 1(6): 8- 9,15.
[14] Chen J P, Liu Z G, Duan A W, Meng Z J, Zhang J Y. Effects of soil moisture on physiological characteristics and the dynamic state of factors causing photosynthesis decline in potted tomato leaves in greenhouse, Acta Botanica Boreali-occidentalia Sinica, 2004, 24(9): 1589- 1593.
[15] Wang L, Ren S M, Bi Y G, Liu H L, Wu W Y. Effects of soil water and organic manure to the photosynthesis and chlorophyll of tomato in the greenhouse. Journal of Irrigation and Drainage, 2004, 23(2): 66- 68, 80.
[16] Wang X W, Fu Q S, Wang Y J, Zhang J H, Lu H, Guo Y D. Effects of water stress on growth and photosynthetic system characters ofLycopersiconesculentumL. Journal of China Agricultural University, 2010, 15(1):7- 13.
[17] Jiang F C Shi L, Li Y X, Liu M Y. Effects of water and Zinc on leaf potential, photosynthetic characteristics and transpiration efficiency of tomato. Journal of Irrigation and Drainage, 2009, 28(6): 125- 127.
[18] Gao F S, Xu K, Wang L, Su H, Xu Y G. Effects of soil water content on H2O and CO2exchange in tomato leaves in different seasons. Chinese Journal of Applied Ecology, 2007, 18(2): 371- 375.
[19] Li J M, Wang P, Li J. Effect of irrigation amount on physiology, biochemistry and fruit quality of greenhouse tomato under sub-low temperatures. Transactions of the Chinese Society of Agricultural Engineering, 2010, 26(2): 129- 134.
[20] Mills T M., Li J M, Hossein M H. Physiological response of gold kiwifruit (Actinidiachinensis) to reduced irrigation. Journal of America Soc Hort Sci, 2009, 134(6): 667- 683.
[21] Bassman J B, Zwier J C. Gas exchange characteristics ofPopulustrichocarpa,PopulusdeltoidesandPopulustrichocarpa×Populusdeltoidesclone. Tree Physiology, 1991, 8: 145- 149.
[22] Farquhar G D, Sharkey T D. Stomatal conductance and photosynthesis. Ann Rev Plant Physiol, 1982, 33: 317- 345.
[23] Lambers H, Chapin F S, Pons T L//Plant physiological ecology. Zhang G P, Zhou W J,trans. Hangzhou: Zhejiang University Press, 2003: 29.
[24] Lin Z F, Ehleringer J. Effects of leaf age on photosynthesis and water use efficiency of papaya. Photosynthetica, 1982, 16: 514- 519.
[25] Xie S and Luo X. Effect of leaf position and age on anatomical structure, photosynthesis, stomatal conductance and transpiration of Asian pear. Botanical Bulletin of Academia Sinica, 2003, 44: 297- 303.
[26] Kitajima K, Mulkey S S, Samaniego M, Wright S J. Decline of photosynthetic capacity with leaf age and position in two tropical pioneer tree species. American Journal of Botany. 2002, 89(12): 1925- 1932.
[27] Field C B, Mooney H A. Leaf age and seasonal effects on light, water and nutrient use efficiency in a California shrub, Oecologia, 1983, 56: 348- 355.
[28] Huo H, Wang C K. Effects of canopy position and leaf age on photosynthesis and transpiration ofPinuskoraiensis. Chinese Journal of Applied Ecology, 2007, 18(6): 1181- 1186.
[29] Peri P L, Arena M, Pastur M G, Lencinas M V. Photosynthetic response to different light intensities, water status and leaf age of twoBerberisspecies (Berberidaceae) of Patagonian steppe, Argentina. Journal of Arid Environments, 2011, 75: 1218- 1222.
[30] Ai X Z, Ma X Z, Yu M, Xing Y X. Changes of photosynthetic characteristics of cucumber leaves during unfolding in solar-greenhouse. Scientia Agricultura Sinica, 2005, 38(3): 558- 564.
[31] Wang L P, Mao W H, Hu T H, Bao C L, Dai D L. The Changes of photosynthetic properties in radish leaves during leaf senescence. Acta Agriculturae Boreali-Sinica, 2007, 22(S1):36- 39.
[32] Chen J, Zhang G C, Zhang S Y, Wang M J. Response processes of Aralia elata photosynthesis and transpiration to light and soil moisture. Chinese Journal of Applied Ecology, 2008, 19(6): 1185- 1190.
[33] Lang Y, Zhang G C, Zhang Z K, Liu S S, Liu D H, Hu X L. Light response of photosynthesis and its simulation in leaves ofPrunussibiricaL.under different soil water conditions. Acta Ecologica Sinica, 2011, 31(16): 4499- 4508.
[34] Zhang S Y, Zhou F Z; Xia J B, Zhang G C. The responses ofeuonymusfortuneivar.radicansSieb. leaf photosynthesis to light in different soil moisture. Acta Botanica Boreali-Occidentalia Sinica, 2007, 7(12): 2514- 2521.
[35] Xia J B, Zhang G C, Liu G, Han W, Chen J, Liu X. Light response ofWisteriasinensisleaves physiological parameters under different soil moisture conditions. Chinese Journal of Applied Ecology, 2007, 18(1): 30- 34.
[36] Zhu Y Y, He K N, Tang D F, Gong Y X. Response to light ofUlmuspumilain different soil moisture. Research of Soil and Water Conservation, 2007, 14(2): 92- 94.
[37] Zhu Y B, Bo L D, Han M M,. Light response of maize leaves physiology parameters under different soil moisture conditions. Agricultural Research in the Arid Areas, 2007, 25(S1): 168- 170.
[38] Han R F, Li J M, Bai R F, Da H G. Effect of water on characteristics of light response of muskmelon leaves. Journal of Northwest A amp; F University (Natural Science Edition), 2011, 39(7): 123- 128.
[39] Li Y X, Shen S H, Li L, Wang X M, Zhang F C, Han X M. Effects of soil moisture on leaf gas exchange and chlorophyll fluorescence parameters of winter wheat during its late growth stage. Chinese Journal of Ecology, 2012, 21(1): 76- 82.
[40] Zhang X Q, Xu D Y. Light responses of 18-year-old China fir shoots in relation to shoot ages and positions within crown. Acta Ecologica Sinica, 2001, 21(3): 409- 414.
参考文献:
[1] 许大全.光合作用效率. 上海: 上海科学技术出版社, 2002, 43,84- 95.
[2] 魏爱丽, 王志敏, 翟志席, 翟志席, 龚元石. 土壤干旱对小麦旗叶和穗器官C4光合酶活性的影响. 中国农业科学, 2003, 36(5): 508- 512.
[5] 于贵瑞, 王秋凤等. 植物光合、蒸腾与水分利用的生理生态学. 北京: 科技出版社, 2010: 190, 191.
[9] 叶子飘. 光合作用对光响应新模型及其应用. 生物数学学报, 2008, 23 (4): 710- 716.
[11] 史为民, 陈青云, 乔晓军.日光温室黄瓜叶片光合速率模型及其参数确定的初步研究. 农业工程学报, 2005, 21(5): 113- 118.
[12] 郭艳波, 冯浩, 吴普特. 水分亏缺对番茄生理特性及水分生产效率的影响. 灌溉排水学报, 2008, 27(3): 52- 55.
[13] 邹冬生. 不同土壤水分条件下番茄叶片光合及蒸腾日变化研究. 中国蔬菜, 1989, 1(6): 8- 9,15.
[14] 陈金平, 刘祖贵, 段爱旺, 孟兆江, 张寄阳. 土壤水分对温室盆栽番茄叶片生理特性的影响及光合下降因子动态. 西北植物学报, 2004, 24(9): 1589- 1593.
[15] 王磊, 任树梅, 毕勇刚, 刘洪禄, 吴文勇.土壤水分及有机肥料对番茄叶片光合特性及叶绿素含量影响的试验研究. 灌溉排水学报, 2004, 23(2): 66- 68, 80.
[16] 王学文, 付秋实, 王玉珏, 张京红, 路河, 郭仰东.水分胁迫对番茄生长及光合系统结构性能的影响.中国农业大学学报, 2010, 15(1): 7- 13.
[17] 姜凤超, 石磊, 李玉欣, 刘孟雨. 水分和锌对番茄叶水势、光合特性及蒸腾效率的影响. 灌溉排水学报, 2009, 28(6): 125- 127.
[18] 高方胜, 徐坤, 王磊, 苏华, 徐立功. 土壤水分对不同季节番茄叶片水和二氧化碳交换特性的影响. 应用生态学报, 2007, 18(2): 371- 375.
[19] 李建明, 王平, 李江.灌溉量对亚低温下温室番茄生理生化与品质的影响. 农业工程学报, 2010, 26(2): 129- 134.
[23] Lambers H, Chapin F S, Pons T L. 植物生理生态学//张国平, 周伟军, 译. 杭州: 浙江大学出版社, 2003: 29.
[28] 霍宏, 王传宽. 冠层部位和叶龄对红松光合蒸腾特性的影响. 应用生态学报. 2007, 18(6): 1181- 1186.
[30] 艾希珍, 马兴庄, 于立明, 邢禹贤.日光温室黄瓜叶片展开过程中光合特性的变化. 中国农业科学, 2005, 38(3): 558- 564.
[31] 王玲平, 毛伟海, 胡天华, 包崇来, 戴丹丽. 萝卜叶片衰老过程中光合性能变化研究. 华北农学报, 22(增刊):36- 39.
[32] 陈建, 张光灿, 张淑勇, 王梦军. 辽东楤木光合和蒸腾作用对光照和土壤水分的响应过程. 应用生态学报, 2008, 19(6): 1185- 1190.
[33] 郎莹, 张光灿, 张征坤, 刘顺生, 刘德虎, 胡小兰. 不同土壤水分下山杏光合作用光响应过程及其模拟. 生态学报, 2011, 31(16): 4499- 4508.
[34] 张淑勇, 周泽福, 夏江宝, 张光灿. 不同土壤水分条件下小叶扶芳藤叶片光合作用对光的响应. 西北植物学报. 2007, 7(12): 2514- 2521.
[35] 夏江宝, 张光灿, 刘刚, 韩炜, 陈建, 刘霞. 不同土壤水分条件下紫藤叶片生理参数的光响应. 应用生态学报, 2007, 18(1): 30- 34.
[36] 朱艳艳, 贺康宁, 唐道锋, 巩玉霞. 不同土壤水分条件下白榆的光响应研究. 水土保持研究, 2007, 14(2): 92- 94.
[37] 朱永波, 张仁和, 卜令铎, 韩苗苗, 薛吉全.不同土壤水分条件下玉米叶片生理参数的光响应. 干旱地区农业研究(增刊), 2007, 25: 168- 170.
[38] 韩瑞锋, 李建明, 白润峰, 达会广, 赵艳艳.水分对甜瓜叶片光响应特性的影响. 西北农林科技大学学报: 自然科学版, 2011, 39(7): 123- 128.
[39] 李永秀, 申双和, 李丽, 汪秀敏, 张富存, 韩小梅 土壤水分对冬小麦生育后期叶片气体交换及叶绿素荧光参数的影响, 生态学杂志, 2012, 21(1): 76- 82.
[40] 张小全, 徐德应. 18年生杉木不同部位和叶龄针叶光响应研究. 生态学报, 2001, 21(3): 409- 414.
Effectsofwateronphotosynthesisindifferentageoftomatoleaves
CHEN Kaili, LI Jianming*, HE Huiqiang, HU Xiaohui, YAO Yongzhe, SUN Sanjie
CollegeofHorticulture,NorthwestAgricultureandForestryUniversity,YanglingShaanxi712100,China
Water deficit is one of the major rate-limiting factors for plants growing in arid and semiarid region and directly reduce the plant′s photosynthetic rate. Tomato (LycopersiconesculentumMill.) is one of the most valuable commercial vegetables all over the world and widely cultivated in China whether in open field or in greenhouse. Light-responsive photosynthetic model provides the reference to evaluate and simulate the plant photosynthesis production and the production capacity of the ecological system. The maximum net photosynthetic rate(Pmax) and apparent quantum yield(α) derived from light response curve are the vital parameters of light-responsive photosynthetic model, which are the critical physiology-ecology parameters of plant photosynthesis characteristics and vary with the plants species, varieties, growth condition and their metabolic status etc.. The leaf age directly affects the efficiency of plants photosynthesis and the values of light-responsive photosynthetic parameters. Water deficit could accelerate leaf senescence and shorten the leaf lifespan. Even the photosynthetic performance of leaves under different water conditions shows the obvious difference at the same leaf age. Up to now there are a few pieces of works reported on light-responsive photosynthetic characteristic of photosynthesis in different age of tomato leaves under multiple supplementary irrigation level. The objective of this study is to profile light-responsive photosynthetic characteristics in tomato with different leaf age of leaves under different levels of supplementary irrigation amount, which will provide the light-responsive photosynthetic parameters for photosynthetic model under different water condition and leaf age. In this study, tomato plants potted in greenhouse as material, the gas exchange and light-responsive photosynthetic characteristics parameters of tomato leaves with the change of leaf age under 4 levels of supplemental irrigation amount (50%ET, 75%ET, 100%ET, and 125%ET) were investigated. The results showed that net photosynthetic rate (Pn), stomatal conductance (Gs) and transpiration rate (Tr) all gradually reduced, but water use efficiency (WUE) ascended firstly and then descended with the increase of tomato leaf age; When leaf age were 18 d and 29 d, bothPmaxincreased and then decreased with the increase of supplementary irrigation amount, peaking at 75%ET and 100%ET, respectively. When leaves age were 38 d and 47 d, with the rising of supplementary irrigation amount, bothPmaxascended, peaking at 125%ET treatment. As forα, it rose with leaf aging, reaching a peak at the leaf age of 38 d. And for light saturation point (LSP), it decreased with leaf aging. The light-responsive photosynthetic parameters of different leaf age of tomato leaves under different supplementary irrigation amount were as follows. When leaf age was 18 d,Pmaxwas 20.64—26.73 μmol·m-2·s-1,αwas 0.0518—0.0556; when leaf age was 29 d,Pmaxwas 11.00—24.24 μmol·m-2·s-1;αwas 0.0522—0.0594; when leaf age was 38 d,Pmaxwas 11.77—18.18 μmol·m-2·s-1,αwas 0.0619—0.0693; when leaf age was 47 d,Pmaxwas 9.09—18.17 μmol·m-2·s-1,αwas 0.0538—0.0606. In conclusion, with leaf aging, increasing supplementary irrigation amount could delay the fall of photosynthetic capacity of tomato leaves. Stomatal limiting factor was the main factor of influencing photosynthesis of tomato leaves; Stomatal limiting and non-stomatal limiting factors were the causes of photosynthetic rate varying of tomato leaves according to the leaf age.
tomato; leaf age; water; light response characteristics; gas exchange parameter
国家“十二五”科技支撑计划项目(2011BAD12B03-3)
2012- 05- 17;
2013- 04- 15
*通讯作者Corresponding author.E-mail: lijianming66@163. com
10.5846/stxb201205170736
陈凯利,李建明,贺会强,胡晓辉,姚勇哲,孙三杰.水分对番茄不同叶龄叶片光合作用的影响.生态学报,2013,33(16):4919- 4929.
Chen K L, Li J M, He H Q, Hu X H, Yao Y Z, Sun S J.Effects of water on photosynthesis in different age of tomato leaves.Acta Ecologica Sinica,2013,33(16):4919- 4929.
猜你喜欢
科学(2022年4期)2022-10-25
中国特种设备安全(2021年5期)2021-11-06
装备制造技术(2021年4期)2021-08-05
种业导刊(2019年9期)2019-10-21
江苏农业科学(2018年4期)2018-03-27
制造技术与机床(2017年11期)2017-12-18
农民致富之友(2017年7期)2017-04-27
农民致富之友(2017年2期)2017-03-23
Coco薇(2016年5期)2016-06-03
考试周刊(2016年6期)2016-03-11
