
小齿猬的染色体研究
2016-12-12 10:22孔飞郭健民吴家炎
四川动物 2016年2期
孔飞, 郭健民, 吴家炎
(1. 陕西省动物研究所,西安710032; 2. 海南师范大学,海口571158)
DOI:10.11984/j.issn.1000-7083.20150309
小齿猬的染色体研究
孔飞1, 2, 郭健民1, 吴家炎1
(1. 陕西省动物研究所,西安710032; 2. 海南师范大学,海口571158)
采用骨髓细胞染色体标本制片法对分布在陕西榆林小壕兔地区的小齿猬Mesechinusmiodon种群的染色体进行研究。结果表明,小齿猬二倍体染色体数目为44,臂数NF=87,同时由于存在数目不等的B染色体,使得观察计数染色体2n=46,2n=48的中期细胞出现频率有所升高,故小齿猬核型可以简化描述为2n=♂44~48,NF=87~95,18(m)+24(sm)+0~4 B m(m或sm),XY(st,t),支持Thomas等将miodon认作独立种的观点。
小齿猬;B染色体;核型
小齿猬Mesechinusmiodon隶属于劳亚食虫目Eulipotyphla猬科Erinaceidae猬亚科Erinaceinae林猬属Mesechinus,分布于陕西省榆林地区沿毛乌素沙漠一线,除榆林外,紧靠内蒙古自治区乌审旗,伊金霍洛旗的鄂尔多斯市,以及神木县可能也有分布。Thomas于陕西榆林小壕兔地区获得该种并命名为Erinaceusmiodon(Thomas,1908)。王应祥(2003)因其第二个上臼齿(P2)均小于2 mm等特点,将其中文学名定为“小齿猬”。
从陕西榆林的miodon发现以来,涉及到它的分类地位基本上有2种观点。一种观点是把它当成达乌尔猬Mesechinusdauuricus或亚种(Allen,1938;Ellerman & Morrison-Scott,1951;Corbet,1978;林炜,闵芝兰,1989),另一种观点是将miodon认作独立的种(Thomas,1908;LÖonberg,1922;王应祥,2003)。目前关于小齿猬的分类地位和染色体核型尚未见报道。本文通过对小齿猬染色体数目及核型的研究,明确了该种的分类地位,为其他相关研究积累资料。
1 材料和方法
1.1 实验材料
2012—2014年每年8—10月,采用笼捕的方法在陕西榆林小壕兔地区共捕捉到小齿猬7只(YX 1202♂、YX 1301♀、YX 1302♀、YX 1303♂、YX 1304♂、YX 1401♂、YX 1402♀,标本现保存于陕西省动物研究所标本馆),将其带回到陕西省动物研究所实验室内进行饲养和染色体研究。
1.2 实验方法
1.2.1 染色体的制备 采用骨髓细胞固定法制备染色体标本。具体操作方法为:给每只小齿猬腹腔注射1%秋水仙素溶液,剂量为每千克体质量4 mg。4 h后处死,取其肱骨和股骨,剔净骨上的肌肉后将两端的骨骺横断,用注射器吸入0.46%KCl溶液冲出骨髓细胞,37 ℃低渗溶液处理30 min。1 000 r·min-1离心10 min,去掉上清液后加入预冷新配置的卡诺氏固定液(V甲醇∶V冰醋酸=3∶1)固定,10 min后1 000 r·min-1离心10 min,并重复固定2次;在最后一次固定并离心后,弃去上清液,加入0.5 mL卡诺氏固定液,制成悬浊液并滴于预先冰冻的载玻片上,空气干燥过夜。载玻片用10%Giemsa磷酸液染色30 min后自来水冲洗干净,待干燥后在显微油镜下进行染色体的照相和计数分析。
1.2.2 核型分析 使用Leica DC500显微镜油镜进行观察、统计染色体数目,选取来自不同个体分散良好的染色体中期分裂相进行照相。拍照后计数并确定基数最大的为其染色体数目。选取染色体完整、分散良好、边缘清晰且较平整的中期细胞分裂相,用Adobe Photoshop软件处理图片、配对,并测量计算染色体的相对长度、臂比值、臂数等。按Levan等(1964)提出的染色体分组标准进行染色体分类和臂数统计。
2 结果
2.1 染色体数目
对小齿猬染色体数目的统计表明,在计数的150个中期细胞中具有44条染色体的细胞占多数,达46%,因此可以说小齿猬的染色体数目为2n=44。另外表中也反映出2n=46,2n=48的细胞以较多的频率出现,分别为17.3%和19.3%(表1)。
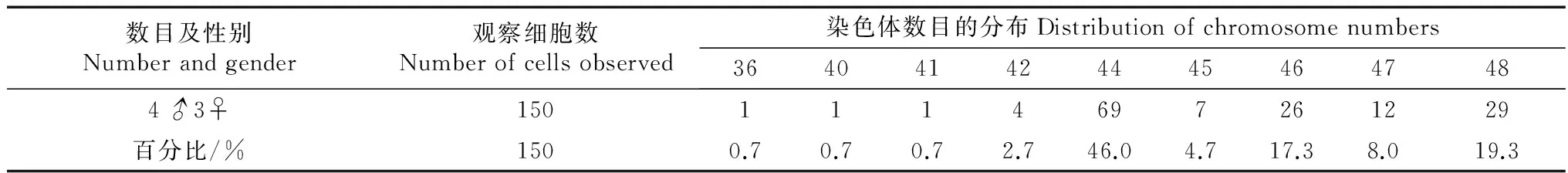
表1 小齿猬二倍体染色体数目统计
2.2 染色体核型
选择染色体形态清晰、分散较好的分裂相进行拍照,测量染色体的相对长度、臂比,并分组(表2)。
小齿猬有丝分裂中期细胞染色体都是大小不同的双臂染色体,仅Y染色体是很小的单臂染色体。经核型分析,可以看出在2n=44的细胞中有9对中部着丝粒染色体,其中1~4号较大,形态清晰;12对亚中部着丝粒染色体,其中10~13号较大,易辨别,10号亚中部着丝粒染色体是核型中最大的1对双臂染色体。X染色体为中等大小的亚端部着丝粒染色体,Y染色体是核型中最小的不成对的末端着丝粒染色体。在2n=44,NF=87染色体组型中,为了便于观察和分析,常染色体还可以分为A、B、C、D 4个组,A组为4对较大的中部着丝粒染色体:1~4号;B组是5对中部着丝粒染色体:5~9号;C组是4对较大的亚中部着丝粒染色体:10~13号;D组14~21号是8对亚中部着丝粒染色体(图1)。在2n=46的细胞中,多了1对B组的最小中部着丝粒染色体;2n=48的核型中,B组多了1对小的中部着丝粒染色体,D组多了1对最小的亚中部着丝粒染色体。2n=47的细胞出现较少,多了3个最小的中部着丝粒和亚中部着丝粒染色体。这些正常核型中多出来的小染色体即超数染色体,也称B染色体。数目不等的B染色体使得观察计数染色体时,2n=46,2n=48的有丝分裂中期细胞出现频率有所升高。根据以上分析,小齿猬的核型可以简化描述为2n=♂44~48,NF=87~95,18(m) +24(sm)+0~4 B m(m或sm),XY(st,t)。

表2 小齿猬的染色体参数及类型


图1 小齿猬的核型(2n=44,NF=87,X和Y为性染色体)
3 讨论
该研究表明小齿猬的染色体组型为2n=♂44~48,NF=87~95,18(m)+24(sm)+0~4 B m(m或sm),XY(st,t),有别于KopaσπeB等(1996)报告的达乌尔猬的染色体组型为2n=48,NF=92,10(st)+22(m)+14(sm),性染色体XY(sm,mt)。闵芝兰和林炜(1989)也专题研究了陕西省的猬科分类,并由林炜专门采集了Thomas(1908)所订的Erinaceusmiodon模式产地——陕西榆林活体标本,并进行了染色体组型测定,其结果为2n=44,NF=82,10(sm)+30(m)+2(t),XY(m,t)(林炜,闵芝兰,1989)。这和我们所做核型也有很大的相似性,染色体数目相同,其中的差别仅仅是在较小的常染色体及性染色体的形态识别划分上,但其认定该标本是达乌尔猬,显然与KopaσπeB等(1996)报道在达乌尔猬指名亚种的模式产地测得的达乌尔猬染色体组型结果不同。参考形态性状特点,我们认为林炜和闵芝兰(1989)所测的榆林标本正是Thomas(1908)当年所订立的Erinaceusmiodon,应恢复Thomas(1908)提出在陕西榆林地区获得的Mesechinusmiodon种的地位,并支持王应祥等因其第二上臼齿(P2)均小于2 mm等特点,将其中文学名定为“小齿猬”。
高等动物的染色体数目及组型比低等动物稳定,但染色体数目和结构的变化是物种进化的重要因素之一(陈广文等,2010)。根据已发表的资料统计出我国分布的猬科4种的染色体数目为2n=48(陈宜峰,郭健民,1986;林炜,闵芝兰,1989;杨凤堂等,1991;汪松等,2001),而本研究小齿猬染色体数目为44~48,这种多余染色体的性质,为超数染色体。超数染色体即B染色体,是某些物种中除A染色体之外的一些额外染色体,多数由异染色质组成,没有强烈的整倍体效应,存在着种群间、种群内差异。束峰珏等(1999)研究发现,由于B染色体在传递过程中的不稳定性,常常造成染色体数目和形态在种群间、个体间的差异。B染色体在植物和昆虫中比较常见,现在已知植物有600多种,昆虫有158种都存在B染色体,而哺乳动物中较为少见。牧野佐二郎(1982)报道了赤狐Vulpesvulpes染色体组型也存在微小的B染色体,使染色体数目有所变异,2n=♂34~38。束峰珏等(1999)首次提出毛冠鹿ElaphoduscaphalophusB染色体多态,并对其传递机制进行了探讨。在啮齿类大仓鼠Tscherkiatriton、鄂毕环颈旅鼠Dicrostonyxtorguatus、印度灌鼠Golundaellioti、昆士兰大裸尾鼠Uromyscaudimaculatus、黄足裸尾鼠Melomyscervinipes、宽齿鼠Pseudomysfuscus、沼泽鼠Rattusfuscipes等物种中都存在数目不等、大小不一的B染色体。在黑家鼠R.rattus中,无或有2个中部着丝粒染色体,可使它的染色体数目为2n=♂38或40。本文报道的食虫类小齿猬存在B染色体还属首次报道,由于一般认为B染色体主要为异染色质,且有个体小、形态不清晰的特点,其来源及遗传功能有待继续深入研究。
致谢:研究工作进行中,承中国科学院昆明动物研究所王应祥教授,西北大学杨兴中教授、李智选教授,陕西师范大学于小平教授、李晓晨教授,中国人民解放军第四军医大学师长宏教授,榆林市人民政府动物管理站胡彩娥站长、李怀杰站长,榆林市榆阳区小壕兔乡杨世雄乡长,小壕兔村张起云村长以及陕西省动物研究所吴晓民研究员及蒋志武、党蕊叶、陈洁、张广平等大力协助、诚恳指导,特此一并致谢。
陈广文, 董自梅, 田士瑞, 等. 2010. 河北3 产地日本三角涡虫的染色体变异与核型多样性分析[J]. 动物学杂志, 45(6): 85-91.
陈宜峰, 郭健民. 1986. 哺乳动物染色体[M]. 北京: 科学出版社: 8.
林炜, 闵芝兰. 1989. 两种短棘猬染色体组型分析[J]. 西北大学学报, 19(1): 69-72.
闵芝兰, 林炜. 1989. 陕西省猬科(Erinaceidae)分类的研究[J]. 西北大学学报, 19(4): 71-79.
束峰珏, 张锡然, 曹祥荣, 等. 1999. 毛冠鹿B染色体多态及遗传机制探讨[J]. 遗传, 21(6): 23-26.
汪松, 解焱, 王家俊. 2001. 世界哺乳动物名典[M]. 长沙: 湖南教育出版社: 17-18.
王应祥. 2003. 中国哺乳动物种和亚种分类名录与分布大全[M]. 北京: 中国林业出版社: 2-4.
杨凤堂, 马彩霞, 施立明. 1991. 远东刺猬和大耳猬的核型分析[J]. 动物学研究, 12(4): 393-398.
牧野佐二郎. 1982. 放射線·化学物質と染色体異常[D]. 医学書院.
Allen GM. 1938-1940. Mammals of China and Mongolia vols Ⅰ[M]. New York: The American Museum Natural: 43-54.
Corbet GB. 1978.The mammals of the Palaearctic region: a taxonomic review[M]. British Museum (Natural History): Cornell University Press: 15.
Ellerman JR, Morrison-Scott TCS. 1951. Checklist of Palaearctic and Indian mammals[M]. London: British Museum: 14-26.
Levan A, Fredga K, Sandberg AA. 1964. Nomendature for cetrometric position on chromosome[J]. Hereditas, 52: 201-220.
LÖunberg E. 1922. Some remarks about eastern hedgehogs[J]. Annals and Magazine of Natural History, 9(9): 620-629.
Thomas O. 1908. The duke of Bedford's zoological exploration in eastern Asia-Ⅺ on mammals from the provinces of Shansi and Shensi, northern China[J]. Proceedings of the Zoological Society London: 963-983.Кораблев ВП, Кирилюк ВЕ, Головушкин МИ. 1996. ИССЛЕДВАНИ Е КА Р И-ОТИПА ДАУРСКОГОЕ ЖАMesechinusdauricus(Mammalia, Erinaceidae) ИЗ TERRA TYPICA[J]. ЗОО Л. ЖУ РНАЛ. том 75, вып. 4: 558-564 (with English summer).
四川省动物学会第十届会员代表大会暨第十届第一次理事会会议纪要
2015年9月18—20日,四川省动物学会第十届会员代表大会暨第十一次学术研讨会在乐山师范学院召开。本次研讨会由四川省动物学会、四川省野生动植物保护协会主办,乐山师范学院承办。来自全省20个单位的120多名动物学工作者参加了本次会议。四川省动物学会第十届会员代表大会进行了换届选举和章程修改等工作。四川省动物学会理事长岳碧松教授代表第九届理事会做了学会工作报告,李德生理事做了学会章程修订说明。会议经过民主选举产生了四川省动物学会第十届理事会。第十届第一次理事会于19日晚召开,选举了理事长、副理事长、常务理事,任命了秘书长,学会秘书处设在四川大学生命科学学院内。
四川省动物学会第十届理事会名单
理事长: 岳碧松
副理事长: 刘建雄、唐业忠、王红宁、王永、周材权
秘书长: 冉江洪
常务理事: 窦丰满、杜军、江建平(新增)、郭鹏、乐天、李德生(新增)、李建国、刘建雄、刘少英、冉江洪、唐业忠、王强、王红宁、王永、岳碧松、曾晓茂、张和民、张志和、张泽钧(新增)、周材权
理 事: 曹成全(新增)、陈红卫、陈建平、窦丰满、杜军、龚继恩、顾海军、郭鹏、郭宪光、侯蓉、黄炎(新增)、江建平、乐天、李德生、李家堂(新增)、李建国、李贤中、廖文波(新增)、刘昊(新增)、刘建雄、刘少英、孟杨(新增)、齐敦武(新增)、钱微萍、冉江洪、沈富军(新增)、石谦、石爱民(新增)、宋昭彬、孙治宇、谭速进、唐业忠、王强、王永、王光西、王红宁、王鹏彦、王雅静、王淯(新增)、徐怀亮、杨文、杨光友、余建秋(新增)、岳碧松、曾晓茂、曾燏(新增)、张贵权(新增)、张和民、张泽钧、张志和、周材权
四川省动物学会
2015-09-21
Study on the Chromosome ofMesechinusmiodon
KONG Fei1, 2, GUO Jianmin1, WU Jiayan1
(1. Shaanxi Institute of Zoology, Xi’an 710032, China; 2. Hainan Normal University, Haikou 571158, China)
The chromosome ofMesechinusmiodonfrom Shaanxi was studied using medulla cells colchicine-hypotonic air-drying technique. The basic diploid and fundamental autosomal arm numbers were 44 and 87, respectively. Moreover, chromosomal variation was observed which might be due to the existence of B-chromosomes. The karyotype formula was 2n=♂44~48, NF=87~95, 18(m)+24(sm)+0~4 B m (m or sm), XY (st, t). The results suggested thatMesechinusmiodonshould be defined as a valid species rather than a synonym ofM.dauuricus.
Mesechinusmiodon; B chromosome; karyotype
2015-10-09 接受日期:2015-12-15 基金项目:陕西省科学院科技计划项目(2014K-16)
孔飞, 助理研究员, 研究方向: 动物生态学, E-mail:k.coffee@163.com
Q959.8
A
1000-7083(2016)02-0217-04
猜你喜欢
生物技术进展(2022年1期)2022-02-11
小猕猴智力画刊(2021年6期)2021-08-05
宁夏医学杂志(2020年3期)2021-01-21
科学之谜(2018年9期)2018-12-17
中国民族医药杂志(2016年5期)2016-05-09
作文大王·低年级(2016年3期)2016-03-11
哈尔滨医药(2015年2期)2015-12-01
中国当代医药(2015年9期)2015-03-01
遗传(2015年4期)2015-02-04
应用海洋学学报(2014年3期)2014-11-22
