
化能异养微生物呼吸与发酵比较
2016-11-08 10:03杨恬然辛明秀
生物学杂志 2016年5期
冯 芬, 杨恬然, 陈 萍, 辛明秀
(北京师范大学 生命科学学院, 北京 100875)
化能异养微生物呼吸与发酵比较
冯 芬, 杨恬然, 陈 萍, 辛明秀
(北京师范大学 生命科学学院, 北京 100875)
化能异养微生物的呼吸和发酵作用是其重要的产能代谢方式,但呼吸与发酵,尤其是无氧呼吸与发酵经常出现混用错用的情况。通过阐述3种产能方式的概念、特点及不同氧浓度下的作用方式,分析其相互联系及本质区别,加深对化能异养微生物有氧呼吸、无氧呼吸及发酵的理解。
化能异养微生物;有氧呼吸;无氧呼吸;发酵
化能异养微生物是一类以有机化合物作为碳源、能源和电子供体的微生物,包括已知的绝大多数细菌和古菌,全部的放线菌、真菌和原生动物[1]。该类微生物通过呼吸(respiration)或发酵(fermentation)产能,但其产能方式由于底物、能量来源及电子供体相同,因此存在很多混用错用的情况。本文通过介绍呼吸与发酵的基本概念和类型,以及它们的相互联系与本质区别,以加深对化能异养微生物产能方式的理解。
1 呼吸
对于大部分化能异养微生物而言,呼吸是其最主要的产能方式。根据最终电子受体及电子传递链组分的不同将呼吸分为有氧呼吸(aerobic respiration)和无氧呼吸(anaerobic respiration)。
1.1 有氧呼吸
在氧气充足条件下,好氧或兼性厌氧微生物以外源分子氧作为最终电子受体时发生的一类高产能效率的呼吸方式[2]。以通用底物葡萄糖为例,有氧呼吸可分为3个阶段:第1阶段是1分子葡萄糖在细胞质基质中通过糖酵解(EMP)产生2分子丙酮酸,该阶段不需要氧气,通过底物水平磷酸化产生少量ATP;第2阶段是丙酮酸氧化脱氢产生的2分子乙酰CoA进入三羧酸循环(TCA)后脱氢,该阶段需要O2的参与,发生在真核生物的线粒体基质或原核生物的细胞基质中[3];第3阶段是上述两个过程中脱下的电子进入电子传递链,与氧化磷酸化相偶联生成ATP,O2与[H+]结合生成水,发生在真核生物的线粒体内膜和原核生物的细胞膜上[4]。总的结果是1分子葡萄糖彻底氧化后生成6分子二氧化碳和12分子水,并产生38分子ATP。
有氧呼吸是化能异养微生物的主要产能方式,利用外源O2将有机底物彻底氧化,并通过电子传递链与氧化磷酸化产生大量能量,满足细胞的生命活动[5]。在3种产能方式中其底物氧化程度最强,利用率最高,产能效率也最高,保证了化能异养微生物的快速繁殖。
1.2 无氧呼吸
在无氧或缺氧条件下,厌氧或兼性厌氧微生物以外源无机氧化物(少数为有机氧化物如延胡索酸)作为最终电子受体时发生的一类产能效率较低的呼吸方式[6]。极大部分进行厌氧呼吸的化能异养微生物是细菌[7],根据最终电子受体的不同可将无氧呼吸分类。
1.2.1 硝酸盐呼吸(nitrate respiration)或反硝化作用(denitrification)
某些细菌如脱氮副球菌(paracoccusdenitrificans)等兼性厌氧菌以硝酸盐作为最终电子受体将硝酸盐还原为亚硝酸、NO、N2O及N2的过程[8]。在农业生产中,由于硝酸盐作为氮源比N2更易利用,反硝化作用会使土壤肥力减弱,同时增加了温室气体排放。但另一方面土壤中的硝酸盐会溶于水流入其他水体,减少硝酸盐的沉积,避免土壤水体变质。硝酸盐呼吸是海水中微生物的主要产能方式,促进自然界氮素循环[9]。
1.2.2 硫酸盐呼吸(sulfate respiration)
脱硫弧菌属(Desulfovibrio)、脱硫杆菌属(Desulfofaba)等的细菌以硫酸盐作为最终电子受体将硫酸盐还原为H2S的过程,这些细菌常常是专性厌氧的硫酸盐还原细菌[10]。硫酸盐呼吸可促进厌氧环境中的有机物循环并对自然界硫素循环具有重要意义[11],但硫酸盐还原会造成地下金属管道腐蚀。
1.2.3 碳酸盐呼吸(carbonate respiration)或碳酸盐还原(carbonation reduction)
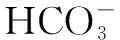
1.2.4 延胡索酸呼吸(fumarate respiration)
一些兼性厌氧菌如巨大脱硫弧菌(Desulfovibriogigas)等以延胡索酸作为外源末端电子受体,还原产生琥珀酸并产能的过程[13]。
除上述类型外,无氧呼吸类型还包括利用Fe3+、Mn2+等作为最终电子受体的产能方式[14],如金属还原地杆菌(Geobactermetallireducens)、奥奈达希瓦氏菌(Shewanellaonedensis)等[15]。
与有氧呼吸相比,无氧呼吸也利用电子传递链与氧化磷酸化相偶联产能,只是其电子受体比氧的标准还原势更低,即电势差小,故产能较少,效率较低。但无氧呼吸却具有非常重要的生物学意义:在长时间的无氧条件下为微生物持续供给能量,保证微生物的生长繁殖,在短时间无氧条件下,微生物也可迅速将产能方式调整至无氧呼吸。同时无氧呼吸也是自然界中氮循环、硫循环中的重要环节。
2 发酵(fermentation)
在无氧或缺氧条件下,有机底物不完全氧化产生的电子不经电子传递系统而直接交给内源性的不完全分解产物,将其还原生成发酵产物,通过底物水平磷酸化产生少量ATP的低效产能反应[16]。根据发酵产物种类的不同将其分类,其中乙醇发酵与乳酸发酵是发酵中的两种基本形式。
2.1 乙醇发酵(alcoholic fermentation)
包括酵母型乙醇发酵和细菌型乙醇发酵[1]。在酵母型乙醇发酵过程中,葡萄糖经EMP途径分解产生的丙酮酸被还原为乙醛,乙醛接受电子还原产生乙醇。细菌型乙醇发酵过程中,葡萄糖经ED途径分解产生丙酮酸,进一步形成乙醇,同时产生少量能量[6]。另外,一些肠道细菌及高温细菌由于缺少丙酮酸脱羧酶而含有乙醛脱氢酶,可利用乙酰辅酶A产生的乙醛进一步还原产生乙醇,且不产生二氧化碳。
2.2乳酸发酵(lactic acid fermentation)
包括同型乳酸发酵和异型乳酸发酵。同型乳酸发酵中葡萄糖经EMP途径产生的丙酮酸直接接受[H+]被还原为乳酸。异型乳酸发酵中葡萄糖经HMP或PK途径产生乳酸及乙醇或乙酸等[17]。
除乙醇发酵与乳酸细菌外,有些化能异养微生物还可进行混合酸发酵(mixed acid fermentation)、丁二醇发酵(butanediol fermentation)等。发酵过程中起始底物只有少部分碳原子被氧化,起始电子供体与内源性电子受体之间的电势差很小,不足以利用电子传递链而只能利用底物水平磷酸化产能,故发酵过程产能最少、效率最低。但发酵却增加了微生物多样性和环境适应性,使其在缺少外来电子受体时也可生长。另外,发酵产物对食品、药品生产具有非常重要的意义。
3 好氧菌、兼性厌氧菌和厌氧菌与呼吸及发酵的关系
根据不同微生物对氧的需求及其耐受能力将化能异养微生物分为好氧菌、兼性厌氧菌及厌氧菌[2]。好氧菌只有在分子氧存在的条件下才能生长繁殖,具有完整的呼吸链,通过有氧呼吸产能,无氧或缺氧条件下其生长受到抑制。如枯草芽孢杆菌(Bacillussubtilis)和铜绿假单胞菌(Pseudomonasaeruginosa)等。兼性厌氧菌在有氧或无氧或缺氧条件下都能生长,具有完整的呼吸链,有氧时通过有氧呼吸产能并迅速分裂增殖,无氧可转换为无氧呼吸或发酵产能[18-19]。如大肠埃希氏菌(Escherichiacoli),在氧气充足时通过有氧呼吸产能并迅速分裂增殖;在无氧条件下,进行以硝酸盐为最终电子受体的无氧呼吸或混合酸发酵[20-21]。一般在有氧条件下,由于电子传递链对NADH+H+的亲和力更高,故兼性厌氧菌会优先利用有氧呼吸[22]。但当某些生长因素改变时,即使在有氧条件下也不会利用有氧呼吸,如当葡萄糖含量超过5%时,酵母细胞中呼吸酶的合成和线粒体的形成被抑制,迫使其进入发酵[23]。
耐氧型厌氧菌不具有呼吸链,故不能利用呼吸产能。该类型菌有氧条件下可以生存,但却不能利用氧气,仅依靠专性发酵或底物水平磷酸化获取能量。如乳酸乳杆菌(Lacticacidbacteria)通过同型乳酸发酵产能供给生长。专性厌氧菌由于不能分解或转化分子氧产生的超氧化物,即使短暂的接触氧也会导致生长抑制甚至死亡。专性厌氧菌不具有完整呼吸链,但一些细菌含有细胞色素,故可利用无氧呼吸或发酵供给其生命活动所需要的能量。如普通脱硫弧菌(Desulfovibriovulgaris)由于含有单一细胞色素C3,可通过无氧呼吸将硫酸盐逐步还原为硫化物。
4 呼吸与发酵的联系与区别
呼吸与发酵的底物均为有机物,都可利用底物水平磷酸化产生能量,对于有氧呼吸及无氧呼吸而言,它们均利用电子传递链与氧化磷酸化相偶联产能,最终电子受体都来源于微生物体外,对于无氧呼吸和发酵而言,它们都是在无氧或缺氧条件下产生少量能量,对有机底物的氧化不彻底。有氧呼吸与发酵能量产生的联系与区别见图1。

图1 有氧呼吸与发酵能量产生的联系与区别
Fig1 Connection and difference between energy production
of aerobic respiration and fermentation
只列出同型乙醇发酵和同型乳酸发酵,所有反应均只标明其还原态氢的产生及流向,其数量、载体与反应的其他产物未示出。
呼吸与发酵之间最本质的区别在于是否利用电子传递链和最终电子受体的性质。发酵仅通过底物水平磷酸化产生少量能量,而呼吸还可利用电子传递链与氧化磷酸化相偶联产能;发酵的最终电子受体是内源性的有机物,而在呼吸作用中,有氧呼吸只能利用外源性的O2,无氧呼吸利用除O2外的外源性无机氧化物或个别有机物。另外呼吸与发酵在其底物氧化程度、代谢微生物等也有区别,详见表1。

表 1 化能异养微生物有氧呼吸、无氧呼吸与发酵异同Table 1 Differences and similarities of aerobic, anaerobic respiration and fermentation
A:在代谢底物相同且等量的情况下
化能异养微生物通过有氧呼吸、无氧呼吸和发酵产能,3种产能策略对于微生物自身生长繁殖、人类工业生产和自然界的物质循环都非常重要。其多种产能方式展现了化能异养微生物多样的生物产能策略,使其能更好地适应环境。
[1]黄秀梨, 辛明秀. 微生物学[M]. 北京:高等教育出版社, 2009:133-149,193.
[2]周德庆. 微生物学教程[M]. 北京:高等教育出版社, 2002:101-119,161-164.
[3]朱钦士. 正转和反转的三羧酸循环[J].生物学通报, 2015, 50(1):16-19.
[4]HAN H, HEMP J, PACE L A, et al. Adaptation of aerobic respiration to low O2environments[J]. Proc Natl Acad Sci U S A, 2011, 108(34):14109-14114.
[5]李兴杰, 陈 超. 关于生物细胞产能代谢类型划分和有关概念界定标准的商榷[J]. 生物学通报, 2009, 44(6):20-23.
[6]李 颖, 文 莹, 关国华, 等. 微生物生理学[M]. 北京:科学出版社, 2009:287-303.
[7]王 晋. 厌氧发酵产酸微生物种群生态及互营关系研究[D]. 无锡:江南大学, 2013.
[8]KRAFT B, STROUS M, TEGETMEYER H E. Microbial nitrate respiration genes, enzymes and environmental distribution[J].Journal of Biotechnology, 2011, 155(1):104-117.
[9]KALVELAGE T, LAVIK G, JENSEN M M, et al. Aerobic microbial respiration in oceanic oxygen minimum zones[J].PLoS ONE, 2015, 10(7):e0133526.
[10]蔡 靖, 郑 平, 张 蕾. 硫酸盐还原菌及其代谢途径[J].科技通报, 2009, 25(4):427-431.
[11]洪义国, 郭 俊, 许志诚, 等. 与环境污染物转化相关的细菌厌氧呼吸研究动态[J]. 应用与环境生物学报, 2006, 12(6):878-883.
[12]李煜珊, 李耀明, 欧阳志云. 产甲烷微生物研究概况[J].环境科学, 2014, 35(5):2025-2030.
[13]KRÖGER A, BIEL S, SIMON J, et al. Fumarate respiration ofWolinellasuccinogenes: enzymology, energetics and coupling mechanism[J].Biochimica et Biophysica Acta, 2002, 1553(1/2):23-38.
[14]许 伟, 胡 佩, 李艳红, 等. 微生物铁呼吸机制研究进展[J].生态学杂志, 2008, 27(6):1037-1042.
[15]李 明. 微生物的无氧呼吸、发酵及其在生产和环境保护上的应用[J]. 生物学通报, 2003, 38(5):20-22.
[16]MÜLLER M, MENTEL M, VAN HELLEMOND J J, et al. Biochemistry and evolution of anaerobic energy metabolism in eukaryotes[J]. Microbiology and Molecular Biology Reviews, 2012, 76(2):444-495.
[17]ABDEL-RAHMAN M A, TASHIRO Y, SONOMOTO K. Recent advances in lactic acid production by microbial fermentation processes[J]. Biotechnology Advances, 2013, 31(6):877-902.
[18]沈 楠. 厌氧微生物体系中还原力的产生及其应用[D]. 合肥:中国科学技术大学, 2014.
[19]Richardson D J. Bacterial respiration: a flexible process for a changing environment[J]. Microbiology, 2000, 146:551-571.
[20]辛明秀. 分子生物学研究的好材料——大肠杆菌(上)(形态结构及生理生化)[J]. 生物学通报, 1996, 31(11):15-17.
[21]李晓敏. 大肠杆菌氧胁迫适应性蛋白质组学、O-抗原研究和嗜热菌NG80-2醛脱氢酶分子生物学研究[D]. 天津:南开大学, 2010.
[22]BUENO E, MESA S, BEDMAR E J, et al. Bacterial adaptation of respiration from oxic to microoxic and anoxic conditions: redox control[J]. Antioxid Redox Signal, 2012, 16(8):819-852.
[23]周景文, 董志姚, 刘立明, 等. 葡萄糖酸钠对光滑球拟酵母CCTCC M202019能量代谢和丙酮酸积累的影响[J].华中农业大学学报, 2010, 29(4):527-532.
Comparison between respiration and fermentation of chemoheterotrophic microorganisms
FENG Fen, YANG Tian-ran, CHEN Ping, XIN Ming-xiu
(College of Life Sciences, Beijing Normal University, Beijing 100875, China)
Chemoheterotrophic microorganisms obtain energy by respiration or fermentation. However, respiration or fermentation, especially anaerobic respiration and fermentation are usually misused in many situations. The concepts, characteristics and utilization of aerobic respiration, anaerobic respiration and fermentation were compared. The relationship and difference between them were also analyzed in this paper.
chemoheterotrophic microorganisms; aerobic respiration; anaerobic respiration; fermentation
2015-12-23;
2016-01-07
国家自然基金(31470251)
冯芬,硕士,主要研究方向为微生物生理生化,E-mail:fengfen@mail.bnu.edu.cn
辛明秀,博士,副教授,主要研究方向为微生物生理生化,E-mail:xinmingxiu@bnu.edu.cn
Q935
A
2095-1736(2016)05-0083-04
doi∶10.3969/j.issn.2095-1736.2016.05.083
猜你喜欢
云南化工(2021年6期)2021-12-21
河北医药(2020年23期)2020-12-10
科学(2020年2期)2020-08-24
生物工程学报(2020年6期)2020-07-31
中国保健营养(2019年2期)2019-10-21
疯狂英语·新读写(2018年3期)2018-11-29
检验医学(2018年2期)2018-03-07
山西建筑(2014年6期)2014-11-09
天津医科大学学报(2011年4期)2011-07-13
