
龙须菜配子体发育和不同品系四分孢子体四分孢子形成过程显微结构观察
2017-08-10 12:17米萍隋正红阙州周伟商二磊
海洋学报 2017年8期
米萍,隋正红*,阙州,周伟,商二磊
(1.中国海洋大学 海洋生物遗传与育种教育部重点实验室,山东 青岛 266003)
龙须菜配子体发育和不同品系四分孢子体四分孢子形成过程显微结构观察
米萍1,隋正红1*,阙州1,周伟1,商二磊1
(1.中国海洋大学 海洋生物遗传与育种教育部重点实验室,山东 青岛 266003)
龙须菜配子体和四分孢子体在生殖结构的形成和发育的过程中,出现一系列的细胞形态和结构层面的变化。本实验以龙须菜雌雄配子体和不同品系龙须菜四分孢子体为实验材料,采用连续徒手切片的方法,观察龙须菜配子体发育和不同品系四分孢子体四分孢子形成过程。观察到,雌雄配子体发育经过幼苗期、毛细胞期、性成熟期,幼苗期和毛细胞期雌雄配子体间无明显差异,进入性成熟期后,产生生殖结构时才能将其区分开。四分孢子起源于四分孢子体表皮层细胞,初始为鲜红色直径约5~10 μm的细胞,逐渐生长形成成熟的直径约20~25 μm的四分孢子囊,并分裂为4个四分孢子聚集在四分孢子囊中,在藻体外,四分孢子释放变形成球形,龙须菜四分孢子的形成和发育过程伴随着髓部细胞中红藻淀粉颗粒的减少。其中“981”龙须菜形成较多的畸形四分孢子,可能是其表现低育的一个原因。龙须菜自基部到尖端的表皮层细胞层数、髓部细胞数目和体积均发生连续的变化。细胞连接广泛存在于龙须菜表皮层、皮层和髓部细胞之间,但各层细胞连接大小、数目、长度等都存在差异。释放的四分孢子可能会附着在四分孢子体上,固着器覆盖包绕四分孢子体甚至侵入到破损有伤口的四分孢子体内,形成四分孢子体和配子体在同一株上的世代混杂现象,这也可能是导致龙须菜表现遗传复杂性的原因。
龙须菜;配子体;四分孢子体;生殖结构;显微观察
1 引言
龙须菜(Gracilariopsislemaneiformis)隶属于红藻门(Rhodophyta)、红藻纲(Rhodophyceae)、真红藻亚纲(Florideae)、杉藻目(Gigartinales)、江蓠科(Gracilariaceae),是一种重要的大型海洋经济红藻。龙须菜是生产琼胶的重要原料[1—2],同时也是鲍鱼养殖的饲料[3—4],另外龙须菜还有食用[5]和净化水体[6—7]等作用。
1998年选育出的有耐高温、速生、抗逆、琼胶含量高等优良特性的“981”龙须菜新品种[8],奠定了龙须菜栽培的基础,随后在2007年选育出“2007”良种[9],2014年培育出“鲁龙1号”[10],使得龙须菜栽培养殖得到更大规模的推广。据2013年中国渔业统计年鉴报道,以龙须菜为代表的江蓠栽培成为我国第二大海藻栽培产业。
龙须菜是典型的三世代交替(或称多管藻)型生活史,包括二倍体的四分孢子体、附生于雌配子体藻枝上的果孢子体世代和单倍体的配子体世代[11]。从20世纪40年代起,对江蓠科精子囊类型有详细的报道[12],对精子囊和果胞的分布组成和形态结构,也有了深入研究[13—14],较为全面地了解了配子体生殖结构的特征和细胞组成,而配子体性成熟前期的发育过程未见报道,也缺乏对配子体生长发育阶段的连续观察,另外雌雄配子体在形态上相似,配子体性成熟前难于从形态上加以区分,对配子体发育阶段的典型过程缺乏判断,也限制了杂交育种工作的进程。“981”龙须菜是四分孢子体,其在生长过程中表现出不同的发育特性,即四分孢子的数量低于野生型,表现低育晚育的特性[15]。对于龙须菜四分孢子体的研究,局限于四分孢子在藻体表面的存在状态及四分孢子附着生长为配子体幼苗过程的研究[16],而对四分孢子形成和释放过程缺乏研究报道。“981”龙须菜发育特性的不同使之成为揭示龙须菜四分孢子体发育控制与变化的难得材料,另外在产业上具有低育晚育特性的品系(种)有更重要的应用价值,与其高产的生产性状可能存在遗传和生长发育层面的联系。
本实验进行了龙须菜配子体发育和不同品系四分孢子体四分孢子形成过程显微结构观察,一方面可以完善配子体生长发育的过程,寻找性成熟前期雌雄配子体间的差异,促进杂交育种进程;另一方面找到“981”龙须菜的细胞结构和显微观察层面的生长特性,阐释四分孢子体发育控制机理。
2 材料和方法
2.1 实验材料
本实验的材料包括龙须菜雌雄配子体,及“981”龙须菜、“鲁龙1号”两个栽培良种和龙须菜四分孢子体野生品系。
配子体来源:实验室条件下收集2013年12月份在湛山湾采集的野生四分孢子体放散出的四分孢子培养长成的配子体。
四分孢子体材料来源:“981”龙须菜和“鲁龙1号” 两个栽培良种来自福建莆田龙须菜栽培海区和青岛胶州湾龙须菜栽培海区,采集时间为2015年6月至2015年9月;野生龙须菜四分孢子体采自青岛湛山湾,采集时间为2015年4月至2015年10月。
2.2 实验材料的处理和培养
实验材料采集后,先用海水冲洗3遍后,再用灭菌海水冲洗并用毛刷刷掉表面其他不易冲洗掉的杂藻和杂质等。其中,2013年12月份采集的野生四分孢子体,实验室条件下收集放散的四分孢子,待四分孢子长成配子体幼苗后单株培养30株。培养条件:温度23℃, 光照40 μmol/(m2·s), 光周期12L/12D。
2015年采集的“981”龙须菜、“鲁龙1号”和野生四分孢子体,用于四分孢子体四分孢子形成观察,培养条件:(20±1)℃,光照30 μmol/(m2·s), 12L/12D条件下培养,每3天换一次加入Pro培养基的新鲜灭菌海水。
2.3 实验材料的观察
2.3.1 野生龙须菜配子体观察
根据Zhou等[17]开发的龙须菜性别相关SCAR标记,按照相应的实验步骤对30株龙须菜配子体幼苗进行雌雄鉴定,根据鉴定结果选取雌雄配子体各3株,连续徒手切片在显微镜(Nikon ECLIPSE 80i,日本)下观察雌雄配子体生殖结构的形成和发育过程。
2.3.2 野生龙须菜四分孢子体观察
实验材料在显微镜(Nikon ECLIPSE 80i,日本)下观察,判断藻体状态和初步判断生长发育阶段。
采集到的龙须菜在显微镜下镜检观察,不同采集时间的龙须菜生长状态和发育阶段不同,挑选出同一时间采集的“981”龙须菜和“鲁龙1号”未成熟材料各3株,在实验室条件下培养。同时挑选同一时间段采集的未成熟野生龙须菜,从外观形态上挑选藻体表面无囊果、藻枝较粗壮的未成熟的野生龙须菜,每3天进行一次观察,连续观察藻体表面细胞状态和变化,并进行徒手切片,观察藻体横切面上表皮、皮层和髓部细胞的结构和变化及四分孢子的形成和发育过程。
在实验室条件下培养,培养一段时间后出现四分孢子囊原始细胞并形成四分孢子囊的藻株即为野生龙须菜四分孢子体。对鉴定确认为野生四分孢子体的藻株连续观察。
2.4 龙须菜不同部位和不同层次细胞特征及细胞连接
雌雄配子体和野生四分孢子体从靠近固着器的基部、中部、藻尖进行连续切片,观察不同部位细胞特征。
采用高碘酸-Schiff试剂染色(PAS)和考马斯亮蓝-R250(CBB)染色[18—19]。其中PAS染色将多糖类染为红色,CBB染色将蛋白质成分染为蓝色。将材料用中性甲醛固定后,用PAS染色和CBB染色,观察细胞中多糖和蛋白质的组成和相关细胞结构。
3 结果
3.1 龙须菜配子体生长发育过程观察
龙须菜雌雄配子体生长首先经历幼苗期,藻体只有一层表皮细胞,随后在表层细胞中会出现与表皮细胞大小相近颜色偏绿的细胞(图1 a),细胞不断生长变大,产生大液泡(图1 b)并发生分裂形成一个上位产毛细胞和1个下位细胞(图1 c),产毛细胞上端生长突出浅色毛状结构,毛状结构能不断生长甚至大于毛细胞(图1 d~f),毛细胞出现阶段藻体生长快速。在幼苗期和毛细胞期雌雄配子体形态和发育过程无明显差异。
藻体进入生殖成熟期后,雄配子体表皮细胞通过一次斜纵分裂产生两个精子囊母细胞(图1 g);精子囊母细胞再通过一次横分裂产生一个上位的精子囊细胞(spermatangium)和一个下位的较小精子囊母细胞(图1 h);精子囊细胞释放不动精子后,颜色变浅(图1 i)。精子细胞直径约3~5 μm,较周围表皮细胞(直径约10~15 μm)小(图1 j)。

图1 龙须菜雌雄配子体毛细胞发育过程和生殖结构的形成Fig.1 Development process of hair cell and reproductive apparatus in the male and female gametophyte of G. lemaneiformis. a. 初始毛细胞(箭头所示),b. 毛细胞增大,出现大液泡(箭头所示),c. 毛细胞分裂产生产毛细胞(箭头所示)和下位细胞(无柄箭头所示),d~f. 突出的毛结构生长(箭头所示);g~j. 雄配子体精子囊发育,g. 初始精母细胞进行一次斜纵分裂产生2个精母细胞(箭头所示),h. 精母细胞进行一次横分裂产生1个靠上的精细胞和1个靠下的小精母细胞(箭头所示),i. 精细胞(箭头所示)成熟颜色变浅,j. 精母细胞(sp)较表皮细胞(ep)小; k~l. 雌配子体生殖结构发育,k.支持细胞(su)和上方的原始果胞细胞(cp),l. 原始果胞细胞横分裂形成1个上方的果胞(cp)和1个下方的下位细胞(hy), 两边分布着2~3个营养细胞枝(sb).l、w表示长度、宽度,单位μma. The primary hair cell(see arrow), b. The hair cell increased in volume and formed a big vacuole(see arrow), c. The hair cell divided into an upper hair produced cell(see arrow) and a lower cell(see arrowhead), d-f. The growth of protruded hair structure(see arrow);g-j. Development of male apparatus, g. The primary spermatangium mother cell splits oblique-longitudinally to produce two spermatangium mother cells(see arrow), h. The spermatangium mother cell divided transversely to produce an upper spermatangium and a lower small spermatangium mother cell(see arrow), i The spermatangium turned white after the sperms were released,j. The spermatangium was smaller than the epidermal cell. k-l. Development of female apparatus,k. The supporting cell(su) and an upper carpogonial branch initial(cp), l. The carpogonial branch initial splited into an upper carpogonium(cp) and a lower hypogynous cell(hy), 2-3 cells sterile branches distribute on the both side. l:length, w:width,unit:μm
雌配子体表皮层中,支持细胞通过分裂产生一个上方的原始果胞细胞(cp)和下方的支持细胞(su)(图1 k),原始果胞细胞两边存在两个营养细胞,随后,原始果胞细胞通过分裂产生2~3个细胞组成的营养细胞枝(sb)(图1l),成熟的雌性生殖结构由果胞(cp)、下位细胞(hy)、支持细胞(su)(图1l)及果胞两边2~3个细胞的营养细胞枝组成,并最终通过受精丝与精子结合。

图2 不同品系龙须菜四分孢子形成过程中表皮层和髓部细胞的变化Fig.2 The change of the epidermal cells and medullary cells in different strains of tetrasporophyte of G. lemaneiformisa1~a6,c1~c6,e1~e6.分别为野生龙须菜,“鲁龙1号”,“981”龙须菜四分孢子形成过程;b1~b6,d1~d6,f1~f6.分别为野生龙须菜四分孢子体,“鲁龙1号”,“981”龙须菜四分孢子形成过程中对应髓部细胞的变化a1-a6,c1-c6,e1-e6. Development of tetrasporgonia in tetrasporophyte of wild type, cultivar “Lulong NO.1”, and cultivar 981 respectively; b1-b6,d1-d6,f1-f6. The change of medullary cells during the development of tetraspogonia in tetrasporophyte of wild type, cultivar “Lulong NO.1”, and cultivar 981 respectively
3.2 龙须菜四分孢子体生长发育过程观察
观察到包括四分孢子囊的形成过程和发育过程,及四分孢子的放散过程。
3.2.1 龙须菜四分孢子囊形成过程中表皮层、皮层和髓部细胞的形态变化
在相同培养条件下,龙须菜初期表皮层细胞颜色大小均一(图2 a1,c1,e1),野生龙须菜四分孢子体和“鲁龙1号”在实验室培养1个月左右藻体表皮中出现鲜红色的四分孢子囊母细胞;但“981”龙须菜较难形成四分孢子囊,相同培养条件下,培养2个月左右藻体表面才出现鲜红色四分孢子囊母细胞。
形成四分孢子囊母细胞后,四分孢子囊的生长发育可以分为:四分孢子囊母细胞出现和细胞生长期,该时期在表皮细胞中出现颜色鲜红、直径在5~10 μm大小与表皮层细胞大小相似的细胞,即为四分孢子囊母细胞(图2 a2,c2,e2箭头所示),四分孢子囊母细胞大量出现,对应的髓部细胞中含有大量的红藻淀粉颗粒(图2 b2,d2,f2),随后细胞生长体积变大,细胞直径达到15~25 μm(图2 a3,c3,e3),对应的髓部细胞中红藻淀粉颗粒减少(图2 b3,d3,f3),时间持续1周左右;接着进入四分孢子囊分裂期,该时期四分孢子囊直径增大到15~25 μm,在藻体表面能观察到四分孢子囊中间的分裂沟(图2 a4,c4 箭头所示),对应的髓部细胞中红藻淀粉颗粒进一步减少并且髓部细胞边缘出现不规则的褶皱状突起(图2 b4,d4,f4),此时四分孢子囊母细胞已经进行了2次分裂,形成4个四分孢子母细胞;四分孢子放散期,这一时期四分孢子囊从藻体上脱落,在藻体外四分孢子囊内的四分孢子母细胞变形释放出球形四分孢子,同时在藻体表面留下孔洞(图2 a5,a6,c5,c6,e5,e6)。对应的髓部细胞中红藻淀粉颗粒进一步减少到几乎不可见的程度并且髓部细胞边缘出现不规则的褶皱状突起更加明显(图2 b5,d5,f5),放散完全后,髓部细胞不规则突起程度减缓(图2 b6,d6,f6)。
3.2.2 横切面上龙须菜四分孢子形成及释放过程
四分孢子囊母细胞出现和生长期,该时期四分孢子囊母细胞不断生长,体积变大,横切面上观察,四分孢子囊母细胞从直径(5~10) μm×(10~15) μm左右逐渐长大到(20~25) μm×(40~45) μm左右的椭球形细胞(图3 a1~a3,b1~b3,c1~c3 箭头所示)。同时周边表皮细胞随着四分孢子囊母细胞的生长逐渐变形,细胞变细长,由两层表层细胞相连接,形成围绕四分孢子囊母细胞的一个囊窝状结构(图3 a3,b5 无柄箭头所示)。在藻段表面观,能观察到四分孢子囊母细胞体积进一步扩大,直径从5~10 μm增大到15~25 μm,颜色呈现均匀分布的鲜红色,而周围表皮细胞色素颜色较浅,围绕细胞外周,中间颜色浅(图2 a,c,e)。
四分孢子囊母细胞分裂期,四分孢子囊母细胞生长到大约(20~25)μm×(40~50)μm的椭球形细胞,细胞生长到这个阶段后四分孢子囊母细胞开始分裂,第一次分裂为横分裂,分裂面垂直横切面半径方向,细胞一分为二;第二次分裂的分裂面沿着横切面的半径方向,但是第一次分裂形成的上下2个细胞与第二次分裂时的分裂面大部分并不在同一平面,2个分裂面呈一定角度(0°~90°),因此在横切面上看到较多的是四分孢子囊中的3个细胞,呈现“T”形分布(图4 a,b),少数情况下能看到第二次分裂上下2个细胞分裂面重合,横切面上能观察到4个细胞呈现“十”字形排列(图4c)。形成的4个四分孢子原始细胞在藻体上并没有形成细胞壁及4个完全分离的四分孢子,而是4个四分孢子原始细胞聚集在一起。此时在藻体表面上观察到四分孢子囊直径约15~20 μm,同时能明显观察到细胞分裂为2个细胞(图2 a4,c4,e4)。
四分孢子释放期,四分孢子囊母细胞在藻体上进行两次减数分裂形成聚集在一起的一个由4个四分孢子原始细胞组成的四分孢子囊,整个四分孢子囊从藻体上脱离,游离到藻体外,在藻体表面留下孔洞(图3 a5,a6,b5,b6,c5箭头所示)。游离出的四分孢子囊,一般在10 min内完成四分孢子的释放过程,四分孢子的释放过程,由原来分裂形成的四分孢子原始细胞,发生原生质体流动及细胞变形,从四分孢子囊中脱离,形成球形四分孢子(图5)。
3.2.3 不同品系龙须菜在四分孢子形成和发育过程中的差异
野生龙须菜四分孢子体、“鲁龙1号”、“981”3种品系龙须菜在四分孢子形成过程中,观察到野生龙须菜四分孢子体和“鲁龙1号”四分孢子形成和发育过程与“981”龙须菜存在差异。
“981”龙须菜在形成四分孢子囊和四分孢子释放过程中,与“鲁龙1号”和野生四分孢子体相比:
(1)出现较多畸形发育的四分孢子囊,四分孢子囊发育不完全、细胞发生变形、颜色变为粉红色(图6a~f)。
(2)从藻体脱离的四分孢子囊,很大一部分不能正常释放形成4个正常的四分孢子。一部分游离出的四分孢子囊,伴随着色素的消失发生破裂溶解(图6g~l);一部分释放出的四分孢子很快破裂溶解,仅剩1~3个正常四分孢子(直径20~25 μm)(图6 m,n);一部分释放出4个四分孢子大小存在明显差异,直径从5 ~25 μm不等,形成的较小的四分孢子会很快破裂内容物流出(图6 o,p);一部分游离的四分孢子囊长时间(小于等于2 h)内,形态上无明显变化,四分孢子不能正常释放(图7 f);另外还有一部分释放4个正常四分孢子。

图3 不同品系龙须菜四分孢子体四分孢子囊的发育和释放过程Fig.3 Development of tetraspogonium in different strains of tetrasporophyte of G. lemaneiformisa1~a6,b1~b6,c1~c6. 分别是野生龙须菜四分孢子体,“鲁龙1号”,“981”龙须菜四分孢子囊形成和释放过程。a1~a4,b1~b5,c1~c4. 箭头所示为四分孢子囊细胞;a5, a6,c5. 箭头所示为四分孢子囊放散后留下的伤口,其中a6表示四分孢子已经释放出来a1-a6,b1-b6,c1-c6. The development of tetraspogonium. a1-a4,b1-b5,c1-c4. The arrows showed the tetraspogonium; a5, a6,c5. The arrows showed the wound left after the release of tetraspogonium, a6. The tetraspores had been released

图4 龙须菜四分孢子分裂后排列方式Fig.4 The arrangement pattern of the divided cells in tetraspogoniuma,b.“T”形排列四分孢子;c. “十”字形排列四分孢子a,b .“T” type; c. The cross vertical type

图5 藻体外四分孢子释放过程Fig.5 The releasing process of tetraspores outside the matrixa~e.显示四分孢子释放和发生变形变为球形的过程a-e.The process of releasing and deforming into spherical

图6 “981”龙须菜畸形的四分孢子囊及异常的四分孢子Fig.6 The deformed tetraspogonia of cultivar 981 a~f. 畸形的四分孢子囊;a. 表皮层中的畸形四分孢子囊,颜色粉色且畸形;b. 表皮层细胞横切面畸形四分孢子囊细胞;c,d. 四分孢子囊中的四分细胞不规则排布;e,f. 畸变四分孢子囊游离藻体外. g~l.四分孢子囊游离后溶解;g,h. 四分孢子囊开始溶解;i,j. 细胞溶解色素消失;k,l. 细胞中色素消失,溶解颗粒状碎片. m~n.四分孢子部分发生溶解,仅剩1个或2个正常细胞;o~p. 同一个四分孢子囊形成2个正常大小的四分孢子和2个较小的细胞a. The deformed tetraspogonia in the epidermal layer, the color is pink and the cell shape is deformed; b. The deformed tetraspogonium in the epidermal layer in the transaction; c,d. Irregular arrangement of the tetraspores in the deformed tetraspogonium.; e,f. The deformed tetraspogonium outside the matrix. g-l. The tetraspores dissolved after released. g,h. The tetraspogonium began to dissolve; i,j. The cells dissolved and the pigment decrease; k,l. The pigment of the cells disapeared and dissolved into granular debris. m-n. The tetraspores dissolved partially, only one or two normal tetraspores left; o-p. two normal tetraspores and two small tetraspores from one tetraspogonium
3.3 龙须菜不同部位细胞特征及细胞连接观察
3.3.1 龙须菜藻体不同部位细胞的组成和特征
龙须菜雌雄配子体和四分孢子体细胞组成,无明显差异,在基部接近固着器的地方,无髓部细胞(图7a),表皮层细胞5~20层,大小与最外表层细胞相近(图7a,b),髓部细胞中的红藻淀粉颗粒密度大,从基部到中部藻段,表皮层细胞层数减少到1~2层(图7c,d),髓部细胞出现并增多变大,细胞中的红藻淀粉颗粒密度减少(图7d)。再往上到尖端部位时,表层细胞只有一层,髓部细胞中含有较多色素,基本无红藻淀粉颗粒(图7e)。
3.3.2 细胞连接
配子体和四分孢子体均能在表皮层、皮层、髓部细胞中观察到细胞之间的连接,雌雄配子体和四分孢子体无明显差异。
细胞连接广泛存在龙须菜不同时期和不同部位的细胞之间。在表皮细胞之间、表皮细胞和皮层细胞之间,皮层细胞之间,皮层细胞与髓部细胞之间,髓部细胞之间普遍存在。细胞连接的数目,大小和形态在不同部位的连接存在差异。
(1)表皮细胞之间的细胞连接
最外一层的表皮细胞之间没有观察到细胞连接(图8 a1);下层的表皮细胞之间存在连接(图8 a2)。最外表皮细胞与下方紧邻的一层表层细胞之间存在连接(图8 b1~b3),由于大多数情况下,表皮细胞层数为1~2层,细胞连接存在于上下表皮细胞之间,及下层表皮细胞之间,两个细胞之间的连接数目通常为1个细胞连接。但是在野生四分孢子体基部,靠近附着器的部位,进行切片观察能观察到表皮细胞层数能达到3~4层或更多层,位于下层的多层表皮细胞之间存在上下表皮层细胞间纵向的细胞连接和同层表皮层细胞间横向的细胞连接(图8 c1~c3)。表皮层细胞排列紧密,在光镜下不经染色处理不易观察到表层细胞之间的连接。综上,表层细胞之间存在连接,连接数目通常为1个,细胞连接之间的方向排列较为规律,沿着横切半径方向或垂直于半径方向(纵向连接),连接处的直径大小约0.5 μm,长度约小于1 μm。染色结果显示,连接处PAS染色呈现红色,CBB染色呈现蓝色,说明含有多糖和蛋白组分。
(2)表皮细胞与皮层细胞的细胞连接
通常一个皮层细胞能与多个表皮细胞之间存在细胞连接,细胞连接长度约3~5 μm,直径大小约1~2 μm。通常2个细胞间连接长度不等长,表皮细胞的连接常短于皮层细胞的连接,2个表皮细胞连接到一个皮层细胞上,2个细胞连接呈”Y”型排列,表皮细胞与皮层细胞之间的细胞连接数目通常为1个(图8 d1~d3)。
(3)皮层细胞之间的细胞连接
皮层细胞之间的细胞连接数目通常也为一个,细胞连接处长度约为3~6 μm,直径大小约为1~2 μm,2个细胞之间的连接长度基本相同,2个细胞间的连接数目大多数为一个(图8 e1,e2)。
(4)皮层细胞与髓部细胞之间的细胞连接
皮层细胞与髓部细胞之间的细胞连接长度约6~10 μm,连接处直径大小约2~5 μm,细胞连接方向没有明显规律和方向性,皮层细胞与髓部细胞之间连接长度不等长,细胞连接的数目可能大于1个,1个皮层细胞可以与1个髓部细胞有1个或2个细胞连接,多个皮层细胞可以与1个髓部细胞间产生多个细胞连接(图8 f1~f3)。
(5)髓部细胞之间的连接
髓部细胞之间连接通常比较明显,细胞连接长度约为5~10 μm,连接处直径大小约为3~5 μm,2个髓部细胞间的连接长度基本等长(图8 g1~g5),2个相邻的髓部细胞之间可以形成数目3个以上,甚至超过10个以上的细胞连接(图8 g3)。髓部细胞在形成褶皱时,在同一个细胞褶皱处存在细胞连接(图8 g6)。
龙须菜不同层细胞连接的特征见表1。

表1 各层细胞间细胞连接特征比较

图7 龙须菜不同部位细胞组成及异常四分孢子囊Fig.7 Different parts of the cell composition in G. lemaneiformis and deformed tetraspogoniuma,b. 靠近固着器的基部;c. 基部藻段 表皮层细胞1~2层,淀粉颗粒密度大; d. 中部藻段 表皮层细胞1~2层,淀粉颗粒密度减少; e. 尖部藻段 表皮层细胞1层,无明显淀粉颗粒,含有色素; f. 不能正常释放四分孢子的游离四分孢子囊a,b. The base part near the holdfast; c. Base part, one or two layers of epidermal cells, with more starch granules; d. Middle part, one or two layers of epidermal cells, the density of the starch granules decreased; e. Tip part, only one layer of epidermal cells, little starch granules, had pigment granules; f. The deformed tetraspogonium did not release tetraspores

图8 龙须菜不同层间的细胞连接Fig.8 The pit connection in different layers of G. lemaneiformisa1.最外表皮细胞, a2.下层表皮细胞之间的细胞连接;b1~b3.最外表皮层和下层表皮细胞之间的细胞连接;c1~c3.基部靠近固着器部位多层表皮细胞之间的细胞连接;d1~d3.表皮层与皮层细胞间的细胞连接呈现“Y”形;e1,e2.皮层细胞之间的细胞连接;f1~f3.皮层和髓部细胞之间的细胞连接;g1~g6.髓部细胞之间的细胞连接。a1,a2,b2,c2,d2,e1,e2. PAS染色;b3,c3,d3. CBB染色;箭头示细胞连接a1. The most outside layer of epidermal cells, a2. The cell connections between the lower epidermal cells; b1-b3. Pit connection between the most outside epidermal cells and the lower epidermal cells; c1-c3. Pit connections between epidermal cells; d1-d3. The ”Y” pattern arrangement of pit connections between epidermal cells and the cortial cells; e1,e2. Pit connections between cortical cells; f1-f3. Pit connection between cortical cells and medullary cells; g1-g6. Pit connections between medullary cells, g1-g5. Pit connection between adjacent medullary cells; g6. Pit connection existed in the fold region of one medullary cell. a1,a2,b2,c2,d2,e1,e2. Dyeing by Perildate-Schiff reageant; b3,c3,d3. Dyeing by Coomassie Brilliant Blue; arrow showed cell connections
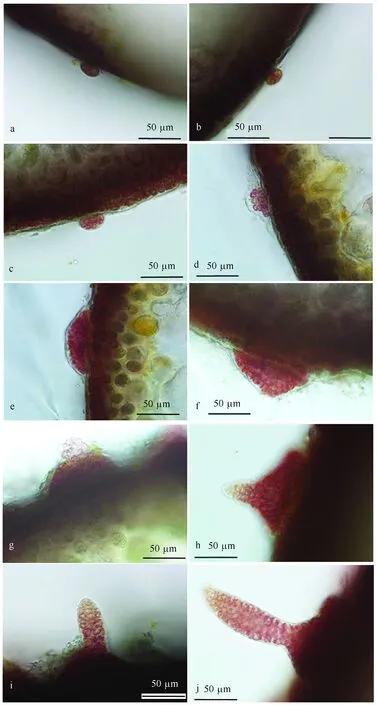
图9 附着在四分孢子体藻体表面的四分孢子生长发育过程Fig.9 Development process of the attached tetraspores on the tetrasporophytea. 四分孢子附着到四分孢子体表面细胞变扁平;b. 四分孢子分裂产生四细胞;c. 细胞继续分裂产生多层的细胞团;d. 细胞团向周边扩张,附着面积增大;e,f. 细胞不断分裂和扩张,形成半球形盘状体;g. 盘状体最顶端的细胞生长突出表面;h. 长出直立幼苗;i. 直立幼苗不断生长;j. 直立幼苗基部缢缩区域a. The tetraspore attached to the surface of the tetrasporophyte and became flat; b. Tetraspore divided into four cells; c. Cells grew to multilayer cell cluster; d. Cell cluster grew, and the attached area enlarged; e,f. Cell cluster grew and became hemispherical discoidal body; g. The cells in the tip of the discoidal body extruded; h. Upright seeding formed; i. The seeding grew up; j. The constricted region of the seedling

图10 配子体固着器包绕四分孢子体藻枝结构和藻体表面破损处配子体与四分孢子体细胞的分布及形态Fig.10 The structure of the holdfast grew surrounding the branch of the tetrasporophyte and the cell distribution between gametophyte and tetrasporophyte in epidermal damaged regiona,b.配子体固着器包绕覆盖四分孢子体藻枝一周,c,d. 配子体包绕覆盖四分孢子体藻枝横切;e. 配子体固着器与四分孢子体相交部位分界面;f.配子体固着器与四分孢子体相交部位分界面放大;g.配子体固着器与四分孢子体相交部位明显的分界线.(箭头示配子体与四分孢子体的分界线).h~j.位于藻体表面破损处配子体与四分孢子体细胞的分布及形态a,b. The holdfast wrapped the tetrasporophyte branch; c,d. The transection of the holdfast wrapped the branch; e. The boundary of holdfast of gametophyte and tetrasporophyte; f. The magnification of the boundary of holdfast of gametophyte and tetrasporophyte; g. The boundary of holdfast of gametophyte and tetrasporophyte. (arrow showed the boundary); h-j. The cell distribution between gametophyte and tetrasporophyte in epidermal damaged region
3.3.3 四分孢子释放后附着在藻体上生长发育过程
实验室培养条件下,观察到龙须菜释放的四分孢子与四分孢子体在同一培养体系下,部分四分孢子会附着在四分孢子体的藻枝上,生长分裂形成配子体幼苗,在生长发育的过程中,部分配子体幼苗会从四分孢子体上脱落;部分配子体幼苗会一直附着在四分孢子体表面,固着器固着面积不断扩大,覆盖藻枝,四分孢子体和配子体混生在一起。而四分孢子附着在四分孢子体生长发育的过程如下:
(1)部分游离的四分孢子附着在四分孢子体藻体表面,四分孢子细胞变形,细胞由球形变为扁球形附于藻体表面(图9 a)。
(2)细胞开始分裂,最开始的几次分裂仅增加细胞数目,整体体积无变化,细胞体积不断减小,使得四分孢子由一个细胞形成一个多层细胞团结构(图9b,c)。
(3)细胞生长,细胞体积变大,细胞不断进行分裂,附着的边缘沿着藻体扩大,形成半球形的盘状体(图9 d,e)。
(4)半球形结构顶端的几个细胞快速分裂和生长,形成突出半球形藻体表面的细小突起(图9 f,g)。突起处的细胞不断分裂生长,伸长产生幼小的主枝,主枝靠近盘状体的下部藻段有一段缢缩区域(图9 h,i,j)。
(5)主枝细胞继续分裂生长,主枝伸长变粗并产生分枝,主枝生长过程中固着器沿着四分孢子体藻体表面不断生长,固着面积不断扩大,覆盖四分孢子体藻体表面,甚至能够包裹四分孢子体藻枝,扩大生长的固着器上能形成新的突起,继续生长为新的藻枝,形成四分孢子体和配子体混生在一起的现象。
四分孢子附着在四分孢子体上生长发育为配子体过程中,有一部分配子体的固着器与藻体表面接触较小,随着主枝生长,没有明显生长,这种配子体容易从附着藻体表面脱落,在生长发育过程中很大一部分附着生长的配子体幼苗发生脱落。四分孢子附着在完整无破损的四分孢子体表面时,四分孢子发育产生的配子体与四分孢子体在横切面上观察分界明显,配子体的固着器随着主枝的生长也不断分裂生长,附着面积不断扩大,覆盖甚至包绕四分孢子体藻体,使得配子体较为牢固的附着在四分孢子体上,外观形态上配子体与四分孢子体生长在同一藻枝上(图10 a,b),显微镜下切片观察,配子体的固着器包绕四分孢子体藻段一周(图10 c,d),配子体和四分孢子体之间能区分出细胞界线(图10 e~g);四分孢子附着在有破损的四分孢子体上时,配子体的细胞分裂生长,填充四分孢子体的破损伤口处,使得四分孢子体和配子体细胞嵌合生长在一起(图10 h~j),也导致配子体和四分孢子体不同世代混杂。
4 讨论
4.1 配子体生殖结构生长发育过程
雌雄配子体在性成熟前经过幼苗期后,进入毛细胞期,表皮层中的出现毛细胞并伴随毛细胞的生长,毛结构的伸长,藻体快速生长。徒手切片连续观察没有发现雌雄配子体性成熟之前的差异,将藻体雌雄区分开来。雌雄性别的确定在杂交实验中至关重要,所以新的技术手段仍需要开发。Zhou等[17]和Li等[20]的SCAR标记,借助分子手段鉴定雌雄配子体,对于早期配子体幼苗的筛选具有重要指导意义。
本研究首次在龙须菜中报道了毛细胞的存在。毛细胞在红藻中普遍存在,超过4 000种红藻有产毛细胞[21],Sjostedt[22]对Gracilariacompressa毛细胞发育和结构进行了描述,毛细胞起源表皮细胞,起初大小和形状与表皮细胞无差别,随后细胞变大,顶端产生小突起并伸长产生毛结构。Yamamoto[23]对G.Punctata(Okam) Yamada和G.arcuateZanard的描述的毛细胞成簇分布于藻体表面,从对龙须菜毛细胞观察结果来看,龙须菜的毛细胞和毛结构的特征与上述描述物种中的毛细胞结构特征一致,但龙须菜中毛细胞分布并不广泛。有关毛细胞产生的条件在其他物种中均有报道与季节[24]、光周期和N元素[25—26],本实验也观察到龙须菜的毛细胞并不总是存在,每次出现时藻体状态良好,可能在最适合其生长的条件下才会出现,毛细胞的出现可能是为了短暂满足藻体的营养和能量需求。但是关于毛细胞产生毛的作用目前尚无定论。
4.2 不同品系龙须菜四分孢子体四分孢子形成过程的生长发育过程
四分孢子的形成和释放过程伴随着藻体细胞形态的一系列变化,四分孢子母细胞来源于表皮层细胞,颜色鲜红色,细胞生长体积增大,随后发生2次减数分裂,在四分孢子囊内形成4个四分孢子,成熟后的四分孢子囊从藻体上脱离,四分孢子在藻体外释放变形形成球形四分孢子。在四分孢子囊的生长发育过程中,髓部细胞中的红藻淀粉颗粒呈现减少趋势,说明四分孢子囊的生长发育消耗了部分红藻淀粉,红藻淀粉是在红藻中是一种能源储备物质。Wang等[15]对龙须菜四分孢子形成释放阶段的ATP含量测定结果显示,藻体从四分孢子形成到释放过程,ATP含量呈现下降趋势,也说明四分孢子的放散过程是一个能量大量消耗的过程,与形态学上观察到的红藻淀粉颗粒减少呈现一定相关性。当藻受到镉胁迫[27]或营养盐N/P胁迫[28]影响时红藻淀粉颗粒增多,也说明红藻淀粉作为能源储备物质应对外界环境胁迫,而四分孢子放散后红藻淀粉颗粒明显减少,同时表皮细胞中存在破损伤口,可以推测后续随着藻体的伤口愈合髓部细胞中红藻淀粉会重新积累。四分孢子在四分孢子囊中的分布呈十字形的很少,在一个视野平面上多数能看到3个细胞,说明第二次分裂的2个细胞分裂面并不都是重合的,2个分裂面存在一定角度。四分孢子的释放在藻体外进行,四分孢子从四分孢子囊中逐个释放出来并变形呈球形,释放的过程能看到细胞质的流动,在四分孢子囊中形成的四分孢子原始细胞并没有细胞壁,在释放变成球形四分孢子后才开始形成新的细胞壁。而“981”龙须菜形成的四分孢子囊畸形个数较多,能观察到较多变形、颜色异常、分裂异常的四分孢子囊,这些与野生型龙须菜和“鲁龙1号”良种存在差异,高畸形率可能是除自身形成四分孢子数量少外,导致“981”龙须菜低育的一个原因。
4.3 龙须菜不同部位细胞特征及特殊细胞结构观察
龙须菜藻体不同部位的细胞组成和细胞特征存在差异,固着器及接近固着器的基部表皮层细胞层数较多,髓部细胞从无到有,靠近基部的藻体内淀粉颗粒密度大,沿形态学上端方向,表皮层细胞层数减少,髓部细胞数量增多,体积增大,含有的淀粉颗粒密度减少;到藻尖部位,表皮层1层,髓部细胞体积数目又变少,红藻淀粉含量很少,含有色素颗粒。
细胞连接(pit connection)在红藻多个种属[29—32]中均有发现,在超微结构上不同种属上存在差异,超微结构下细胞连接处都含有蛋白质栓塞(pit plug)[33]。在光镜下,通过染色等手段,观察到龙须菜不同部位和不同层次细胞之间的细胞连接的特征,在连接数量和大小方面存在差异,总体上从表皮层到髓部细胞,细胞连接由小到大,数量由少到多,与细胞大小变化趋势一致。细胞连接根据连接来源可以分为初级和次级纹孔连接[34],最外表皮层细胞与下层表皮层细胞之间的细胞连接为初级纹孔连接,由于细胞有丝分裂不完全,保留在2个子代细胞间;皮层和髓部细胞间的细胞连接众多,而PAS染色和CBB染色结果显示,连接处含有多糖和蛋白质组分。
放散出的四分孢子能附着在四分孢子体上,进行生长产生配子体幼苗,出现世代混杂的现象。配子体附着在四分孢子体上,固着器会不断生长扩大附着面积甚至包绕四分孢子体母体藻段而牢固结合或通过四分孢子体上的伤口侵入四分孢子体内与四分孢子体的细胞嵌合在一起。相关文献中有报道江蓠科藻类在孢子早期发育中会发生相邻盘状体或孢子早期发育中的黏连嵌合现象,真江蓠果孢子盘状体在发育过程中黏连,真江蓠中果孢子和四分孢子在早期发育中会发生盘状体黏连[35],江蓠中也存在盘状体黏连嵌合现象[36—37],真红藻亚纲中很多物种在孢子早期生长发育过程中产生嵌合现象[38—39],江蓠属Gracilariatikvahiae,G.deblilis,G.gracilis中也发现四分孢子体和配子体的嵌合混生现象[40]。说明世代混杂在江蓠科中普遍存在,增加了遗传的多样性和复杂性。
总的来说,龙须菜配子体和孢子体在生长发育过程中在形态和细胞结构上存在相似性,而在产生生殖结构的过程中表现出明显差异,雌雄配子体前期的生长没有明显差异。观察到了四分孢子的形成放散的完整过程,不同品系龙须菜形成的四分孢子存在差异,其中“981”龙须菜有较为明显的畸形现象,在细胞形态层面上说明了“981”龙须菜低育的原因。另外对龙须菜不同部位不同层次的细胞特征及细胞连接进行观察,对龙须菜生长发育研究有了更加深入的认识。
[1] Lapointe B E, Ryther J H. Some aspects of the growth and yield ofGracilariatikvahiaein culture[J]. Aquaculture, 1978, 15(3): 185-193.
[2] McHugh D J. Worldwide distribution of commercial resources of seaweeds includingGelidium[J]. Hydrobiologia, 1991, 221(1): 19-29.
[3] Chopin T, Buschmann A H, Halling C, et al. Integrating seaweeds into marine aquaculture systems: a key toward sustainability[J]. Journal of Phycology, 2001, 37(6): 975-986.
[4] Lu Huiming, Xie Haihui, Gong Yingxue, et al. Secondary metabolites from the seaweedGracilarialemaneiformisand their allelopathic effects onSkeletonemacostatum[J]. Biochemical Systematics and Ecology, 2011, 39(4/6): 397-400.
[5] 余杰, 王欣, 陈美珍, 等. 潮汕沿海龙须菜的营养成分和多糖组成分析[J]. 食品科学, 2006, 27(1): 93-97.
Yu Jie, Wang Xin, Chen Meizhen, et al. Analysis on nutritional components and polysaccharide composition ofGracilarialemaneiformisfrom Chaoshan coast[J]. Food Science, 2006, 27(1): 93-97.
[6] Lu Huiming, Liao Xiaojian, Yang Yufeng, et al. Effects of extracts fromGracilarialemaneiformon microalgae[J]. Ecological Science, 2008, 27(5): 424-426.
[7] 徐永健, 钱鲁闽, 焦念志. 江蓠作为富营养化指示生物及修复生物的氮营养特性[J]. 中国水产科学, 2004, 11(3): 276-280.
Xu Yongjian, Qian Lumin, Jiao Nianzhi. Nitrogen nutritional character ofGracilariaas bioindicators and restoral plants of eutrophication[J]. Journal of Fishery Sciences of China, 2004, 11(3): 276-280.
[8] 张学成, 费修绠, 王广策, 等. 江蓠属海藻龙须菜的基础研究与大规模栽培[J]. 中国海洋大学学报:自然科学版, 2009, 39(5): 947-954.
Zhang Xuecheng, Fei Xiugeng, Wang Guangce, et al. Genetic studies and large scale cultivation ofGracilarialemaneiformis[J]. Periodical of Ocean University of China, 2009, 39(5): 947-954.
[9] 孟琳, 徐涤, 陈伟洲, 等. 龙须菜新品系07-2的筛选及性状分析[J]. 中国海洋大学学报, 2009, 39(S1): 94-98.
Meng Lin, Xu Di, Chen Weizhou, et al. Selection and characterization of a new strain ofGracilarialemaneiformis[J]. Periodical of Ocean University of China, 2009, 39(S1): 94-98.
[10] 周伟, 隋正红. 龙须菜“鲁龙1号”[J]. 中国水产, 2016(3): 56-57.
Zhou Wei, Sui Zhenhong. The Cultivar “Lulong NO.1” ofGracilariopsislemaneiformis(Gracilariaceae, Rhodophyta)[J]. China Fisheries, 2016(3): 56-57.
[11] 张学成, 秦松, 马家海, 等. 海藻遗传学[M]. 北京: 中国农业出版社, 2005.
Zhang Xuecheng, Qin Song, Ma Jiahai, et al. The Genetic of Marine Algae[M]. Beijing: China Agriculture Press, 2005.
[12] Dawson E Y. Studies of northeast pacific gracilariaceae[J]. Allan Hancock Foundation Publications, Occasional Papers, 1949, 7: 1-105.
[13] Fredericq S, Hommersand M H. Comparative morphology and taxonomic status ofGracilariopsis(Gracilariales, Rhodophyta)[J]. Journal of Phycology, 1989, 25(2): 228-241.
[14] Xu Di, Zhu Jianyi, He Yang, et al. Structure of reproductive apparatus ofGracilaria/Gracilariopsislemaneiformis(Gracilariaceae, Rhodophyta)[J]. Science in China Series C: Life Sciences, 2008, 51(12): 1137-1143.
[15] Wang Jinguo, Sui Zhenghong, Hu Yiyi, et al. Assessment of photosynthetic performance, carboxylase activities, and ATP content during tetrasporic development inGracilariopsislemaneiformis(Gracilariaceae, Rhodophyta)[J]. Journal of Applied Phycology, 2016, 28(5): 2939-2952.
[16] Ye Naihai, Wang Hongxia, Wang Guangce. Formation and early development of tetraspores ofGracilarialemaneiformis(Gracilaria, Gracilariaceae) under laboratory conditions[J]. Aquaculture, 2006, 254(1/4): 219-226.
[17] Zhou Wei, Ding Hongye, Sui Zhenghong, et al. Development of a SCAR marker for male gametophyte ofGracilariopsislemaneiformisbased on AFLP technique[J]. Chinese Journal of Oceanology and Limnology, 2014, 32(3): 522-526.
[18] Rover T, Simioni C, Hable W, et al. Ultrastructural and structural characterization of zygotes and embryos during development inSargassumcymosum(Phaeophyceae, Fucales)[J]. Protoplasma, 2015, 252(2): 505-518.
[19] Dring D M. A periodic Acid-Schiff technique for staining fungi in higher plants[J]. New Phytologist, 1955, 54(2): 277-279.
[20] Huan Li, He Linwen, Zhang Baoyu, et al. AFLP and SCAR markers associated with the sex inGracilarialemaneiformis(Rhodophyta)[J]. Journal of Phycology, 2013, 49(4): 728-732.
[21] Kraft G T, Woelkerling W J. Rhodophyta[M]//Clayton M N, King R J. Biology of Marine Plants. Melbourne: Longman Cheshire, 1990: 41-85.
[22] Sjöstedt L G. Floridean Studies[M]. Otto Harrassowitz: CWK Gleerup, 1926.
[23] Yamamoto H. Systematic and anatomical study of the genusGracilariain Japan[J]. Memoirs of the Faculty of Fisheries, Hokkaido University, 1978, 25(2): 97-152.
[24] Dixon P S. Biology of the Rhodophyta[M]. Edinburgh: Olivier and Bold, 1973: 285.
[25] Rueness J, Mathisen H A, Tananger T. Culture and field observations onGracilariaverrucosa(Huds.) Papenf. (Rhodophyta) from Norway[J]. Botanica Marina, 1987, 30(3): 267-276.
[26] O′connor K A, West J A. The effect of light and nutrient conditions on hair formation inSpyridiafilamentosa(Wulfen) Harvey (Rhodophyta)[J]. Botanica Marina, 1991, 34(4): 359-364.
[27] dos Santos R W, Schmidt É C, Bouzon Z L. Changes in ultrastructure and cytochemistry of the agarophyteGracilariadomingensis(Rhodophyta, Gracilariales) treated with cadmium[J]. Protoplasma, 2013, 250(1): 297-305.
[28] Yu Jiang, Yang Yufeng. Physiological and biochemical response of seaweedGracilarialemaneiformisto concentration changes of N and P[J]. Journal of Experimental Marine Biology and Ecology, 2008, 367(2): 142-148.
[29] Lee R E. The pit connections of some lower red algae: ultrastructure and phylogenetic significance[J]. British Phycological Journal, 1971, 6(1): 29-38.
[30] Ramus J. Pit connection formation in the red algaPseudogloiophloea[J]. Journal of Phycology, 1969, 5(1): 57-63.
[31] Wetherbee R, Quirk H M. The fine structure of secondary pit connection formation between the red algal alloparasiteHolmsellaaustralisand its red algal hostGracilariafurcellata[J]. Protoplasma, 1982, 110(3): 166-176.
[32] Bourne V L, Conway E, Cole K. On the ultrastructure of pit connections in the conchocelis phase of the red algaPorphyraperforataJ. Ag[J]. Phycologia, 1970, 9(1): 79-81.
[33] 王素娟, 裴鲁青, 段德麟. 中国常见红藻超微结构[M]. 宁波: 宁波出版社, 2004: 138-158.
Wang Sujuan, Pei Luqing, Duan Delin. Ultrastructure of Common Red Seaweeds in China[M]. Ningbo: Ningbo Press, 2004: 138-158.
[34] 许璞, 张学成, 王素娟, 等. 中国主要经济海藻的繁殖与发育[M]. 北京: 中国农业出版社, 2013: 85-90.
Xu Pu, Zhang Xucheng, Wang Sujuan, et al. Reproduction and Development of Major Economic Marine Algae in China[M]. Beijing: China Agriculture Press, 2013: 85-90.
[35] Jones W E. Effect of spore coalescence on the early development ofGracilariaverrucosa(Hudson) papenfuss[J]. Nature, 1956, 178(4530): 426-427.
[37] Santelices B, Correa J A, Aedo D, et al. Convergent biological processes in coalescing Rhodophyta[J]. Journal of Phycology, 1999, 35(6): 1127-1149.
[38] Santelices B, Aedo D, Varela D. Causes and implications of intra-clonal variation inGracilariachilensis(Rhodophyta)[J]. Journal of Applied Phycology, 1995, 7(3): 283-290.
[39] Santelices B, Correa J A, Meneses I, et al. Sporeling coalescence and intraclonal variation inGracilariachilensis(Gracilariales, Rhodophyta)[J]. Journal of Phycology, 1996, 32(2): 313-322.
[40] Kain J M, Destombe C. A review of the life history, reproduction and phenology ofGracilaria[J]. Journal of Applied Phycology, 1995, 7(3): 269-281.
Microscopic observation of the reproductive development of gametophyte and tetrasporohyte ofGracilariopsislemaneiformis(Rhodophyta)
Mi Ping1, Sui Zhenghong1, Que Zhou1, Zhou Wei1, Shang Erlei1
(1.TheKeyLaboratoryofMarineGeneticsandBreeding,OceanUniversityofChina,MinistryofEducation,Qingdao266003,China)
A series of cell morphology and structure changes were displayed during the development of reproductive structure of gametophyte and tetrasporaphyte ofGracilariopsislemaneiformis. In this study, the gametophyte and different strains of tetrasporophyte were used to make continuous free hand section, in order to observe the development of the gametophyte and the formation of tetraspores in different strains ofG.lemaneiformis. The result showed that the development of male and female gametophytes undertook seedling phage (SP), hair cell phage (HCP) and sexual mature phage (SMP). There was no significant difference between male and female gametophyte in SP and HCP. The male and female gametophyte were differentiable after entering SMP when the reproductive apparatus formed. The tetraspores were derived from the epidermal cells of tetrasprophyte, the initial tetrasporogonium was scarlet, and 5-10 μm in diameter. It later grew to 20-25 μm in diameter and became mature. The mature tetrasporogonium divided into four initial tetraspores gathered together, and then the tetraspores were released out of the matrix and became globular. During the development of tetraspores, the density of floridean starch granules decreased in medullary part. Cultivar 981 formed more deformed tetraspores, which may be one of the reasons leading to the low fertility of cultivar 981. The layers of epidermal cells, the numbers and the volume of medullary cells changed continuously from the basal to the tip part. Pit connections existed wildly in the epidermal, cortical and medullary cells, however varying by size, numbers and length between different layers. The released tetraspores were observed attaching on the tetrasporophyte, and the holdfast grew to cover the tetrasporophyte or invaded into the wound tissue of tetrasporophyte, which may cause gametophyte and tetrasporophyte generation promiscuous and lead to the genetic complexity ofG.lemaneiformis.
Gracilariopsislemaneiformis; gametophyte; tetrasporophyte; reproductive apparatus; microscopic observation
10.3969/j.issn.0253-4193.2017.08.010Mi Ping, Sui Zhenghong, Que Zhou,et al. Microscopic observation of the reproductive development of gametophyte and tetrasporohyte ofGracilariopsislemaneiformis(Rhodophyta) [J]. Haiyang Xuebao,2017,39(8):99—114, doi:10.3969/j.issn.0253-4193.2017.08.010
2016-10-28;
2017-02-24。
国家自然科学基金(31372529);鳌山科技创新计划课题(2015ASKJ02);中国博士后科学基金项目(2016M590662)。
米萍(1992—),女,山东省临沂市人,主要研究藻类遗传育种与发育。E-mail:miping10@163.com
*通信作者:隋正红,教授,主要从事大型红藻的遗传育种与基因组研究。E-mail:suizhengh@ouc.edu.cn
S917.3
A
0253-4193(2017)08-0099-16
米萍,隋正红,阙州,等. 龙须菜配子体发育和不同品系四分孢子体四分孢子形成过程显微结构观察[J].海洋学报,2017,39(8):99—114,
猜你喜欢
上海农业科技(2022年1期)2022-11-18
中国海洋大学学报(自然科学版)(2022年8期)2022-08-15
广西科学(2022年1期)2022-04-12
福建农业学报(2021年10期)2022-01-07
中国饲料(2021年17期)2021-11-02
热带海洋学报(2021年1期)2021-03-05
现代农业科技(2020年21期)2020-12-14
水产科学(2019年4期)2019-07-23
生物学通报(2019年7期)2019-07-08
科技创新与应用(2017年20期)2017-07-15
