
杆状病毒Ac110蛋白的保守氨基酸位点N27对病毒在中肠建立有效感染起重要作用
2017-08-02 01:39朱乐园刘建涛王燕玲袁美妗吴文碧
环境昆虫学报 2017年3期
朱乐园,刘建涛, 2,王燕玲,袁美妗,吴文碧*,杨 凯
杆状病毒Ac110蛋白的保守氨基酸位点N27对病毒在中肠建立有效感染起重要作用
朱乐园1,刘建涛1, 2,王燕玲1,袁美妗1,吴文碧1*,杨 凯1
( 1.中山大学生命科学学院,有害生物控制与资源利用国家重点实验室,广州 510275;2.江西科技师范大学生命科学学院,南昌 330013)
杆状病毒是节肢动物,特别是鳞翅目昆虫的重要病原微生物,在生物防治中有着广泛的应用前景。其对昆虫虫体口服感染的过程中,需要一类称为杆状病毒口服感染因子的帮助才能在虫体中肠成功建立系统感染。Ac110是本实验室最新发现的口服感染因子之一,但其作用机制仍是未知。在本研究中,通过定点突变技术成功构建了两株Ac110保守氨基酸位点突变型重组杆状病毒,将病毒DNA转染昆虫细胞Sf9后,通过蛋白免疫印迹实验验证了Ac110的突变体蛋白均能够正常表达;接着通过病毒滴度测定实验证明了重组病毒均能在Sf9细胞中产生与补回型病毒相当的感染性病毒粒子,说明Ac110的这两个保守氨基酸位点的突变并不影响病毒在细胞中的复制和在细胞之间的传播;最后将纯化的病毒多角体口服感染甜菜夜蛾幼虫进行生物测定实验,发现Ac110的N27氨基酸位点的突变对昆虫幼虫的口服感染能力显著性降低,而L35氨基酸位点的突变则没有影响。
杆状病毒;口服感染因子;Ac110;生物测定
杆状病毒 (Baculovirus)是一类寄生于节肢动物的专一性病原微生物,其宿主主要为昆虫纲中的鳞翅目、双翅目、膜翅目和鞘翅目昆虫。杆状病毒的病毒粒子呈杆状,基因组为双链闭合环状 DNA,病毒基因组大小介于80-180 kb 之间。杆状病毒是目前对人类最为有益的病毒,不仅对人的健康无害,在控制农林害虫的种群密度方面更是起到重要作用。杆状病毒在其生活周期中产生两种类型的病毒粒子:芽生型病毒粒子BV (Budded virus)和包埋型病毒粒子ODV (Occlusion derived virus)。BV出芽时在核膜或细胞膜处获得囊膜,而囊膜可以介导病毒与宿主细胞表面特定的受体相结合,对病毒粒子在虫体内的传播及病毒在离体培养细胞间的传播起到重要作用;ODV包埋于由多角体蛋白质组成的包涵体蛋白晶体中,形成多角体(Occlusion body, OB),而多角体还被多角体囊膜(polyhedron envelope, PE)包被。多角体在自然环境中非常稳定,对包埋在其中的病毒粒子起到保护作用,利于病毒在自然界中的传播(Braunageletal., 2003)。
杆状病毒感染昆虫是一个较为复杂的过程,昆虫摄食了含有杆状病毒多角体的食物后,多角体入宿主中肠腔体,在中肠的碱性环境下被裂解,释放其包裹的ODV。随后,ODV要穿越多重障碍进入昆虫宿主中肠上皮细胞建立初始感染,其中第一个障碍就是宿主肠道的保护屏障围食膜(peritrophic matrix, PM)。PM是介于宿主中肠上皮细胞和肠腔间的网状结构,由几丁质、糖蛋白和几丁质结合蛋白组成(Hegedusetal., 2009)。ODV的大小远大于PM的孔径,因此ODV除了可能借助由于宿主吞食食物过程中造成PM机械损伤导致其孔径变大使得ODV顺利穿越之外(Wang and Granados, 2000),还可能借助多角体自身包裹的杆状病毒编码的Enhancin和金属蛋白酶等因子对PM起到降解的作用(Leporeetal., 1996),从而使ODV顺利穿越PM到达中肠上皮细胞(Ishimweetal., 2015)。ODV到达中肠上皮细胞之后,需要穿过第二个障碍即中肠上皮细胞微绒毛而进入中肠上皮细胞,已有研究表明ODV通过膜融合,即ODV囊膜与中肠柱状上皮细胞的微绒毛发生结合和融合,释放出ODV中的核衣壳,使其进入中肠上皮细胞,从而成功建立初始感染(Faulkneretal., 1997)。而这个过程需要杆状病毒编码的一类称为口服感染因子(perosinfectivity factor, PIF)的蛋白发挥至关重要的作用。
目前已发现的杆状病毒PIF包括了P74 (PIF-0或Ac138) (Faulkneretal., 1997)、PIF-1 (Ac119) (Kikhnoetal., 2002)、PIF-2 (Ac22) (Fangetal., 2006; Pijlmanetal., 2003)、PIF-3 (Ac115) (Ohkawaetal., 2005)、PIF-4 (Ac96) (Fangetal., 2009)、PIF-5 (Ac148) (Harrisonetal., 2010; Xiangetal., 2011)、PIF-6 (Ac68) (Nieetal., 2012)、PIF-8 (Ac83) (Javedetal., 2017; Zhuetal., 2013)、以及本实验室发现的PIF-7 (Ac110) (Javedetal., 2017; Liuetal., 2016)。已有的研究表明,在这些PIFs中,缺失其中的任何一个都会极大降低杆状病毒对宿主的口服感染能力。同时发现很多PIF之间存在相互作用,可能共同行使着促进杆状病毒在宿主中肠细胞建立初始感染的功能。
本实验室前期的研究中,通过构建Ac110缺失型和补回型重组病毒,发现Ac110是杆状病毒一个新的口服感染因子,而且PM并不是其作用位点(Liuetal., 2016)。为了对Ac110的功能进行深入的探究,我们对Ac110的两个保守氨基酸位点分别进行了定点突变,构建出两株Ac110突变型重组病毒,研究其对Ac110功能的影响。
1 材料和方法
1.1 实验昆虫,细胞和病毒
Sf9细胞:草地贪夜蛾Spodopterafrugiperda细胞Sf9购自英潍捷基(上海)公司。甜菜夜蛾幼虫:由中山大学昆虫学研究所昆虫饲养室饲养提供。甜菜夜蛾幼虫饲料:由中山大学昆虫学研究所昆虫饲养室提供。
Ac110缺失型重组病毒vAc110KO和补回型重组病毒vAc110HA为本实验室保存(Liuetal., 2016)。
1.2 Ac110氨基酸序列分析
利用 NCBI 网站上的核苷酸序列 BLAST 搜索软件(protein-protein blast, blastp)搜索Ac110同源蛋白的氨基酸序列并将序列下载。运用Clustal W 1.83软件对 AcMNPV Ac110蛋白同源物的氨基酸序列进行多序列比对分析,通过Genedoc软件进行编辑。
1.3 Ac110点突变重组病毒的构建
通过定点突变技术(Chiuetal., 2004)和Bac-to-Bac Expression System(Invitrogen)提供特异性转座,构建Ac110突变型重组病毒。首先在每个需要突变的位点设计四条引物(表1所示),以pUC110HA-SV40质粒DNA (Liuetal., 2016)(此质粒以pUC18质粒为骨架,插入包含ac110自身启动子、开放阅读框和多聚腺苷酸的片段,是构建补回型重组病毒转座载体的中间载体)为模板进行PCR反应,以获得pUC18-Ac110N27A和pUC18-Ac110L35A质粒;通过酶切将Ac110N27A和Ac110L35A片段从pUC18-Ac110N27A和pUC18-Ac110L35A中切下来并连接至pFB1-PH-GFP (Wuetal., 2006)载体上,构建出转座载体pFB1-Ac110N27A-PH-GFP和pFB1-Ac110L35A-PH-GFP。将转座载体分别转化至含有Ac110缺失型重组病毒bAc110KO (Liuetal., 2016)的大肠杆菌细胞DH10B中,通过特异性位点转座将ac110点突变型基因转座至缺失了ac110基因的杆状病毒bAc110KO中,再通过蓝白斑筛选出带有绿色荧光基因蛋白gfp和多角体蛋白基因polyhedrin的Ac110点突变重组病毒vAc110N27A和vAc110L35A。最后通过PCR和测序对重组病毒进行鉴定。

表1 Ac110点突变引物序列Table 1 Sequences of primers for Ac110 site mutation
1.4 病毒对细胞的转染和感染
病毒的转染和感染根据文献(Wuetal., 2006)操作进行:首先把1×106个处于对数生长期的Sf9细胞接种于直径为35 mm的细胞培养皿中,然后将重组病毒DNA与脂质体混匀,室温温育45 min,再加入到细胞上孵育5 h,最后吸除孵育细胞的病毒和脂质体混合液,给细胞换上新鲜的培养基,并以此时间点作为病毒转染后0 h来计算病毒转染的时间。
病毒感染实验的操作如下:将病毒液以一定的感染复数(multiplicity of infection, MOI)接种于细胞并孵育1 h,然后吸除孵育细胞的病毒液,给细胞换上新鲜的培养基,并以此时间点作为病毒感染后0 h来计算病毒感染的时间。
1.5 病毒滴度测定及生长曲线的绘制
在病毒感染后的不同时间点取细胞上清液进行病毒滴度的检测,采用TCID50(50% tissue culture infective dose) (Wuetal., 2006)终点稀释法对样品中可感染性病毒粒子BV进行定量。运用GraphPad Prism 6.0软件,根据每个时间点的病毒滴度绘制病毒生长曲线。
1.6 蛋白免疫印迹实验(Western blotting)
蛋白免疫印迹实验的方法参考(Wuetal., 2008)文献:将病毒感染后的细胞离心收集后,用PBS清洗两次,加入蛋白裂解液制备蛋白样品。蛋白样品进行SDS-PAGE胶分离后,通过半干转膜系统将蛋白转印到PVDF膜,以HA兔单抗(Cell Signal Technology)或者β-actin鼠单抗(Proteintech)为一抗和辣根过氧化物酶HRP标记的二抗进行蛋白杂交,最后通过ECL (Pierce ECL Western Blotting Substrate)进行胶片显影。
1.7 病毒的虫体扩增及纯化
多角体的扩增和纯化参考(O’Reilly, 1992)文献中的操作:将病毒多角体涂布于人工饲料表面饲喂感染3龄的昆虫幼虫,并于幼虫发病死亡后收集虫尸。首先加入PBS用匀浆器充分碾磨均匀虫尸,匀浆液先用 4 层纱布过滤,再用 6 层纱布过滤,除去组织碎片,收集滤液。室温下4500 rpm离心30 min,弃上清;以PBS重悬沉淀,室温下500 rpm离心10 min,取上清,8000 rpm离心10 min,取沉淀(重复此步骤5-10 次);以0.05% SDS重悬沉淀,室温下 500 rpm离心10 min,取上清,8000 rpm离心10 min,取沉淀(重复此步骤5-10 次);以ddH2O 重悬沉淀,室温下 500 rpm离心 10 min,取上清,8000 rpm离心 10 min,取沉淀(重复此步骤5-10次);最后沉淀重悬于ddH2O,即为较纯的多角体,应为均质乳白状;置于-20℃保存备用。
1.8 生物测定实验
将收集纯化的病毒多角体进行计数并稀释至相应的浓度,将饲料切成1 mm见方的大小,涂抹1 μL多角体悬液于饲料表面喂食已饥饿处理的甜菜夜蛾3龄幼虫,待幼虫吃完涂有病毒多角体的饲料后再喂食新鲜的饲料。每天喂食足够的饲料并观察幼虫的感染情况。
2 结果与分析
2.1 Ac110同源蛋白氨基酸序列同源性及保守氨基酸位点分析
Ac110是杆状病毒中高度保守的基因(Garavagliaetal., 2012),目前认为它是杆状病毒中第38个核心基因(Javedetal., 2017)。通过NCBI网站上的blastp进行搜索,将Ac110的部分同源蛋白的氨基酸序列下载并进行分析。表2是所下载的Ac110同源蛋白的信息。通过多序列比对软件对Ac110同源蛋白的氨基酸序列进行同源性分析,结果显示Ac110序列与PlxyMNPV、BmMNPV、OpNPV和AngeNPV 对应ORF的一致性可达到69%-98%,与其他鳞翅目杆状病毒同源蛋白一致性在26%-48%。通过多序列比对软件对Ac110同源蛋白的氨基酸序列进行多序列比对,结果如图1所示,所有进行比对的氨基酸序列当中,有11个氨基酸残基是高度保守的(图1)。

表2 杆状病毒Ac110及其同源蛋白 Table 2 Baculovirus Ac110 and its homologs
续上表
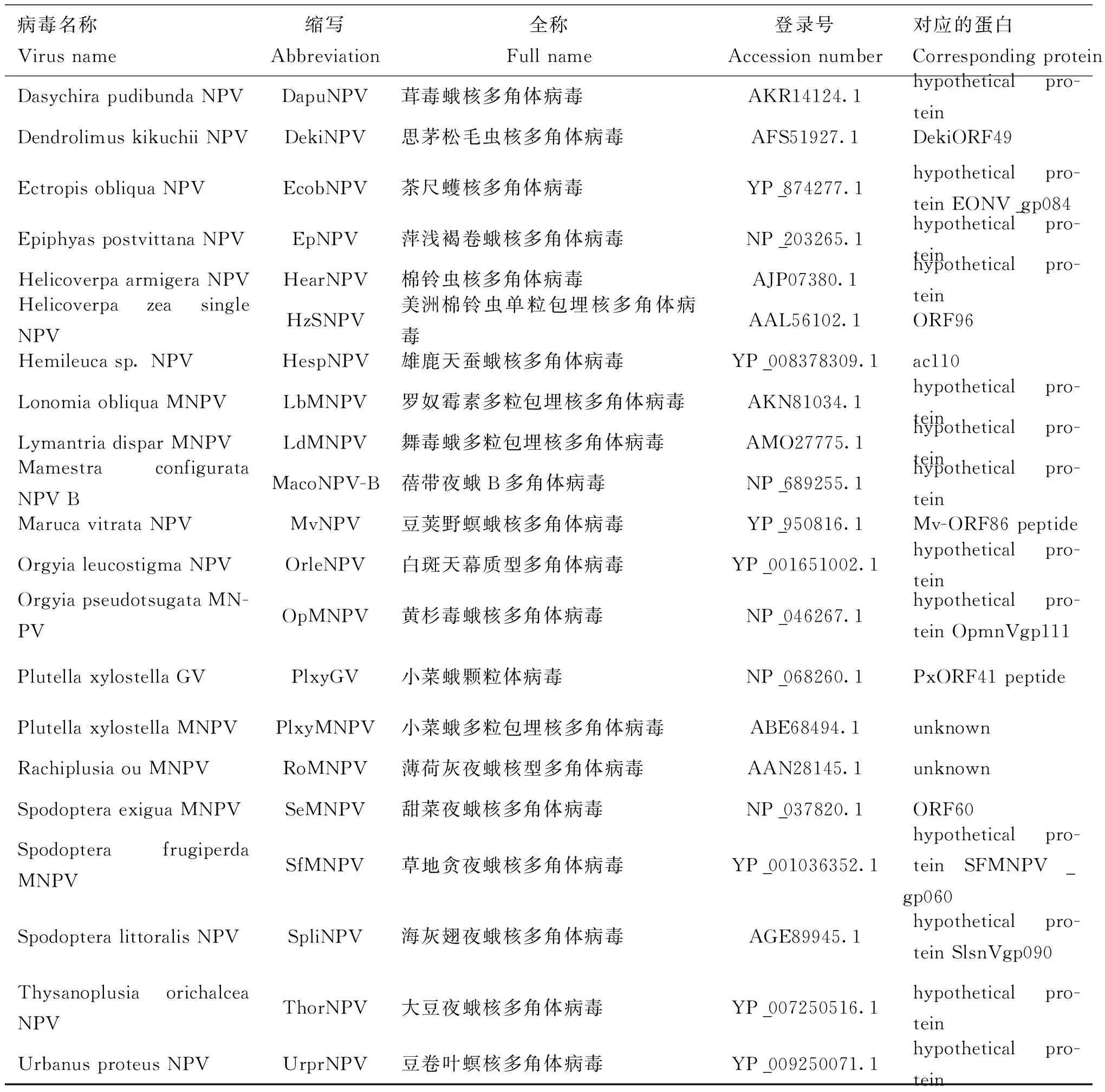
病毒名称Virusname缩写Abbreviation全称Fullname登录号Accessionnumber对应的蛋白CorrespondingproteinDasychirapudibundaNPVDapuNPV茸毒蛾核多角体病毒AKR14124.1hypotheticalpro-teinDendrolimuskikuchiiNPVDekiNPV思茅松毛虫核多角体病毒AFS51927.1DekiORF49EctropisobliquaNPVEcobNPV茶尺蠖核多角体病毒YP_874277.1hypotheticalpro-teinEONV_gp084EpiphyaspostvittanaNPVEpNPV萍浅褐卷蛾核多角体病毒NP_203265.1hypotheticalpro-teinHelicoverpaarmigeraNPVHearNPV棉铃虫核多角体病毒AJP07380.1hypotheticalpro-teinHelicoverpazeasingleNPVHzSNPV美洲棉铃虫单粒包埋核多角体病毒AAL56102.1ORF96Hemileucasp.NPVHespNPV雄鹿天蚕蛾核多角体病毒YP_008378309.1ac110LonomiaobliquaMNPVLbMNPV罗奴霉素多粒包埋核多角体病毒AKN81034.1hypotheticalpro-teinLymantriadisparMNPVLdMNPV舞毒蛾多粒包埋核多角体病毒AMO27775.1hypotheticalpro-teinMamestraconfigurataNPVBMacoNPV-B蓓带夜蛾B多角体病毒NP_689255.1hypotheticalpro-teinMarucavitrataNPVMvNPV豆荚野螟蛾核多角体病毒YP_950816.1Mv-ORF86peptideOrgyialeucostigmaNPVOrleNPV白斑天幕质型多角体病毒YP_001651002.1hypotheticalpro-teinOrgyiapseudotsugataMN-PVOpMNPV黄杉毒蛾核多角体病毒NP_046267.1hypotheticalpro-teinOpmnVgp111PlutellaxylostellaGVPlxyGV小菜蛾颗粒体病毒NP_068260.1PxORF41peptidePlutellaxylostellaMNPVPlxyMNPV小菜蛾多粒包埋核多角体病毒ABE68494.1unknownRachiplusiaouMNPVRoMNPV薄荷灰夜蛾核型多角体病毒AAN28145.1unknownSpodopteraexiguaMNPVSeMNPV甜菜夜蛾核多角体病毒NP_037820.1ORF60SpodopterafrugiperdaMNPVSfMNPV草地贪夜蛾核多角体病毒YP_001036352.1hypotheticalpro-teinSFMNPV_gp060SpodopteralittoralisNPVSpliNPV海灰翅夜蛾核多角体病毒AGE89945.1hypotheticalpro-teinSlsnVgp090ThysanoplusiaorichalceaNPVThorNPV大豆夜蛾核多角体病毒YP_007250516.1hypotheticalpro-teinUrbanusproteusNPVUrprNPV豆卷叶螟核多角体病毒YP_009250071.1hypotheticalpro-tein

图1 Ac110同源蛋白氨基酸序列比对Fig.1 Alignment of Ac110 homologs
2.2 Ac110点突变重组病毒的构建和鉴定
通过比较Ac110同源蛋白的氨基酸序列的同源性,选取2个高度保守的氨基酸位点(图1中黑色箭头所标识的N27和L35位点)进行点突变,分别构建重组病毒vAc110N27A和vAc110L35A。病毒构建示意图如图3所示,其中bMON14272为Bac-to-Bac Expression System中缺失了多角体蛋白基因并能在该基因原位点提供特异性转座的杆状病毒基因组。
2.3 Ac110突变型蛋白的检测
由于重组病毒构建过程中,Ac110蛋白的C末端加上了一个HA标签,因此可用抗HA的标签抗体来检测重组病毒中Ac110HA点突变蛋白是否表达。结果如图3所示,Ac110点突变重组病毒感染的细胞中,能检测到与补回型病毒vAc110HA感染的细胞中大小一样的特异蛋白带,而缺失型重组病毒vAc110KO感染的细胞中则没有这样的蛋白产生,说明Ac110突变体蛋白均能正常表达。同时以β-actin的多抗检测了细胞宿主蛋白β-actin以作为内参。
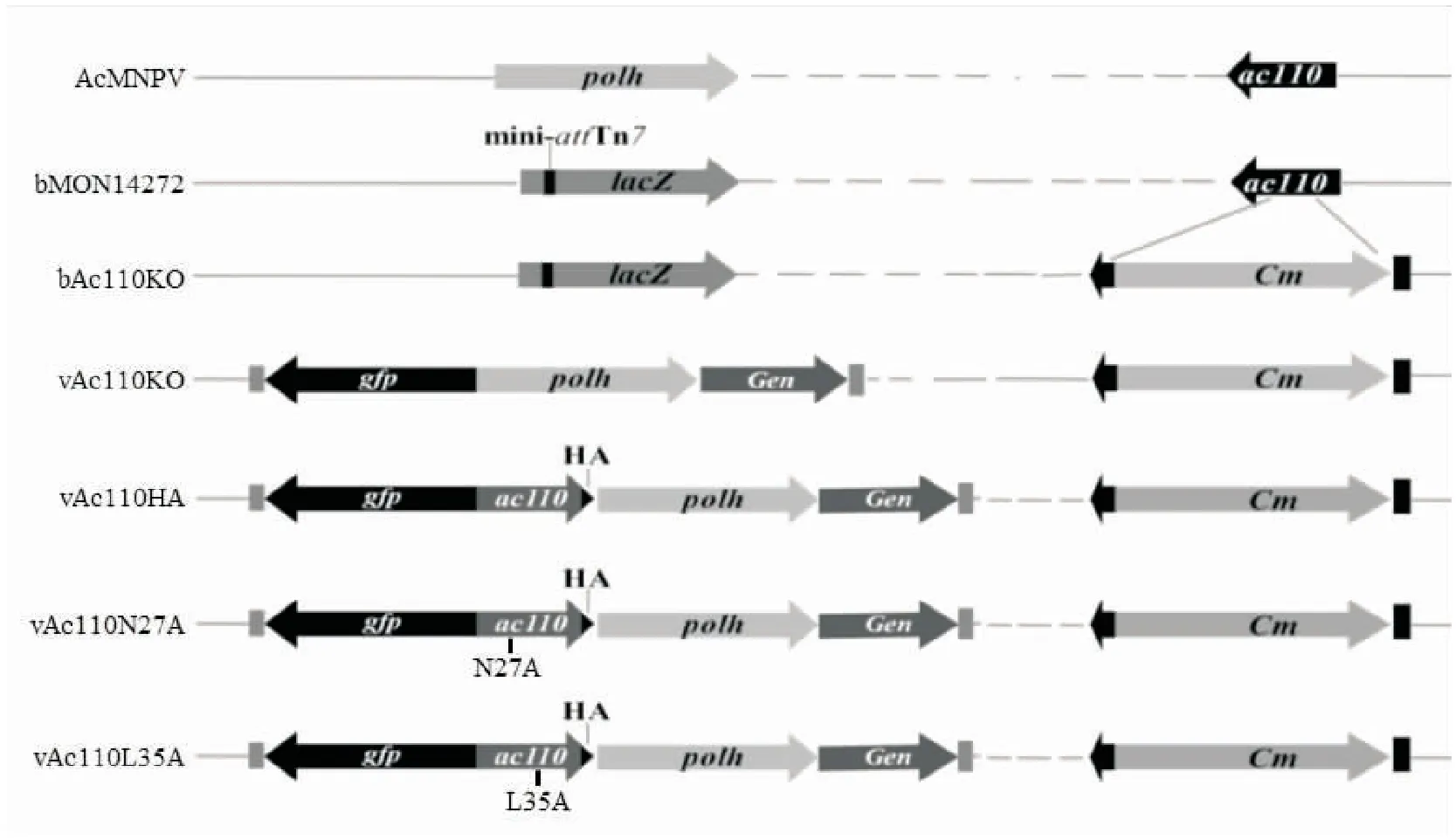
图2 Ac110重组病毒构建示意图Fig.2 The diagram of the construction of Ac110 recombinant viruses AcMNPV polyhedrin (polh) locus was disrupted by lacZ and mini-attTn7 as showed in bMON14272.The ac110 gene was replaced by chloramphenicol resistance gene (Cm) in bAc110KO.Green fluorescent protein gene (gfp) and polh gene were transposed to the polyhedrin locus of bAc110KO to obtain the recombinant virus vAc110KO.The Ac110 with an HA tag at the C terminus was repaired in the vAc110HA.The vAc110N27A and vAc110L35A contained the mutated Ac110 N27A and L35A, respectively.All constructs were confirmed by PCR and sequencing analyses.
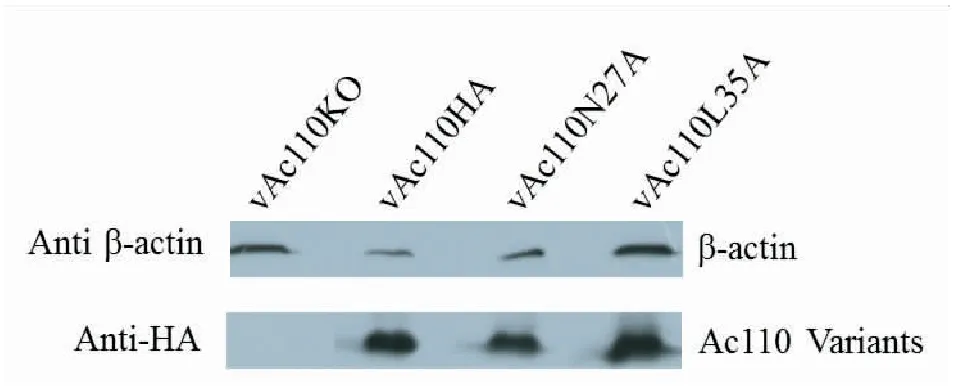
图3 Ac110点突变蛋白表达检测Fig.3 The detection of the Ac110 variants expression by Western blot analysis
2.4 Ac110保守氨基酸的突变不影响病毒在细胞中的正常增殖和扩散
将vAc110N27A和vAc110L35A的bacmid DNA转染至Sf9细胞中,在转染后的不同时间点分别进行荧光显微镜观察和光学显微镜观察,并拍照记录结果。如图4所示,在病毒转染后24 h,不同病毒转染的细胞中,能产生荧光的细胞数量相当,说明病毒DNA的转染效率差不多。而到了病毒转染后72 h,所有细胞都产生了荧光,说明重组病毒均能在细胞中正常复制并扩散。通过光学显微镜可以在白光下看到不同病毒转染的细胞中均有部分细胞已产生了多角体(图4)。收集病毒转染后120 h上清液检测病毒滴度,结果显示Ac110突变型重组病毒的病毒产量和对照组病毒的病毒产量相当(图5A),同时收集该时间点的细胞纯化多角体,通过血球计数板对多角体进行计数,发现突变体与补回型重组病毒相比,多角体产量差别不大(结果未展示)。将重组病毒以低MOI感染Sf9细胞,于不同时间点收取细胞上清液进行病毒滴度测定,绘制病毒生长曲线,发现Ac110突变型重组病毒与补回型重组病毒生长趋势一致。以上结果说明Ac110点突变后不影响病毒在细胞中的增殖和扩散。

图4 重组病毒转染Sf9细胞Fig.4 Transfection of recombinant viruses in Sf9 cellsNote:The bacmid DNA of recombinant viruses were transfected in Sf9 cells and the cells were observed by fluorescent microscopy and light microscopy at indicated time points.

图5 重组病毒在细胞中的复制Fig.5 Replication of recombinant viruses in Sf9 cells注:(A)重组病毒转染Sf9细胞的滴度测定(B)重组病毒低MOI感染Sf9细胞的生长曲线测定。Note: (A) Titres of Ac110 recombinant viruses harvested at 120 h post-transfection.(B) Viral growth curves of Ac110 recombinant viruses.Sf9 cells were infected with recombinant viruses at an MOI of 0.01 TCID50/cell.The supernatants of viruses infected cells were collected at designated time points and the titres were determined by using a TCID50 assay.Each data point represents the average from two independent experiments and error bars represent the standard deviations.Statistical analyses were carried out using GraphPad Prism 6.0 software.
2.5 Ac110蛋白N27位点的突变显著降低了病毒对虫体的口服感染能力
将收集的各个重组病毒多角体(OB)分别稀释至浓度为1×106,1×107和1×108OB/mL,每头三龄期甜菜夜蛾幼虫喂食1 μL OB悬液,即分3种喂食剂量,每头分别喂食1×103,1×104和1×105个OB。每种病毒的每个喂食剂量设置两组实验,每组12头幼虫,另外喂食H2O作为空白对照。统计病毒感染幼虫的死亡率(表2),与补回型重组病毒vAc110HA相比,vAc110L35A用不同剂量感染时,死亡率与vAc110HA基本一致,不存在显著性差异;而vAc110N27A感染的幼虫死亡率会下降约20%,感染剂量为1×105和1×104OB时,二者之间存在显著性差异,但剂量为1×103OB时不存在显著性差异(图6)。
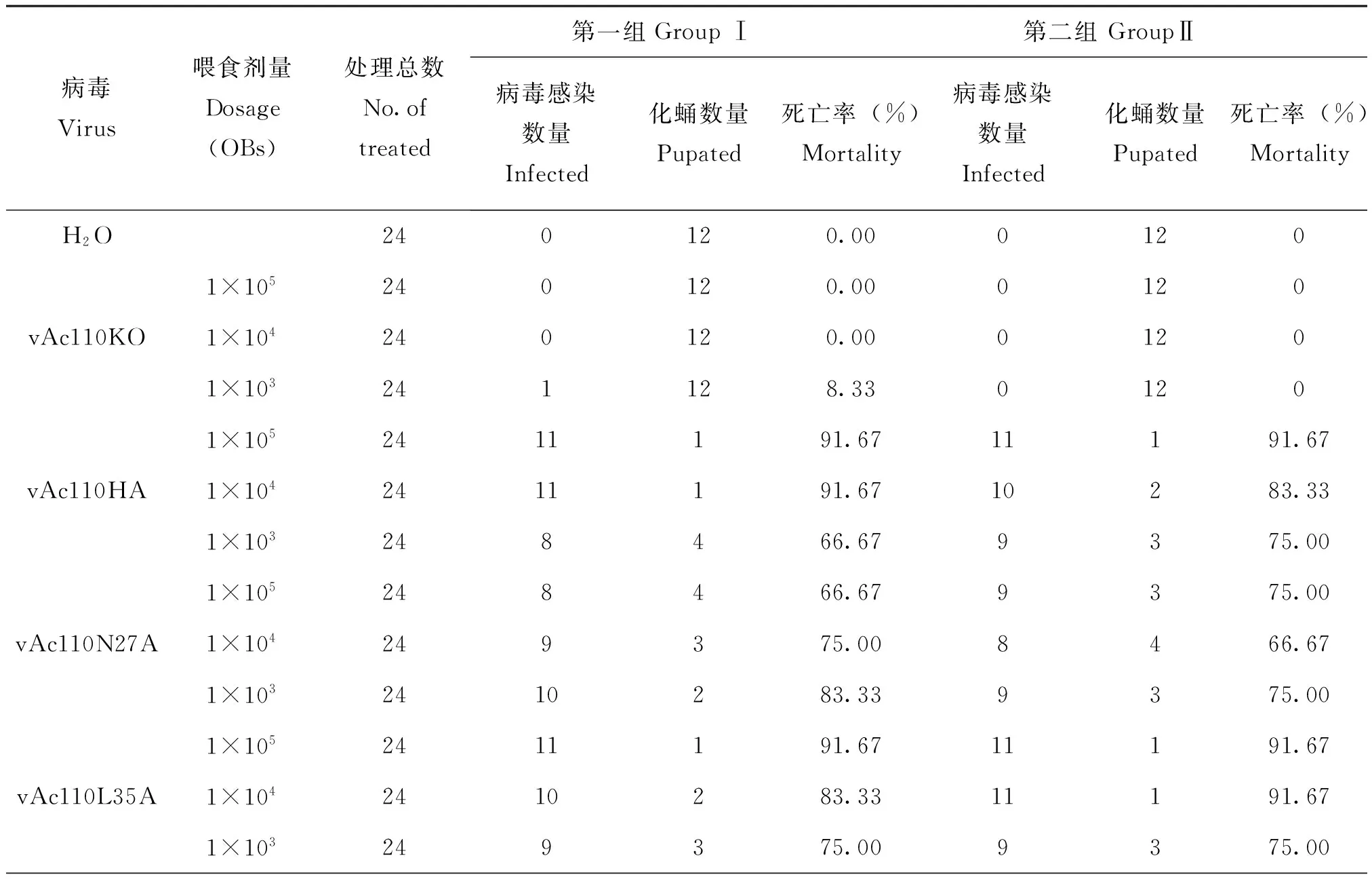
表2 重组病毒生物测定Table 2 Bioassays of recombinant viruses
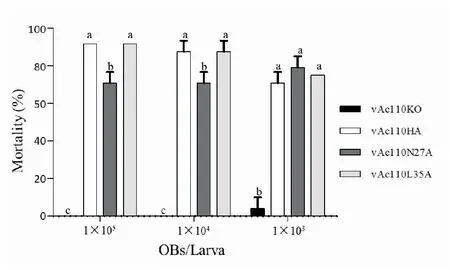
图6 重组病毒生物测定分析Fig.6 Mortality analyses of bioassays with recombinant viruses
3 结论与讨论
本实验室前期对杆状病毒Ac110的功能研究中,发现Ac110是一个新的口服感染因子PIF,其缺失对病毒在细胞中的增殖和扩散不受影响,但是病毒却不能通过口服感染昆虫幼虫,并且昆虫的围食膜PM不是Ac110的作用靶标(Liuetal., 2016)。为了进一步揭示Ac110在口服感染过程中的作用机制,本文选取了Ac110蛋白两个高度保守的氨基酸位点N27和L35,构建了两株Ac110点突变重组病毒,展开了相关的实验研究。结果显示这两个氨基酸位点的突变均不影响病毒在细胞的增殖和扩散,并且在不同的病毒多角体的感染剂量下,L35位点的突变不影响病毒对昆虫幼虫的口服感染,但是在每头虫子感染1×105和1×104OB时,N27位点的突变却显著降低了病毒对昆虫幼虫的口服感染能力,说明N27位点对Ac110的功能起着重要的作用。
近年来对PIF的功能研究以及ODV如何进入中肠上皮细胞一直是研究的热点。通过电镜观察,有研究表明ODV在感染昆虫中肠的过程中,会与中肠上皮细胞微绒毛结合和融合(Kawanishietal., 1972),但对这过程中的分子机制了解不多。在目前已经研究发现的众多的PIF中,P74、PIF-1和PIF-2介导了病毒与细胞的结合,缺失三者中的任何一个基因都会造成ODV与微绒毛的结合能力下降(Haas-Stapletonetal., 2004; Ohkawaetal., 2005)。而PIF-3和PIF-5则不参与ODV与中肠上皮细胞的结合和融合,可能参与了ODV与细胞结合后的一些关键过程,比如衣壳的转运,细胞信号转导等(Ohkawaetal., 2005; Sparksetal., 2011)。PIF-1、PIF-2和PIF-3能在ODV的表面形成一个高度稳定的复合体,PIF-4与该复合体有强的相互作用,P74和PIF-8则与该复合体有较弱的相互作用,而PIF-5则与该复合体完全没有相互作用(Pengetal., 2012)。另外,P74与PIF-5能分别与宿主中肠蛋白中大小为35 kDa和97 kDa的两个蛋白产生相互作用,但是这两个宿主蛋白到底是什么却仍旧未知(Sparksetal., 2011; Zhouetal., 2005)。上述的8个PIF以及本实验室所发现的Ac110都是由杆状病毒核心基因所编码的,表明病毒对昆虫的口服感染机制是古老而又高度保守的。然而也有研究表明,非杆状病毒核心基因所编码的SF58,也参与到病毒对虫体的口服感染过程中(Simonetal., 2012),暗示了口服感染机制比预想中更要复杂,可能由于病毒与宿主的协同进化,导致这一过程与病毒的种属特异性有关。
目前Ac110的作用机制未明,Ac110是否通过与PIF复合体结合,或者通过与宿主蛋白相互作用而行使功能,需要进一步的实验验证。而Ac110保守氨基酸位点N27的突变影响了病毒的口服感染能力,说明这个位点对于Ac110蛋白有效发挥功能是至关重要的。该位点的突变可能影响了Ac110蛋白的结构从而影响了Ac110与其他蛋白的相互作用网络,也可能是Ac110直接的功能位点,其中的分子机理有待深入的研究。
References)
Braunagel SC, Russell WK, Rosas-Acosta G,etal.Determination of the protein composition of the occlusion-derived virus of Autographa californica nucleopolyhedrovirus[J].ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica, 2003, 100 (17):9797-9802.
Chiu J, March PE, Lee R,etal.Site-directed, Ligase-Independent Mutagenesis (SLIM): A single-tube methodology approaching 100% efficiency in 4 h[J].NucleicAcidsResearch, 2004, 32 (21):e174.
Fang M, Nie Y, Harris S,etal.Autographacalifornicamultiple nucleopolyhedrovirus core gene ac96 encodes a per Os infectivity factor (PIF-4)[J].JournalofVirology, 2009, 83 (23):12569-12578.
Fang M, Nie Y, Wang Q,etal.Open reading frame 132 ofHelicoverpaarmigeranucleopolyhedrovirus encodes a functional per os infectivity factor (PIF-2)[J].TheJournalofGeneralVirology, 2006, 87 (Pt 9):2563-2569.
Faulkner P, Kuzio J, Williams GV,etal.Analysis of p74, a PDV envelope protein of Autographa californica nucleopolyhedrovirus required for occlusion body infectivityinvivo[J].TheJournalofGeneralVirology, 1997, 78 ( Pt 12):3091-3100.
Garavaglia MJ, Miele SA, Iserte JA,etal.The ac53, ac78, ac101, and ac103 genes are newly discovered core genes in the family Baculoviridae[J].JournalofVirology, 2012, 86 (22):12069-12079.
Haas-Stapleton EJ, Washburn JO, Volkman LE.P74 mediates specific binding of Autographa californica M nucleopolyhedrovirus occlusion-derived virus to primary cellular targets in the midgut epithelia ofHeliothisvirescenslarvae[J].JournalofVirology, 2004, 78 (13):6786-6791.
Harrison RL, Sparks WO, Bonning BC.Autographa californica multiple nucleopolyhedrovirus ODV-E56 envelope protein is required for oral infectivity and can be substituted functionally by Rachiplusia ou multiple nucleopolyhedrovirus ODV-E56[J].TheJournalofGeneralVirology, 2010, 91 (Pt 5):1173-1182.
Hegedus D, Erlandson M, Gillott C,etal.New insights into peritrophic matrix synthesis, architecture, and function[J].AnnualReviewofEntomology, 2009, 54:285-302.
Ishimwe E, Hodgson JJ, Clem RJ,etal.Reaching the melting point: Degradative enzymes and protease inhibitors involved in baculovirus infection and dissemination[J].Virology, 2015, 479-480c:637-649.
Javed MA, Biswas S, Willis LG,etal.Autographa californica multiple nucleopolyhedrovirus AC83 is a Per Os Infectivity Factor (PIF) protein required for Occlusion-Derived Virus (ODV) and budded virus nucleocapsid assembly as well as assembly of the PIF complex in ODV eEnvelopes[J].JournalofVirology, 2017, 91 (5).
Kawanishi CY, Summers MD, Stoltz DB,etal.Entry of an insect virus in vivo by fusion of viral envelope and microvillus membrane[J].JournalofInvertebratePathology, 1972, 20 (1):104-108.
Kikhno , Gutierrez S, Croizier L,etal.Characterization of pif, a gene required for the per os infectivity of Spodoptera littoralis nucleopolyhedrovirus[J].TheJournalofGeneralVirology, 2002, 83 (Pt 12):3013-3022.
Lepore LS, Roelvink PR, Granados RR.Enhancin, the granulosis virus protein that facilitates nucleopolyhedrovirus (NPV) infections, is a metalloprotease[J].JournalofInvertebratePathology, 1996, 68 (2):131-140.
Liu J, Zhu L, Zhang S,etal.The Autographa californica multiple nucleopolyhedrovirus ac110 gene encodes a new per os infectivity factor[J].VirusResearch, 2016, 221:30-37.
Nie Y, Fang M, Erlandson MA,etal.Analysis of the autographa californica multiple nucleopolyhedrovirus overlapping gene pair lef3 and ac68 reveals that AC68 is a per os infectivity factor and that LEF3 is critical, but not essential, for virus replication[J].JournalofVirology, 2012, 86 (7):3985-3994.
O’Reilly DR, Miller L K, Luckow VA.Baculovirus expression vector: a laboratory manual.New York, USA: Freeman, W.H.and Co, 1992.
Ohkawa T, Washburn JO, Sitapara R,etal.Specific binding of Autographa californica M nucleopolyhedrovirus occlusion-derived virus to midgut cells of Heliothis virescens larvae is mediated by products of pif genes Ac119 and Ac022 but not by Ac115[J].JournalofVirology, 2005, 79 (24):15258-15264.
Peng K, van Lent JW, Boeren S,etal.Characterization of novel components of the baculovirus per os infectivity factor complex[J].JournalofVirology, 2012, 86 (9):4981-4988.
Pijlman GP, Pruijssers AJ, Vlak JM.Identification of pif-2, a third conserved baculovirus gene required for per os infection of insects[J].TheJournalofGeneralVirology, 2003, 84 (Pt 8):2041-2049.
Simon O, Palma L, Williams T,etal.Analysis of a naturally-occurring deletion mutant ofSpodopterafrugiperdamultiple nucleopolyhedrovirus reveals sf58 as a new per os infectivity factor of lepidopteran-infecting baculoviruses[J].JournalofInvertebratePathology, 2012, 109 (1):117-126.
Sparks WO, Harrison RL, Bonning BC.Autographa californica multiple nucleopolyhedrovirus ODV-E56 is a per os infectivity factor, but is not essential for binding and fusion of occlusion-derived virus to the host midgut[J].Virology, 2011, 409 (1):69-76.
Wang P, Granados RR.Calcofluor disrupts the midgut defense system in insects[J].InsectBiochemistryandMolecularBiology, 2000, 30 (2):135-143.
Wu W, Liang H, Kan J,etal.Autographa californica multiple nucleopolyhedrovirus 38K is a novel nucleocapsid protein that interacts with VP1054, VP39, VP80, and itself[J]..JournalofVirology, 2008, 82 (24):12356-12364.
Wu W, Lin T, Pan L,etal.Autographa californica multiple nucleopolyhedrovirus nucleocapsid assembly is interrupted upon deletion of the 38K Gene[J].JournalofVirology, 2006, 80 (23):11475-11485.
Xiang X, Chen L, Guo A,etal.The Bombyx mori nucleopolyhedrovirus (BmNPV) ODV-E56 envelope protein is also a per os infectivity factor[J].VirusResearch, 2011, 155 (1):69-75.
Zhou W, Yao L, Xu H,etal.The function of envelope protein P74 fromAutographacalifornicamultiple nucleopolyhedrovirus in primary infection to host[J].VirusGenes, 2005, 30 (2):139-150.
Zhu S, Wang W, Wang Y,etal.The baculovirus core gene ac83 is required for nucleocapsid assembly and per os infectivity of Autographa californica nucleopolyhedrovirus[J].JournalofVirology, 2013, 87 (19):10573-10586.
The conserved amino acid N27 of baculovirus Ac110 is important for efficient establishment of midgut infection during oral infection
ZHU Le-yuan1, LIU Jian-tao1,2, WANG Yan-ling1, YUAN Mei-jin1, WU Wen-bi1, YANG Kai1
(1.State Key Laboratory of Biocontrol, Sun Yat-sen University, Guangzhou 510275, China;2.Jiangxi Key Laboratory of Bioprocess, Jiangxi Science & Technology Normal University, Nanchang, Jiangxi 330013, China)
The family Baculoviridae is a highly selective pathogen in arthropods, mainly in insects of the order Lepidoptera.Ac110 is one of theperosinfectivityfactorsthat are essential for the initial infection of lavae in midgut cells, but the mechanism by which Ac110 employs that affect the midgut infection is still unknown.In the present study, two recombinant viruses, vAc110N27A and vAc110L35A were generated in which the conserved amino acids N27 and L35 of Ac110 were mutated to alanine, respectively.The expression of the mutant protein were confirmed by Western blot analysis.Transfection and infection assays and viral growth curves analysis were done by using Sf9 cells, the results showed that both the recombinant viruses were able to produce compatible infectious budded viruses and the spread of the viruses among Sf9 cells was not affected compared with the control virus.Bioassays were performed by feedingSpodopteraexigualarvae with purified polyhedra to detect the ability to initiate midgut infection for both recombinant viruses.The mortality rate of vAc110N27A was significantly reduced compared to the control virus, while no difference was observed for vAc110L35A.Thus we conclude that N27 of Ac110 is important for the function of Ac110 duringperosinfection.
Baculovirus;perosinfectivityfactor; Ac110; Bioassay
朱乐园,刘建涛,王燕玲,等.杆状病毒Ac110蛋白的保守氨基酸位点N27对病毒在中肠建立有效感染起重要作用[J].环境昆虫学报,2017,39(3):505-514.
中央高校基本科研业务费专项资金(15lgjc20)
朱乐园,女,1994年生,硕士,研究方向为杆状病毒功能基因,E-mail:zhuly23@mail2.sysu.edu.cn
*通讯作者 Author for correspondence, E-mail: wuwenbi3@mail.sysu.edu.cn
Received:2017-05-06; 接受日期Accepted: 2017-05-28
Q968.1;S433.4
A
1674-0858(2017)03-0505-10
猜你喜欢
山东畜牧兽医(2022年10期)2022-12-06
中国生物防治学报(2022年3期)2022-07-09
保健文汇(2022年1期)2022-06-06
当代水产(2021年7期)2021-11-04
放射学实践(2020年2期)2020-03-13
中国生殖健康(2019年9期)2019-01-07
中国生殖健康(2019年7期)2019-01-06
河南农业科学(2017年8期)2017-09-01
江苏农业科学(2016年11期)2017-03-21
中国兽医杂志(2016年5期)2016-06-27
