
含有Th细胞表位的合成PCV2 ORF2 Cap多肽抗原的表达及免疫原性分析
2015-03-23 07:56陈善真李中圣陈克宏王贵平李其昌
畜牧兽医学报 2015年12期
陈善真,赵 焱,李中圣,罗 均,陈克宏,王贵平,李其昌
(广东海大畜牧兽医研究院有限公司,广州 511400)
含有Th细胞表位的合成PCV2 ORF2 Cap多肽抗原的表达及免疫原性分析
陈善真,赵 焱,李中圣,罗 均,陈克宏,王贵平,李其昌*
(广东海大畜牧兽医研究院有限公司,广州 511400)
拟测定在PCV2 ORF2 Cap P1多肽抗原中添加从PCV2 ORF1、ORF3中筛选的Th细胞表位多肽在促进其免疫原性中的作用,以及合成表达的PCV2 ORF2 Cap P1多肽抗原作为疫苗抗原的可行性。采用生物信息学及分子生物学方法,筛选获得1条泛宿主新型Th细胞表位多肽和3条PCV2特异的含有Th细胞抗原表位的多肽序列,然后将这4条Th细胞多肽氨基酸序列与筛选自ORF2 Cap1上的1条B细胞表位多肽序列进行串联组合,密码子优化,插入酶切位点、终止密码子,然后进行密码子适应指数和分布频率分析,预测可以实现高效表达后再进行化学合成。合成的P1多肽抗原连接到pET-30a表达载体,并转化至BL21(DE3)pLysS感受态细胞中,构建表达工程菌BL21(DE3)pLysS-pET-30a-P1。经IPTG诱导表达后P1可实现高效表达,SDS-PAGE鉴定表达量,Western Blot鉴定其生物活性,然后分别免疫小鼠和猪,测定小鼠免疫后的抗体水平以及猪免疫后P1合成多肽抗原对外周血淋巴细胞的刺激增殖情况。结果表明,设计合成表达的PCV2 P1多肽抗原序列,密码子适应性指数为0.89,能够实现高水平表达,目的片段大小约为28.29 ku,具有良好的免疫原性;MTT试验结果表明,P1多肽抗原免疫后机体内IFN-γ和IL-4的表达量明显增加,且与对照组差异显著(P<0.05)。这说明P1能够诱导机体产生细胞免疫和体液免疫反应,为其作为疫苗用抗原的进一步研究奠定了理论基础。
PCV2;Th细胞表位多肽;Cap蛋白
猪圆环病毒2型(PCV2)是引起断奶仔猪多系统衰竭综合征(PWMS)的重要病原,感染PCV2的猪可导致淋巴系统损伤和免疫缺陷,这可能是由于PWMS感染后损伤猪的T、B淋巴细胞,使单核/巨噬细胞系的细胞数目增多,并改变细胞因子的应答模式,从而引起机体抵抗力下降[1-3]。
PCV2能损伤机体细胞因子的免疫应答,患有PMWS的猪主要特征为淋巴细胞缺损和组织细胞浸润[4]。从PCV2感染和患病猪中分离出来的外周血单核细胞,其分泌IFN-γ和记忆性T细胞的能力下降[5],而从健康和患PMWS猪中分离到的PBMS用PHA和SEB刺激后,IL-2、IL-4的产量也会降低[6]。
淋巴细胞增殖试验表明,免疫优势辅助T细胞反应表位始终定位于ORF1和ORF3 非结构蛋白上[7]。没有免疫优势的线性Th表位定位于ORF2编码的非结构蛋白上。这就可以解释传统的以PCV2衣壳蛋白为抗原而制备的亚单位疫苗因其Th细胞表位的稀缺而导致免疫原性的有限性。辅助性T细胞表位可以诱导保护性T辅助细胞(Th)的反应,从而发信号给B细胞产生抗体。因此,以合成肽作为将免疫优势Th表位呈现给免疫系统是影响合成肽免疫原性一个关键因素。
Cap蛋白是PCV2 ORF2编码的结构蛋白,具有良好的免疫原性,免疫动物后能产生较好的中和抗体,从而抵御病毒感染对B细胞的损伤[8]。猪用疫苗的免疫功能,及其预防猪感染病毒的能力受多个病毒的辅助Th细胞位点的影响,本研究中设计的合成肽疫苗,与PCV2衣壳蛋白上筛选定位的B细胞表位相比,除具有免疫优势B细胞表位外,还增加了通用型和PCV2特异的Th细胞表位,辅助Th细胞位点与Cap蛋白B 细胞位点的交叉保护可大大提高疫苗的效力。
鉴于疫苗对防制此病的意义[9],本研究选取ORF2的主要B细胞抗原表位,在此基础上设计增加泛宿主新型辅助T细胞表位多肽序列,组合后进行密码子优化,增加其抗原性及表达量,然后借助大肠杆菌表达系统实现其表达,并通过小鼠和断奶仔猪的免疫试验,检测抗体水平和刺激淋巴细胞增殖的情况来测定合成表达的PCV2合成肽抗原的免疫原性,为下一步的疫苗相关研究奠定基础。
1 材料与方法
1.1 实验材料
BL21(DE3)pLysS感受态细胞、蛋白质Marker,预染Marker,购自TIANGEN生物;限制性内切酶NdeⅠ和XhoⅠ,购自TaRaKa生物;IPTG、卡那霉素、肝素钠、淋巴细胞分离液、MTT、Gibco RPMI-1640培养液及其他常规试剂、培养基购自北京鼎国生物技术有限责任公司;ISA201VG佐剂,由SEPPIC公司提供;勃林格PCV2商品苗,氨基酸序列由南京金斯瑞生物科技有限公司进行合成。
1.2 实验动物及分组
BALB/c小鼠购自南方医科大学实验动物中心,试验用断奶仔猪,12头,购于鹤山市某猪场,均未进行PCV2疫苗免疫,随机分为4组,每组3头:P1抗原免疫组、勃林格疫苗免疫组、空白佐剂免疫组和空白对照组。
1.3 重组多肽序列P1的设计
参考相关的研究[7,10],设计筛选并合成含有泛宿主新型辅助T细胞表位的4条多肽,筛选部位来自于PCV2 ORF1、ORF3,并参考其他学者的研究[11-12]从PCV2 ORF2中筛选出1条主要的B细胞多肽抗原表位,然后利用连接性碱基将这5条多肽进行串联,增加相应的连接性氨基酸及酶切位点,对其中的密码子进行优化,并测定密码子适应指数及密码子选择频率。将优化设计完成的氨基酸序列命名为P1,P1序列含有256个氨基酸,理论相对分子质量为28.29 ku。
1.4 表达质粒的构建
表达序列设计完成后送由南京金斯瑞生物科技有限公司进行合成,并将序列经NdeⅠ和XhoⅠ双酶切后,连接到pET-30a表达载体中,构建表达质粒,构建完成的表达质粒命名为pET-30a-P1。
1.5 P1的诱导表达
将构建完成的表达质粒连接转化至BL21(DE3)pLysS感受态细胞中,构建表达工程菌BL21(DE3) pLysS-pET-30a-P1。挑取单个菌落在含有0.05%卡那霉素的LB液体培养基中37 ℃振荡培养,待OD600 nm值达到0.6 时加入IPTG进行诱导表达,使其终浓度为1.0 mmol·L-1,诱导表达6 h后收集菌液,进行SDS-PAGE电泳检测。
1.6 P1免疫原性的Western Blot分析
应用Western Blot验证表达多肽P1的免疫原性,方法如下:①以表达的P1为样品,进行聚丙烯酰胺凝胶电泳;②转膜,封闭;③以自制纯化的PCV2猪多克隆抗体为一抗,1∶500倍稀释后进行孵育(37 ℃,1 h),清洗;④加1 000倍稀释的HRP-羊抗猪IgG二抗,孵育(37 ℃ 1 h),清洗;⑤应用DAB 显色试剂盒进行显色。
1.7 小鼠免疫试验验证P1免疫原性
将表达纯化的P1按照5 μg·mL-1的浓度分别免疫8~10 g的BALB/c小鼠,一免两周后进行采血并二免,二免剂量同第一次免疫剂量。分别在免疫前、一免后2周、二免后2、3、5周采血,通过ELISA方法检测血清抗体水平,验证P1的免疫原性。
1.8 MTT试验测定P1合成表达多肽对外周淋巴细胞的增殖刺激作用
将P1与ISA 201VG佐剂按说明书比例混匀后,免疫断奶仔猪,2 mL·头-1,佐剂组仅免疫佐剂,2 mL·头-1,勃林格疫苗免疫组,1 mL·头-1,空白对照组免疫灭菌生理盐水,2 mL·头-1。免疫5周后分别无菌采集4个组的颈静脉外周抗凝血,10 mL·头-1,收集猪血清后,超净台内分离淋巴细胞,用台盼蓝染色计数活细胞数(>95%),然后用RPMI-1640培养液调整细胞浓度为5×106mL-1,制成单细胞悬液。
将淋巴细胞悬液加入48孔细胞培养板中,加入PCV2 P1表达多肽纯化抗原10 μL(终质量浓度为10 μg·孔-1),每个样品平行培养三孔,同时用其他组作为对照,培养体系为每孔1 mL。置5% CO2,37 ℃培养48 h。在培养结束前4 h,每孔取500 μL细胞上清液,-20 ℃保存待检,余下每孔加入25 μL MTT(初始质量浓度为5 mg·mL-1),继续培养。培养结束后,每孔吹打混匀,然后每孔取出200 μL依次加入96孔酶标板中,酶标板中再依次加入100 μL DMSO,置摇床上低速振荡 10 min,待结晶物充分溶解后,在酶联免疫检测仪测各孔的OD值(570 nm处)。
1.9 MTT试验中T细胞分泌的IFN-γ和IL-4的检测
取上述MTT试验中,37 ℃培养44 h取样的细胞上清液,分别用IFN-γ和IL-4的试剂盒检测外周血淋巴细胞中两种细胞因子的含量,比较不同试验组细胞上清液中IFN-γ和IL-4的分泌表达情况。
2 结 果
2.1 P1多肽序列的筛选及优化
将筛选的1条含有泛宿主新型Th细胞表位多肽和从PCV2 ORF1、ORF3中筛选的3条Th细胞表位多肽(图1)与从ORF2中筛选的1条含有多个B细胞抗原表位的多肽序列(图2),通过衔接性密码子进行串联,设计插入酶切位点和起始终止密码子,并对组成的碱基序列进行优化,优化完成的含有Th细胞表位、B细胞表位以及酶切位点的P1氨基酸序列见图3。优化完成的碱基序列大小为768 bp,预测可表达256个氨基酸。
2.2 优化重组后P1多肽序列的密码子适应指数分析
作者将序列设计优化完成后对氨基酸序列的密码子偏好性进行了分析。密码子适应指数(codon adaptation index,CAI),常用于基因表达水平的测量。此值为0~1,越接近1表示基因的表达水平越高,当密码子适应指数为1.0时,生物体基因可以达到最佳的表达水平。图4结果表明密码子适应性指数为0.89,说明重组优化的基因序列可以实现高水平表达。

图1 从PCV2 ORF1、ORF3中筛选的含有辅助T细胞表位的多肽序列Fig.1 Screening peptide sequences of helper T cell epitopes derived from PCV2 ORF1 and ORF3

图2 ORF2中筛选的含有多个B细胞表位的多肽序列(47-200 aa)Fig.2 Peptide sequences of B cell epitopes derived from PCV2 ORF2(47-200 aa)

图3 优化完成的重组多肽PCV2 P1氨基酸序列(包含酶切位点和终止密码子)Fig.3 The optimized amino acid sequence of the recombinant peptide P1 containing restriction enzyme cutting site and terminator codon

图4 基因序列长度中的密码子利用频率分布(CAI,密码子适应指数)Fig.4 The distribution of codon usage frequency along the length of the gene sequence
2.3 优化重组序列的密码子选择频率分析
最优密码子是指在某物种高表达基因中使用频率最高的密码子,最优密码子使用频率(frequency of optimal codons,FOP)可以反映一段基因序列中密码子在表达过程中的利用情况。由图5可见,横坐标中数值100是指在一个设定的氨基酸序列中密码子在希望的机体或组织表达过程中的利用频率的最高值。而作者设计优化的序列经最优密码子使用频率分析发现,有71%的密码子利用频率在91%~100%,这说明作者优化设计的序列可以实现高水平的表达。
2.4 重组表达质粒的鉴定
将重组后的表达质粒用限制性内切酶NdeⅠ和XhoⅠ进行双酶切,琼脂糖电泳(图6)鉴定发现,目的片段长度与768 bp的理论值相符。
2.5 P1表达后SDS-PAGE鉴定
将构建表达工程菌BL21(DE3)pLysS-pET-30a-P1培养后,用终浓度为1.0 mmol·L-1IPTG进行诱导,诱导表达6 h后收集菌液,进行SDS-PAGE 电泳检测。由鉴定结果图7可见,诱导后有两条蛋白质条带出现,其中一条为大肠杆菌的本底表达,另一条为目的条带,大小约为28.29 ku,且目的条带表达量高于大肠杆菌的本底表达。
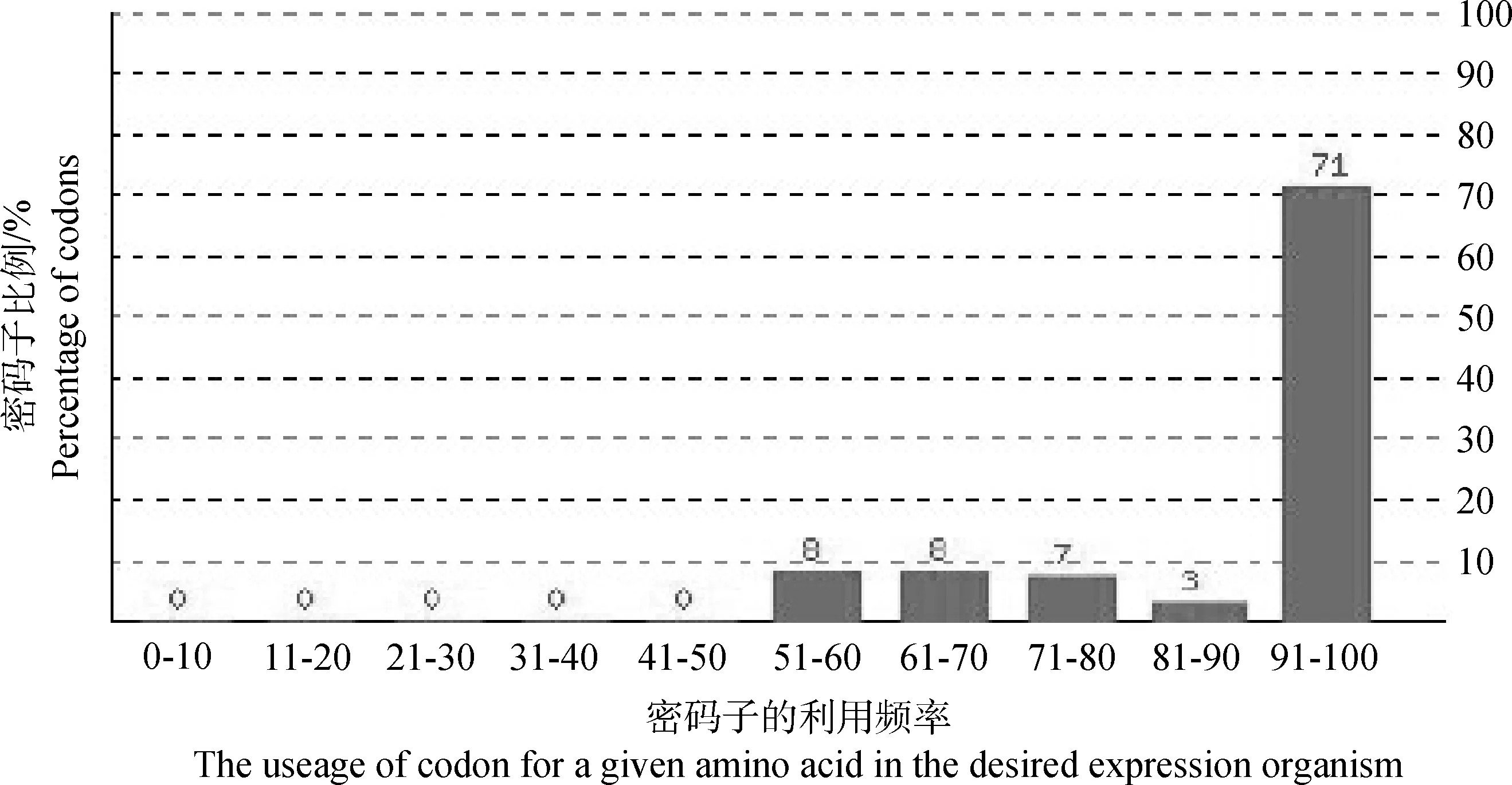
图5 计算机推算的最优密码子的分布频率(FOP)Fig.5 The percentage distribution of codons in computed codon quality groups

1.重组表达质粒;2.酶切后质粒;M.1 kb DNA相对分子质量标准1.5004538-1 plasmid;2.5004538-1 plasmid digested;M.1 kb ladder图6 重组表达质粒的NdeⅠ、XhoⅠ酶切鉴定Fig.6 Identification of recombined PCV2-P1 by double enzyme digestion with NdeⅠ and XhoⅠ
2.6 P1免疫原性的Western Blot分析
将诱导表达的P1多肽抗原进行Western Blot分析,以自制纯化的PCV2猪高免血清抗体为一抗进行检测,结果发现诱导表达的重组多肽抗原可以与PCV2的高免猪血清发生特异性反应(图8),说明表达产物的抗原性良好。
2.7 小鼠免疫P1多肽抗原后抗体水平检测
将表达纯化的P1多肽抗原按1 μg·mL-1包被酶标板,100 μL·孔-1,37 ℃孵育1 h,4 ℃过夜。次日洗涤3次,每次250 μL·孔-1洗涤液,然后加封闭液150 μL·孔-1,37 ℃封闭孵育1 h,同上方法洗涤后,分别对小鼠在免疫前、一免后2周、二免后2、3、5周的血清进行检测,由图9结果可见,免疫后抗体随免疫时间的增加呈上升趋势,说明P1多肽抗原可以引起小鼠机体的免疫反应。2.8 MTT试验测定P1对外周淋巴细胞的增值刺激

M.蛋白质相对分子质量标准;1、3.诱导6 h后样品;2.诱导4 h后样品; 4.诱导前样品M.Protein marker;1,3.Sample after 6 hours’ induction;2.Sample after 4 hours’ induction;4.Sample before induction图7 重组 P1多肽抗原表达后SDS-PAGE鉴定Fig.7 Identification of the expression of recombinant peptide antigen P1 by SDS-PAGE

M.蛋白质预染相对分子质量标准;1.诱导后表达产物M.Prestained protein marker;1.Expression product after induction图8 重组 P1多肽抗原表达后Western Blot分析Fig.8 Western Blot analysis of the expressed recombinant peptide antigen P1
分别用表达纯化的P1多肽抗原对P1疫苗免疫组、空白对照组进行刺激,并设勃林格疫苗免疫组、佐剂免疫组和空白对照无刺激组作为对照,检测P1对猪外周血中淋巴细胞的刺激作用。在免疫65 d后采集外周血,每个样品平行检测3次,具体检测结果见表1。另,刺激指数SI(stimulation index)也可作为判断淋巴细胞增殖滴度的一个重要指标。用表达纯化的P1多肽抗原刺激P1多肽抗原免疫组和空白对照组,并分别设无刺激对照,对SI值进行比较,结果见图10。

图9 P1免疫小鼠后抗体水平检测Fig.9 The antibody level detection of polypeptide antigen P1 in immunized mice
表1 不同免疫组表达多肽P1刺激后淋巴细胞增殖试验结果
Table 1 The results of lymphocyte proliferation stimulated by peptide P1 to different immunity group

组别Group免疫时间/dImmunizationday样品数SamplesOD值ODvalueA、P1免疫+P1刺激P1immunity+P1stimulation6531.2007±0.0119aB、勃林格疫苗免疫+无刺激Boehringervaccineimmunity+nostimulation6530.4123±0.1877cC、空白对照组+P1刺激Black+P1stimulation6530.6780±0.0361bD、佐剂免疫组+无刺激Adjuvantimmunity+nostimulation6530.3460±0.0656dE、空白对照组+无刺激Black+nostimulation6530.3407±0.0674d
同列数据后所标字母相异表示差异显著(P<0.05),所标字母相同表示差异不显著(P>0.05)。下表同
Different letters in the same row means significant difference between the treatments(P<0.05),same letter in the same row means not significant difference between treatments(P>0.05).The same as below
由表1结果可见,A、B、C组与其他各处理试验组之间差异均显著(P<0.05),D组与E组差异不显著(P>0.05),这说明佐剂免疫组与空白对照组无差别,经表达多肽P1刺激后A、C组的淋巴细胞增值情况较B、D、E组均有显著性差异(P<0.05),这说明P1多肽可以有效刺激机体外周血淋巴细胞的增殖;A、C组的差异可以看出同样经P1多肽刺激后,P1多肽免疫组比空白对照组的淋巴细胞增殖能力明显增强(P<0.05),说明P1多肽疫苗有较好的刺激机体产生T细胞免疫反应的能力。
淋巴细胞刺激指数(SI):即试验孔的OD值/对照孔的OD值,可作为反应抗原对淋巴细胞的刺激作用。由图10结果可见,P1多肽对空白组猪血清中淋巴细胞的刺激指数大于2.0,对P1多肽抗原免疫组的刺激指数在3.0以上,说明P1可以有效刺激猪体外周血淋巴细胞的增殖,即可以刺激猪体产生免疫反应。
2.9 T细胞分泌的IFN-γ和IL-4的检测
IFN-γ、IL-4的检测结果见表2。
IFN-γ是由T淋巴细胞中的Th1细胞分泌的,可以参与细胞的免疫应答,因此其含量的检测可以反应机体受到抗原刺激后,细胞免疫应答的水平。多肽P1免疫后的猪外周血液样本再用P1多肽刺激后,IFN-γ含量(表2)与其他试验组和空白对照组相比有明显升高,且差异显著(P<0.05),多肽P1免疫组与勃林格疫苗免疫组IFN-γ的产生水平均高于佐剂对照和空白对照组,这说明疫苗免疫后会刺激猪体内IFN-γ的表达,而PCV2 P1多肽免疫后,猪体内IFN-γ的表达量明显增加,这可能是由于P1多肽序列中既包含有B细胞抗原表位,又含有辅助Th细胞表位,从而可以更好地激活机体的细胞免疫应答,抵抗PCV2病毒的入侵。

图10 P1重组多肽抗原刺激对淋巴细胞刺激指数(SI)的影响Fig.10 The effect of P1 recombinant peptide antigen stimulating to lymphocyte stimulation index(SI)
表2 T细胞分泌IFN-γ、IL-4的检测
Table 2 The level of IFN-γ、IL-4 secreted by T cell

组别Group平行数Parallelnumber平均质量浓度/(pg·mL-1)AverageconcentrationIFN⁃γIL⁃4多肽P1免疫+P1多肽刺激P1immunity+P1stimulation370.4108±1.1221a17.4993±0.7152a多肽P1免疫+不刺激P1immunity+nostimulation327.8327±0.4015c12.1113±0.6794b空白对照组+P1多肽刺激Black+P1stimulation349.6751±1.2854b11.5897±0.5941b勃林格疫苗免疫+不刺激Boehringervaccineimmunity+nostimulation328.0214±0.5349c9.5663±0.1723c佐剂免疫对照组+不刺激Adjuvantimmunity+nostimulation37.4554±0.5072d7.3393±0.3243d空白对照组+不刺激Black+nostimulation36.8706±0.1634d6.7923±0.1009d
IL-4是由Th2细胞分泌的,其对机体免疫应答有重要的作用,可以辅助B细胞产生抗体,因此,其含量的高低可影响机体的体液免疫应答的能力。由表2结果可见,多肽P1免疫组和勃林格疫苗免疫组与空白对照和佐剂对照组差异均显著(P<0.05),这说明两种疫苗免疫后均可使机体内T细胞分泌IL-4的量大大提高;多肽P1免疫+P1刺激组 IL-4的产生水平高于单独疫苗免疫组,同时空白对照组在用P1多肽刺激后T细胞分泌IL-4的能力也明显高于空白对照组,且均差异显著(P<0.05),这说明合成表达的P1多肽与传统疫苗相比,其刺激分泌表达IL-4的量明显增加,可间接的刺激机体的体液免疫应答,这正是由于其序列中包含的Th细胞表位多肽发挥的作用。
3 讨 论
PCV2感染后容易导致免疫抑制,从而引发多重感染和复发感染,造成严重的经济损失,PCV2感染所导致的免疫抑制可能是由于Th细胞受到抑制而造成的,有研究发现,PCV2感染猪的胸腺和扁桃体中的IL-10和IFN-γ mRNA的量均有所增加,而淋巴细胞中IL-2、IL-4、IL-10、IL-12和IFN-γ的mRNA水平则有所下降[6],这些细胞因子含量的异常变化表明PCV2可以导致T细胞免疫抑制。临床预防中主要是采用灭活疫苗和亚单位疫苗免疫。PCV2灭活疫苗存在免疫持续时间短及病毒培养时滴度难以提高而使免疫成本较高的问题,而亚单位疫苗由于Th表位基因的稀缺而存在不能有效激活机体的免疫反应的缺点,因此近年来的研究多倾向于寻找一种有效地可以增强疫苗免疫效果的佐剂或是研发新的表位抗原肽疫苗。
吴胜昔等以固相合成法将3个B细胞抗原表位和1个通用型Th细胞表位串连在一起,利用设计合成的猪圆环病毒2型CAP蛋白新型表位抗原肽免疫小鼠,病毒中和试验测定结果显示,该抗体具有较好的免疫保护作用[13],但该研究中并未进一步测定合成的多肽对机体免疫刺激的影响。本研究中在设计合成的基础上通过对小鼠和猪的免疫实验验证了合成表达的PCV2 Cap P1多肽在刺激机体的细胞免疫和体液免疫方面的作用,并选择代表性的IFN-γ和IL-4的表达量进行检测,结果表明外源Th细胞表位多肽的加入可以刺激机体内该类细胞因子的表达,使其产量明显高于机体正常的表达量,从而可以缓解PCV2感染而造成的Th细胞免疫抑制。
IFN-γ是一种由活化的CD4+、CD8+T淋巴细胞和NK细胞产生的细胞因子,它不仅可以提高CTL细胞和NK细胞的靶细胞的能力,也可以诱导细胞表达IL-2受体,L.Genmei等的研究表明,IFN-γ可以显著增强PCV2疫苗的免疫原性[14]。IL-4是由Th2细胞分泌的细胞因子,具有重要的免疫作用,可以促进B细胞和T细胞的相互作用,辅助B细胞产生抗体,从而促进体液免疫应答[6]。F.Gao等的研究表明,PCV2感染后会引起猪体内IL-4含量的降低[15],因此,在包含刺激这些细胞因子的多肽序列的存在可以明显提高疫苗的保护力,本研究的试验结果也证明了这一点。
4 结 论
一种多肽亚单位抗原其免疫原性的强化,可以通过共价键耦联到有针对性的B细胞表位,即在B细胞表位基础上针对的增加外源而多样的Th细胞表位。本研究在前人研究基础上选取PCV2 Cap蛋白的主要B细胞多肽表位,同时设计加入了4条Th细胞表位多肽,合成表达的PCV2 P1多肽抗原组装成疫苗,疫苗进入机体后可以有效地激活机体的细胞免疫应答和体液免疫应答,从而大大提高了疫苗的保护力,为后续其作为疫苗抗原的进一步研究奠定基础,同时为其他疫病多肽疫苗的研究提供了思路。
[1] FAN H Y,XIAO S B,TONG T Z,et al.Immunogenicity of poricine circovirus type2 capsid protein targeting to different subcellular compartments[J].MolImmunol,2008,45(3):653-660.
[2] HANSEN M S,PORS S E,BILLE-HANSEN V,et al.Occurrence and tissue distribution of porcine circovirus type2 identified by immunohistochemistry in Danish finishing pigs at slaughter[J].JCompPathol,2010,142(2-3):109-121.
[3] 李海超,王 娟,黄秀梅,等.猪圆环病毒2型免疫应答机制研究进展[J].动物医学进展,2014,35(2):105-109. LI H C,WANG J,HUANG X M,et al.Progress on immune response mechanism of porcine circovirus type 2[J].ProgressinVeterinaryMedicine,2014,35(2):105-109.(in Chinese)
[4] FORT M,SIBILA M,NOFRARAS M,et al.Porcine circovirus type 2(PCV2) Cap and Rep proteins are involved in the development of cell-mediated immunity upon PCV2 infection[J].VetImmunolImmunopathol,2010,137(3-4):226-234.
[5] FERRARI L,BORGHETTI P,DE ANGELIS E,et al.Memory T cell proliferative responses and IFN-γ productivity sustain long-lasting efficacy of a Cap-based PCV2 vaccine upon PCV2 natural infection and associated disease[J].VetRes,2014,45:44.
[6] DARWICH L,BALASCH M,PLANA-DURN J,et al.Cytokine profiles of peripheral blood mononuclear cells from pigs with postweaning multisystemic wasting syndrome in response to mitogen,superantigen or recall viral antigens[J].JGenVirol,2003,84(Pt 12):3453-3457.
[7] STEVENSON L S,GILPIN D F,DOUGLAS A,et al.T lymphocyte epitope mapping of porcine circovirus type 2[J].ViralImmunol,2007,20(3):389-398.
[8] 吴胜昔,曾 政,蔡家利,等.猪圆环病毒2型CAP蛋白B细胞表位预测与分析[J].中国兽医杂志,2014,50(1):3-6. WU S X,ZENG Z,CAI J L,et al.Prediction and analysis of the B cell epitopes for capsid protein of porcine circovirus type 2[J].ChineseJournalofVeterinaryMedicine,2014,50(1):3-6.(in Chinese)
[9] 王一平,郭龙军,唐青海,等.猪圆环病毒2型疫苗的研究进展[J].畜牧兽医学报,2012,43(9):1337-1345. WANG Y P,GUO L J,TANG Q H,et al.Research progress on vaccines of porcine cirvirus type 2[J].ActaVeterinariaetZootechnicaSinica,2012,43(9):1337-1345.(in Chinese)
[10] MAHÉ D,BLANCHARD P,TRUONG C,et al.Differential recognition of ORF2 protein from type 1 and type 2 porcine circoviruses and identification of immunorelevant epitopes[J].JGenVirol,2000,81(Pt 7):1815-1824.
[11] GAMAGE L N,ELLIS J,HAYES S.Immunogenicity of bacteriophage lambda particles displaying porcine circovirus 2(PCV2) capsid protein epitopes[J].Vaccine,2009,27(47):6595-6604.
[12] LI W,WANG X,MA T,et al.Genetic analysis of porcine circovirus type 2(PCV2) strains isolated between 2001 and 2009:genotype PCV2b predominate in postweaning multisystemic wasting syndrome occurrences in eastern China[J].VirusGenes,2010,40(2):244-251.
[13] 吴胜昔,曾 政,蔡家利,等.猪圆环病毒2型CAP蛋白新型表位抗原肽的设计、合成及免疫原性研究[J].免疫学杂志,2014,30(5):397-401. WU S X,ZENG Z,CAI J L,et al.Studies on molecular design,chemical synthesis and immunogenicity of novel antigenic peptides against the capsid protein of porcine circovirus type 2[J].ImmunologicalJournal,2014,30(5):397-401.(in Chinese)
[14] GENMEI L,MANLIN L,RUIAI C,et al.Construction and immunogenicity of recombineant adenovirus expressing ORF2 of PCV2 and porcine IFN gamma[J].Vaccine,2011,29(47):8677-8682.
[15] GAO F,XIE J L,JIA C W,et al.Effects of porcine circovirus type 2 and pseudorabies vaccine co-inoculation on regulatory cytokine mRNA expression in pig peripheral blood mononuclear cells[J].GenetMolRes,2014,13(1):1540-1547.
(编辑 白永平)
Expression and Immunogenicity Analysis of PCV2 Cap Recombinant Protein Antigen which Contains Helper T cell Epitope Peptides
CHEN Shan-zhen,ZHAO Yan,LI Zhong-sheng,LUO Jun,CHEN Ke-hong,WANG Gui-ping,LI Qi-chang*
(GuangdongHaidInstituteofAnimalHusbandry&Veterinary,Guangzhou511400,China)
To determine the role of Th cell epitopes,which were screened from PCV2 ORF1,and ORF3,in improving the PCV2 ORF2 Cap P1 immunogenicity when they were added to the sequence,and the feasibility of synthetic expression of PCV2 ORF2 Cap P1 as a vaccine antigen,bioinformatics and molecular biology methods were employed to screen the peptide sequences.A pan-host of T helper cell epitope peptides and three Th cell epitopes peptide sequences of PCV2-specific were identified.These 4 Th cells peptides sequences were combined with a B-cell epitope peptide sequence screened from PCV2 ORF2 Cap1 gene,and then codon was optimized and inserted the restriction sites and stop codons.After analyzing the codon adaptation index and the frequency distribution,predicted it can achieve efficient expression,further chemical synthesis or peptides were carried out.The synthetic polypeptide antigen P1 was connected to the pET-30a expression vector,and transformed into BL21(DE3) pLysS competent cells,the expression in the engineered bacteria BL21(DE3) pLysS-pET-30a-P1 was constructed.The P1 antigen was able to be expressed after IPTG inducting.The expression level was identified by SDS-PAGE,and its biological activity was identified by Western Blot,followed by mice and pigs immunization with P1.Antibody levels in mice and the peripheral blood lymphocyte proliferation in pigs were tested.The results showed that the codon adaptation index of PCV2 P1 sequence was 0.89,it can achieve to high levels of expression,and the peptide fragment is about 28.29 kDa.MTT results showed that P1 peptide antigen had good immunogenicity.In immunized pigs,the expression of IFN-γ and IL-4 can increase obviously,and were significantly different to the control group(P<0.05).These results indicated that P1 could induce a strong humoral and celluar immune response,and it will provide a theoretical basis on PCV2 P1 peptide antigen as a vaccine antigen for further studies.
PCV2;helper T cell epitope peptides;Cap protein
10.11843/j.issn.0366-6964.2015.12.020
2015-03-31
广东省科技计划星火计划项目(2012A020603026)
陈善真(1984-),女,山东安丘人,兽医师,硕士,主要从事动物疫病诊断及疫苗相关研究,E-mail:chensz2@haid.com.cn
*通信作者:李其昌,博士,E-mail:liqc2@haid.com.cn
S852.4
A
0366-6964(2015)12-2273-09
猜你喜欢
生物学通报(2020年11期)2020-10-22
发明与创新·中学生(2019年6期)2019-06-26
温州医科大学学报(2019年4期)2019-04-28
中成药(2018年7期)2018-08-04
上海农业学报(2017年4期)2017-04-10
中国免疫学杂志(2017年1期)2017-01-17
特产研究(2016年3期)2016-04-12
中国学术期刊文摘(2016年1期)2016-02-13
畜牧兽医学报(2015年3期)2015-07-05
医学研究杂志(2015年6期)2015-07-01
