
热带云雾林植物物种多样性与环境关系研究
2015-12-30 07:35熊梦辉龙文兴杨小波李时兴林明基王茜茜
浙江林业科技 2015年4期
熊梦辉,龙文兴*,杨小波*,黄 瑾,李时兴,林明基,王茜茜,林 灯,康 勇
(1. 海南大学热带作物种质资源保护与开发利用教育部重点实验室,海南大学园艺园林学院,海南 海口 570228;2. 海南省黎母山省级自然保护区管理局,海南 琼中 572929)
热带云雾林植物物种多样性与环境关系研究
熊梦辉1,龙文兴1*,杨小波1*,黄 瑾1,李时兴2,林明基2,王茜茜1,林 灯1,康 勇1
(1. 海南大学热带作物种质资源保护与开发利用教育部重点实验室,海南大学园艺园林学院,海南 海口 570228;2. 海南省黎母山省级自然保护区管理局,海南 琼中 572929)
以海南霸王岭国家级自然保护区热带云雾林为研究对象,调查群落物种组成,测定环境因子,分析种——面积和种——多度之间的关系及起测胸径与物种丰富度、多度、Simpson指数、Shannon-wiener指数的关系,探究物种多样性与环境因子之间的关系。结果表明:热带云雾林共调查到木本植物9 323株,分属40科、70属、109种。其中优势科为金缕梅科、桃金娘科、山矾科和壳斗科等;优势种为蚊母树(Distylium racemosum)、赤楠(Syzygium buxifolium)、九节(Psychotria rubra)和黄杞(Engelhardtia roxburghiana)等。热带云雾林种——面积及种——多度之间的关系均呈对数曲线,最小取样面积及最小多度分别为1 000 m2和1 000;物种丰富度、多度、Simpson指数及Shannon-wiener指数随起测胸径变化呈“倒J”型曲线;当群落包含95%物种多样性时,最大起测胸径约为3 cm。多度、物种丰富度观测值、bootstrap指数与土壤有效氮及有效磷密切相关。
热带云雾林;物种多样性;起测胸径;土壤养分
物种多样性是生物多样性研究的重要内容[1],受土壤等生态因子影响。土壤理化性质的差异,会导致植物物种多样性及空间分布格局的变化[2]。例如,土壤有机质含量与物种多样性密切相关。锡林河流域草原群落及科尔沁退化沙质草地的物种多样性与土壤有机碳呈正相关[3]。而物种丰富度指数、Simpson指数和Shannon-Wiener指数与土壤氮磷养分显著相关,沙质草地表层土壤的全氮、全磷和有效氮与草本植物物种多样性正相关[3]。
起测胸径是影响群落调查结果的重要因素[4],与物种多样性指数大小密切相关。杨少红等发现,乔木层的物种丰富度与胸径显著相关[5]。林进添发现不同起测胸径对物种丰富度影响很大[6];在西双版纳热带山地雨林中,乔木物种丰富度随着起测胸径的增加而依次递减[7]。在物种多样性调查中,往往面临进退两难的境地:如果植物个体起测胸径较小,调查工作量可能会很大;如果植物个体起测胸径较大,调查数据则不能反映群落物种组成情况。如何选取合适的起测胸径,既节省工作量,又充分反映群落的物种多样性特征,是生态学家一直在探讨的问题。
热带云雾林指潮湿热带地区经常被云雾掩盖的森林。与低海拔的热带森林相比,其树木矮小、植株密度大、以小叶革质叶为主,类似于旱生生境植株形态[8~9],群落物种多样性偏低[10]。一天中光合有效辐射、日均空气湿度均呈单峰曲线变化,日平均空气温度低于22℃;日均空气相对湿度在88%以上,呈倒“S”型曲线变化[11]。可见,热带云雾林物种多样性及环境特征与低海拔热带林不同,二者可能有独特的关系。
本文在海南霸王岭热带云雾林设置样地,调查物种多样性,探究植物起测胸径大小与物种多样性的关系;测定群落土壤养分,分析物种多样性与土壤因子的关系,为热带森林植物多样性保护及森林经营管理提供参考。
1 材料与方法
1.1 研究地概况
研究样地位于海南霸王岭国家级自然保护区(18° 50′ ~ 19° 05′ N,109° 05′ ~ 109° 25′ E)。保护区面积约50 000 hm2,以山地为主,海拔100 ~ 1 654 m。该地区属热带季风气候,干湿季明显,5-10月为雨季,11月至次年4月为旱季。海拔100 m位置年平均气温23.6℃,年降水量1 677.1 mm[12]。土壤以砖红壤为代表,随海拔增加逐渐过渡为山地红壤、山地黄壤和山地草甸土[26]。
1.2 群落物种调查
在霸王岭石峰和松林顶热带云雾林中各设置1块样地(表1),在两块样地中分别设置10、11个20 m×20 m的样方。根据相邻格子法将每个20 m×20 m的样方划分为4个10 m×10 m和16个5 m×5 m的小样方。调查样地内所有胸径(DBH)≥1 cm的木本植物,记录其物种名、胸径和坐标。

表1 样地分布Table 1 Distribution of smple plots in Bawangling
1.3 土壤取样与分析
在每个5 m×5 m样方的中心取土样。去掉土壤表层的枯枝落叶,挖0.2 m深的土壤剖面,自上而下取1 kg土样,混合均匀,标记后带到实验室处理,测定土壤有机质、全氮、有效氮、全磷和有效磷含量。
所有土样自然风干后测定其成分含量:有机质用高温外热重铬酸钾氧化—容量法测定;全磷测定用HClO4-H2SO4消化法分解,然后用钼锑抗比色法测定;有效磷用酸性氟化铵浸提,然后用抗坏血酸还原比色测定;全氮用凯氏定氮法,有效氮用碱解扩散法测定[13]。
1.4 数据分析方法
1.4.1 物种组成分析 重要值(IV)的计算方法如下[14]:

式中,Ar为相对多度,Cr为相对显著度,Fr为相对频度。
1.4.2 物种多样性 考虑单个体和双个体种,用刀切法(Jack1法,Jack2法)和抽样调查法(Bootstrap estimator)预测物种丰富度大小。其计算公式如下[15]:

式中,S0表示n个样方内观测到的物种丰富度,r1表示在样方中出现 1次的物种数,r2表示在样方中出现 2次的物种数,hi表示物种i的样方比例。
选用Simpson指数和Shannon-Wiener指数,计算公式如下[16]:
Simpson多样性指数:
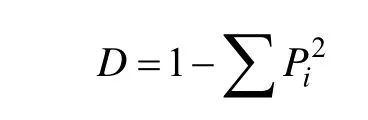
Shannon-Wiener指数:
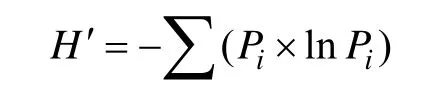
式中,Pi为物种i占所有物种个体总和的比例。
1.4.3 物种丰富度与面积、多度关系及植物多样性与起测胸径的关系 在8 400 m2样方分析种—面积关系。先构建5 m×5 m小样方数据矩阵,随机抽取N个5 m×5 m样方数据放在一起,就可以组成25×N m2面积大小样方数据。对某一面积大小的样方数据进行有放回的重复1 000次随机抽样,取1 000次重复抽样所得物种丰富度的平均值作为该样方面积的物种丰富度。并绘制种—曲线。当群落包含95%的物种丰富度时,求曲线所对应的最小取样面积。
从84 00 m2样方中,随机抽取N个个体构成物种信息数据,计算N个植株个体的物种丰富度,分析物种数与多度关系。重复1 000次随机抽样,取重复抽样的物种丰富度平均数作为N个植株个体的物种丰富度。绘制种—多度曲线。当群落包含95%的物种丰富度时,求曲线所对应的最小多度。
在8 400 m2样方数据中,将所有植株按胸径≥1 cm、≥2 cm、≥3 cm、……≥50 cm建立不同起测胸径的数据库;统计不同起测胸径下的植株多度、物种丰富度、Simpson指数、Shannon-Wiener指数,并绘制起测胸径与多度、物种丰富度、Simpson指数、Shannon-Wiener指数变化曲线。当群落包含95%的物种多样性时,求曲线所对应的最大起测胸径。
1.4.4 物种多样性与土壤环境因子的关系 选择21个20 m×20 m样方,分析群落内物种多样性与土壤养分关系。以土壤有机质、土壤全磷、土壤有效磷、土壤全氮、土壤有效氮含量为自变量,分别以物种丰富度观测值、Jack1预测值、Jack2预测值、bootstrap预测值及多度为因变量,进行逐步线性回归分析。根据AIC值、模型决定系数及P值大小选择最优模型,并得到影响物种多样性的土壤因子。数据分析及作图用R3.1.3软件。
2 研究结果
2.1 物种组成特征
样地内共调查到木本植物9 323株,分属40科、70属、109种。其中优势科为金缕梅科、桃金娘科、山矾科、壳斗科、山茶科和茜草科;优势属为蚊母树属、蒲桃属、山矾属、青冈属、九节属和黄杞属;优势种有蚊母树(Distylium racemosum)、赤楠(Syzygium buxifolium)、九节(Psychotria rubra)、黄杞(Engelhardtiaroxburghiana)、光叶山矾(Symplocos lancifolia)和丛花山矾(S. poilanei)。藤本植物主要为夜花藤(Hypserpa nitida)、巴戟天(Morinda officinalis)、寄生藤(Dendrotrophe frutescens)和山橙(Melodinus suaveolens)。
2.2 物种多样性
物种丰富度观测值为94.50±3.54,Jack1刀切指数为110.41±6.36,Jack2刀切指数为104.96±9.12,bootstrap指数为111.65±2.35,Simpson指数为0.95±0.01,Shannon-Wiener指数为3.54± 0.18,Pielou均匀度指数为0.76 ±0.04。
物种丰富度与取样面积及多度关系均为对数曲线(图1)。当群落包含95%的物种时,最小取样面积约1 000 m2,最小个体数约1 000。
群落多度、物种丰富度、Simpson指数及Shannon-Wiener指数与起测胸径关系都呈“倒J”型曲线(图2A,图2B,图2C和图2D)。当群落包含95%的多度、物种丰富度时,起测胸径最大值都约为3 cm;当群落的Simpson指数和Shannon-Wiener指数达95%时,起测胸径最大值分别为3 cm和4 cm。
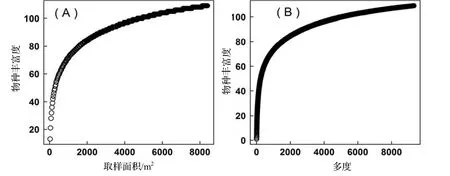
图1 物种丰富度与取样面积、多度的关系Figure 1 Relationship of species richness with the sampling area and abundance

图2 物种多样性指数随起测胸径变化Figure 2 Changes of species diversity with minimum measured diameter
2.3 物种多样性与土壤养分的关系
样地土壤有机质平均含量为(10.73±4.37)%,全磷为0.16±0.06 g/kg,有效磷为16.91±10.94 mg/kg,全氮为2.23±0.75 g/kg,有效氮为156.41±58.71 mg/kg。

表2 不同物种多样性指数与土壤因子的逐步回归Table 2 Stepwise regression analysis of species diversity and soil factors
土壤有效氮含量对群落多度有显著影响(表 2),而土壤有效氮和有效磷对物种丰富度观测值和 bootstrap预测值都有显著影响。Jack1预测值和Jack2预测值与土壤养分的拟合方程不显著。
3 讨论
3.1 物种多样性
霸王岭国家级自然保护区两个样地4 200 m2的样方内,物种丰富度观测值为94.50±3.54,Jack1刀切指数为110.41±6.36,Jack2刀切指数为 104.96±9.12,Bootstrap指数为 111.65±2.35,Simpson指数为 0.95±0.01,Shannon-Wiener指数为3.54±0.18。其物种Shannon-Wiener指数低于尖峰岭热带云雾林(4.71)[17],高于石门台保护区山顶矮林(2.1)[18];而Simpson指数高于海南尖峰岭热带云雾林(0.94)[17],高于石门保护区山顶矮林(0.81)[18]。尖峰林、霸王岭、石门台三地,年降水量和年均温度都为尖峰林>霸王岭>石门台[12,18~19],杨持等认为降水量和年均温度差异可能影响物种多样性[20]。Klopfer认为温度年较差小的地区物种丰富度高于温度年较差大的地区[21],根据Rapoport法则[22],尖峰岭、霸王岭、石门台三个地区的纬度依次增加,温度年较差依次增大,因而物种丰富度呈递减趋势。
3.2 物种多样性与取样面积和起测胸径的关系
两个样地的种—面积曲线和种—多度曲线成对数曲线图形。种—面积关系与前人研究结果一致[23]。热带云雾林的最小取样面积约1 000 m2,低于热带山地雨林2 500 ~ 3 000 m2[24~25]。
物种丰富度、多度、Simpson指数和Shannon-Wiener指数随着起测胸径增大而逐渐减小,呈“倒J”型曲线。植株的胸径级往往对物种丰富度有影响[26]。本研究结果与李宗善等对西双版纳热带山地雨林的研究结果一致[7],也证实了林进添的研究结论,即随着起测胸径的增加,物种丰富度成“倒J”型曲线减小[6]。本文研究发现群落包含95%的物种丰富度、多度、Simpson指数和Shannon-Wiener指数时,植株起测胸径分别为3、3、3、4 cm,该胸径为热带云雾林植物调查的最大胸径,既能节省工作量,又能充分反映群落的物种多样性特征。
3.3 物种多样性与土壤因子的关系
群落多度、物种丰富度和Bootstrap预测指数与土壤有效氮含量正相关;物种丰富度、Bootstrap预测指数与土壤有效磷负相关,说明土壤养分与物种多样性大小密切相关。群落中土壤肥力差异将为植物提供不同水平的营养来源,物种间因资源水平差异形成不同竞争格局,从而影响植物多样性[27]。
氮元素缺乏影响植物正常生长,从而影响物种丰富度[28]。本研究结果与白永飞等在草原群落[3]和徐明锋等[29]在亚热带天然林群落的研究结果一致,植物丰富度与土壤氮密切相关。土壤中H2PO42-易与Al3+和Fe3+形成难溶复合物而不易被植物吸收[30],磷的缺失往往影响植物光合作用,限制植物生长[21],因而常常是热带森林植物的限制因子[30~31]。本文土壤取样时间在雨季,土壤湿度较大,溶解有较多的有效磷,土壤磷差异造成热带云雾林群落组成差异,因而物种丰富度不同。其他研究也证明土壤磷影响植被分布和群落物种组成[29,32]。
[1]马克平. 试论生物多样性的概念[J]. 生物多样性,1993,1(1):20-22.
[2]Mckee K L. Soil physiochemical patterns and mangrove species distribution reciprocal effects[J]. J Ecol, 1993(81):477-487.
[3]白永飞,李凌浩,王其兵,等. 锡林河流域草原群落植物多样性和初级生产力沿水热梯度变化的样带研究[J]. 植物生态学报,2000,24 (6):667-673.
[4]牛丽丽,余新晓,刘彦,等. 不同起测胸径对判定油松分布格局的影响[J]. 北京林业大学学报,2008(S2):12-16.
[5]杨少红,邹文涛,尹光天,等. 广东顺德人工林群落物种多样性与影响因子[J]. 生态科学,2010(5):427-431.
[6]林进添. 天然异龄林资产评估技术改进研究[D]. 福州:福建农林大学,2013.
[7]李宗善,唐建维,郑征,等. 西双版纳热带山地雨林群落乔木树种多样性研究[J]. 应用生态学报,2005,16(7):1 183-1 188.
[8]Nomura N, Kikuzawa K. Productive phenology of tropical montane forest: fertilization experiment along a moisture gradient[J]. Ecol Res 2003(187):573-586.
[9]Williams-Linera G. Tree species richness complementarily, disturbance and fragmentation in a Mexican tropical montane cloud forest[J]. Biodiver Conserv, 2002(11):1 825-1 843.
[10]Bruijnzeel L A, Veneklaas E J. Climatic conditions and tropical montane forest productivity: the fog has not lifted yet[J]. Ecology, 1998(79):3-19.
[11]龙文兴,丁易,臧润国,等. 海南岛霸王岭热带云雾林雨季的环境特征[J]. 植物生态学报,2011,35(2):137-146.
[12]Zang R G, Tao J P, Li C Y. Within community patch dynamics in a tropical montane rain forest of Hainan Island, South China[J]. Acta Oecol, 2005(28):39-48.
[13]中国土壤学会农业化学专业委员会. 土壤农业化学常规分析方法[M]. 北京:北京科学出版社,1983.
[14]王伯荪. 植物群落学[M]. 北京:高等教育出版社,1987.
[15]Palmer M W. The estimation of species richness by extrapolation[J]. Ecology, 1990, 71(3):1 195-1 198.
[16]马克平,刘玉明. 生物群落多样性的测度方法:Iα多样性的测度方法(下)[J]. 生物多样性,1994,2(4):231-239.
[17]欧芷阳,杨小波,吴庆书. 尖峰岭自然保护区扩大区域植物多样性研究[J]. 生物多样性,2007,15(4):437-444.
[18]解丹丹,苏志尧. 石门台自然保护区不同森林群落类型结构组成及物种多样性[J]. 中南林业科技大学学报,2013(11):53-57.
[19]许格希,罗水兴,郭泉水,等. 海南岛尖峰岭12种热带常绿阔叶乔木展叶期与开花期对气候变化的响应[J]. 植物生态学报,2014,38 (6):585-598.
[20]杨持,叶波,邢铁鹏. 草原区区域气候变化对物种多样性的影响[J]. 植物生态学报,1996,20(1):35-40.
[21]Klopfer P H. Environmental determinants of faunal diversity[J]. Am Nat, 1959(93):337-342.
[22]Rapoport E H. Areography: Geographical Strategies of Species[M]. Oxford: Pergamon Press,1982.
[23]姜俊,张春雨,赵秀海. 吉林蛟河42 hm2针阔混交林样地植物种—面积关系[J]. 植物生态学报,2012,36(1):30-38.
[24]李意德,黄全. 对海南岛热带山地雨林植物群落取样面积问题的探讨[J]. 热带林业科技,1986(3):23-29.
[25]朱华,王洪,李保贵,等. 滇南热带雨林物种多样性的取样面积探讨[J]. 生物多样性,1998(4):1-7.
[26]吴裕鹏,许涵,李意德,等. 海南尖峰岭热带山地雨林不同空间尺度和径级水平的物种丰富度与个体密度关联[J]. 植物生态学报,2014 (4):325-333.
[27]Huston M. Soil nutrients and tree species richness in Costa Rican forests[J]. J Biogeogr, 1980(7):147-157.
[28]Whittaker R J, Bush M B, Richards, K. Plant recolonization and vegetation successionon the Krakatau Islands, Indonesia[J]. Ecol Monogr, 1989 (59):59-123.
[29]徐明锋,胡砚秋,李文斌,等. 土壤养分对亚热带天然林物种分布的影响[J]. 中南林业科技大学学报,2014(9):91-97.
[30]Bohn H, McNeal B, O’Connor G. Soil Chemistry thirded[M]. New York: John Wiley & Sons,2001.
[31]Longstreth D J, Nobel P S. Nutrient influences on leaf photosynthesis. Effects of nitrogen, phosphorus and potassium for Gossypium hirsutum L[J]. Plant Physiol, 1980(65):541-543
[32]Walker T W, Syers J K. The fate of phosphorus during pedogenesis[J]. Geoderma, 1976(15):1-19
Relationship between Plant Species Diversity and Habitat in A Tropical Moist Forest
XIONG Meng-hui1,LONG Wen-xing1*,YANG Xiao-bo1*,HUANG jin1,LI Shi-xing2,LIN Ming-ji2,WANG Xi-xi1,LIN Deng1,KANG Yong1
(1. Key Laboratory of Protection and Utilization of Tropical Crop Germplasm Resources, Ministry of Education, College of Horticulture and Landscape Architecture, Hainan University, Haikou 570228, China; 2. Limushan Nature Reserve Administration of Hainan, Qiongzhong 572929, China)
Two sample plots of 8 400 m2were established in Bawanglin National Nature Reserve, Hainan province for investigating species composition of plant communities and habitat factors of the tropical mist forest. The results showed that there are total of 9323 vascular plants, belonging to 109 species of 40 families and 70 genera. The dominant families are Hamamelidaceae, Myrtaceae, Symplocaceae and Fagaceae, and the dominant species are Distylium racemosum, Syzygium buxifolium, Psychotria rubra and Engelhardtia roxburghiana. Relationship of species richness with area and species richness with abundance were logarithmic curves, with the minimum sampling area of 1000 m2and the minimum abundance of 1000. Species richness, abundance, Simpson index and Shannon-wiener index changed with minimum measure diameter, like inverted J-shapedcurves. The maximum measured diameter was 3 cm, the surveyed community had 95% species richness. Community abundance, species richness and bootstrap index had close relationship with soil available nitrogen and phosphorus content.
tropic mist forest; species richness; minimum measured diameter; soil nutrient
S718.51
A
1001-3776(2015)04-0018-06
2015-01-04;
:2015-05-11
国家自然科学基金项目(31260109, 31270474)、海南大学青年基金项目(qnjj1210)、海南省教育厅项目(Hjkj2013-07)、海南省自然科学基金项目(312064)、中科院战略先导专项项目(XDA05050206)和海南大学优秀研究生论文培育计划
熊梦辉(1988-),男,四川渠县人,硕士生,从事植物生态研究;*通讯作者。
猜你喜欢
音乐天地(音乐创作版)(2022年6期)2022-10-14
中学生数理化·八年级物理人教版(2022年4期)2022-04-26
林业勘查设计(2022年1期)2022-02-15
林业科学研究(2021年6期)2022-01-05
内蒙古林业调查设计(2021年5期)2022-01-05
作文·小学低年级(2021年2期)2021-11-02
乡村科技(2021年17期)2021-10-20
安顺学院学报(2021年4期)2021-09-16
林业科技情报(2021年2期)2021-07-13
作文小学中年级(2021年2期)2021-03-15
