
新疆水稻主要育成品种13个稻瘟病的抗性基因分布
2017-10-16 02:13张燕红贾春平文孝荣唐福森王奉斌朱小霞
新疆农业科学 2017年9期
张燕红,贾春平,文孝荣,唐福森,王奉斌,朱小霞,
(1.新疆农业科学院核技术生物技术研究所,乌鲁木齐 830091;2.新疆农业科学院温宿水稻试验站,新疆温宿 843008)
新疆水稻主要育成品种13个稻瘟病的抗性基因分布
张燕红1,贾春平1,文孝荣2,唐福森2,王奉斌1,朱小霞1,2
(1.新疆农业科学院核技术生物技术研究所,乌鲁木齐 830091;2.新疆农业科学院温宿水稻试验站,新疆温宿 843008)
目的明确新疆主要育成水稻品种中稻瘟病抗性基因的分布状况,为稻瘟病抗性育种提供依据。方法研究以31份新疆选育的水稻品种为材料,利用13个抗稻瘟病基因的特异性分子标记,进行PCR鉴定,初步建立新疆主要育成品种的抗性基因分布情况。结果31份供试材料中均无Pi21基因存在;表现为广谱抗性的Pi9、Pita、Pib和Pikm基因所占比例偏少,分别占9.68%、22.58%、6.45%、12.90%;表现为广谱抗性的Pi2、Pigm、Pil、Pikh/Pi54、Pi63和表现为生理小种特异性抗性的Pia、Pid2、Pid3/Pi25基因广泛存在,所占比率67.74%~100%;不同水稻品种抗性基因数量分布在5~11个,含11个抗性基因的品种是新稻23号,其聚合了除Pi21和Pid2外其余所有抗性基因。结论新疆主要水稻育成品种中广泛存在具有光谱抗性的Pi2、Pigm、Pil、Pikh/Pi54、Pi63和表现为生理小种特异性抗性的Pia、Pid2、Pid3/Pi25基因,利用特异性分子标记可以有效快速鉴定品种中的抗性基因的存在和分布情况。
水稻;育成品种;稻瘟病;抗性基因
0 引 言
【研究意义】稻瘟病由真菌(PyriculariagriseaSacc.)引起,既造成水稻(OryzasativaL.)减产,又导致米质下降,是水稻主产区面临的极为棘手的病害之一[1]。目前,种植抗病品种是最为经济、有效的防治策略,但主栽品种多年大面积田间种植后,因抗病基因单一、遗传背景狭窄和稻瘟病菌变异造成的抗性“衰退”直至“丧失”,病害再次爆发流行的现象仍屡见不鲜[2-5]。发掘抗稻瘟病基因,拓宽抗病遗传基础并应用于抗病育种研究工作中可谓当务之急。因此,充分利用现有资源品种中的抗病材料,鉴定不同抗病基因型,不同抗病基因型品种轮流使用和合理布局的有效施行,对通过基因聚合手段的抗病育种工作的大力推进和持续有效的控制稻瘟病的流行发生起至关重要的作用。【前人研究进展】近年来,水稻等作物全基因组测序的陆续公布,分子标记技术及其应用迅猛革新,大幅度提升了鉴定效率。与此同时,有关稻瘟病基因的定位与克隆也获得了极大进展。除水稻3号染色体外,现已有超过90个稻瘟病抗性基因被定位在其它11条染色体上[6]。其中3个主要的基因簇存在于6号、11号和12号染色体上,分别称之为pi9、pik及pita位点。其中,Pi9、Pita、Pib、Pikm、Pi2、Pigm、Pil、Pikh/Pi54具有广谱抗性[5]。Pia、Pid2、Pid3/Pi25表现为生理小种特异性抗性,对应的无毒菌株(小种)依次为B90002,ZB15和Zhong-10-8-14(国家水稻数据中心. 稻瘟病主效抗性基因列表[DB/OL]. http://www.ricedata.cn/gene/gene_pi.htm, 2012-6-20.)。Pi9基因来源于小粒野生稻,编码的蛋白产物在氨基端含有NBS域,在羧基端含有富亮氨酸重复域(LRRs),对来自13个国家的43个稻瘟病菌株表现高抗,组成性表达并不受稻瘟病侵染诱导,不留过敏病斑[7]。Pita基因起源于籼稻,由籼稻逐渐渗入到粳稻中,编码细胞质膜受体蛋白,其位点上抗、感基因编码的产物仅含一个氨基酸的差别。Pikm基因和Pikh基因分别来自高抗品种Tsuyuake和Tetep。Pikm基因基本不受稻瘟病菌接种影响,为组成型表达;而Pikh基因的表达受稻瘟病菌诱导。二者分离于11号染色体长臂的Pik位点,编码的蛋白同属于NBS-LRR家族[8-9]。于2009年被成功克隆的Pigm基因来自广谱、持久抗稻瘟病品种谷梅4号。等位性检测及接种结果表明,Pigm基因与Pi9基因及Pi2基因紧密连锁,相较于Pi9、Pi2和Piz基因,其抗谱更宽,育种应用潜力更大[6]。Pil基因是源自利比亚粳型品种“LAC23”的广谱稻瘟病抗性基因,定位在第 11 染色体上MGR4766标记1.3 cm处,有研究显示其与Pi2基因有良好的互补效应[10-11]。Pid2基因编码受体蛋白激酶(B lectin)。Pi21基因来自旱稻品种‘Owarihatamochi’,编码富含脯氨酸的蛋白结构(Proline-rich protein),是激发慢速抗病反应与基础抗性相关的非小种特异性基因。Pib和Pita是最早被克隆出来的2个抗稻瘟病基因[12-13]。Pib与Pita基因有43.2%的相似性,来自日本抗性水稻材料BL1,属小家族成员之一,对日本绝大多数小种表现高抗性,位于第2染色体长臂近末端区域,编码多肽,因温度、光照等环境条件的变化而诱导调控并影响Pib基因的表达。最近,源于这2个基因及其等位感病位点的一些功能标记也已被开发出来,可以快速、准确地从水稻种质资源中鉴定出Pita和Pib基因[14]。鉴于一些Pita和Pib基因源品种,诸如IR系列(Pib/Pita)、Tetep(Pita)、特青(Pita)、日本BL系列(Pib)品种等,曾广泛应用于我国水稻抗病育种,且Pita和Pib基因目前在我国很多稻区仍表现广谱稻瘟病菌抗性[15-16]。王忠华等[17]成功地将Pita基因分子标记运用到水稻抗病育种中,并设计了田间接种试验,验证了此标记的准确性。刘洋等[18]利用此分子标记对抗病基因Pib进行检测,也设计了田间接种试验,验证了此标记的准确性。张羽等[19]利用该方法研究了pi9基因在陕西省水稻品种的分布,李进斌[20]等研究结果表明Pita和Pib广泛存在云南水稻资源中,其鉴定结果与接菌验证结果相符。时克等[15]研究利用源于Pita和Pib基因本身的特异性分子标记,结合稻瘟病菌接种鉴定,检测和分析了我国58份水稻主栽品种(杂交稻亲本)的Pita和Pib抗性基因型,结果表明,特籼占25、佳禾早占、密阳46、测64-7等4个籼稻品种携带Pita和Pib基因;籼小占等4个籼稻品种(系)和早丰9号等5个粳稻品种携带Pita基因;绵恢501等5个籼稻品种(系)和粳稻品种武育粳7号、辽粳454携带Pib基因。【本研究切入点】以上这些基因或等位感病位点的一些功能标记的开发,可快速、准确、系统地鉴定水稻品种抗稻瘟病基因型,为高效开展抗稻瘟病基因聚合育种带来了新的契机。新疆具有幅员辽阔、地形复杂、气候变化大等特点,稻作资源不同及稻区表现不同,造就了新疆水稻品种资源丰富的遗传多样性。研究利用13个稻瘟病基因的14对分子标记,对新疆自育主要品种资源31份材料进行PCR检测,鉴定各个品种的所含抗稻瘟病基因型。【拟解决的关键问题】研究这些基因型在新疆选育品种中的分布特征,为抗稻瘟病基因在新疆稻种资源中的分布、稻瘟病预防及防治提供依据。
1 材料与方法
1.1 材 料
选取新疆维吾尔自治区种子站审认定的水稻品种以及地方审定的品种共计31份,并具有较大的推广面积的作为试验的参试材料。品种来自于新疆农业科学院核技术生物技术研究所,品种均为实验室长期搜集保存。表1
表1 供试水稻材料及来源
Table 1 Material for experiment

编号Code品种Variety来源Source编号Code品种Variety来源Source1新稻1号新疆农科院核生所17新稻36号新疆农科院核生所2新稻10号新疆兵团第一师18新稻39号新疆农科院核生所3新稻11号新疆农科院核生所19新稻41号新疆农科院粮作所4新稻12号新疆农科院核生所20新稻42号新疆农科院核生所5新稻14号新疆兵团第一师21新稻44号新疆农科院核生所6新稻17号新疆农科院粮作所22新稻45号新疆农科院核生所7新稻19号新疆金丰源种业23A稻4号新疆兵团第一师8新稻21号新疆塔河种业24A稻7号新疆兵团第一师9新稻23号新疆兵团第一师25A稻8号新疆兵团第一师10新稻24号新疆农科院核生所26伊粳8号新疆伊犁州农科所11新稻27号新疆伊犁州农科所27伊粳12号新疆伊犁州农科所12新稻29号新疆农科院核生所28伊粳13号新疆伊犁州农科所13新稻30号新疆农科院核生所29阿稻1号新疆兵团第一师14新稻33号新疆金丰源种业30粮粳5号新疆农科院粮作所15新稻34号新疆金丰源种业31粮香5号新疆农科院粮作所16新稻35号新疆农科院核生所
1.2 方 法
1.2.1 DNA提取和检测
采集幼嫩叶片利用传统CTAB法提取不同水稻材料的DNA。利用超微量分光光度计检测DNA浓度、质量,后琼脂糖凝胶电泳检测DNA完整性。样品置于冰箱-20℃中备用。
1.2.2 13个抗性基因信息
研究选取的13个抗性基因中Pi9、Pita、Pib、Pikm、Pi2、Pigm、Pil、Pikh/Pi54具有广谱抗性[5]。Pia、Pid2、Pid3/Pi25表现为生理小种特异性抗性,对应的无毒菌株(小种)依次为B90002,ZB15和Zhong-10-8-14(国家水稻数据中心. 稻瘟病主效抗性基因列表[DB/OL].http://www.ricedata.cn/gene/gene_pi.htm,2012-6-20.)。表2
1.2.3 引物设计与PCR扩增
Pita基因分子标记采用Jia等[21-22]报道的引物YI 155/YL87和YL183/YI87。YL155/YI87特异性扩增抗病等位基因Pita的相应序列,约1 kb的DNA片段;L183/YL87特异扩增感病等位基因Pita的相应序列,约1 kb的DNA片段。根据Fjellstrom等[23]报道的序列合成引物PibdomF/PibdomR,能特异性扩增出抗病等位基因Pib的相应序列,约365 bp的DNA片段;根据刘洋等[18]报道的序列合成引物Lys145F/Lys145R,扩增Pib感病等位基因的相应片段,约803 bp的DNA片段,不能扩增出抗病等位基因Pib的相应片段。这些引物均由上海祥音生物科技有限公司合成。列出引物序列及引物名称。表3
PCR反应体系为25 μL,含50 ng/μL模板DNA 1 μL,2×dNTP 2 μL,10×Buffer 2.5 μL,10 μmol/L的上下游引物各0.5 μL,10 U/μLTaq酶0.5 μL,超纯水18 μL。反应程序:94℃预变性4 min,94℃变性30 s,55℃退火30 s,72 ℃延伸30~60 s,30个循环,最后72 ℃延伸7 min。
扩增产物在2%琼脂糖凝胶电泳下分离,核酸染料(Gelviem)染色,使用Quantity One凝胶成像软件紫外灯下观察拍照。在相同条件下,每个DNA样品重复扩增并电泳检测2次以上,以确保扩增结果准确可靠。
表2 13个抗性基因信息
Table 2 Information for 13resistance genes of rice blast
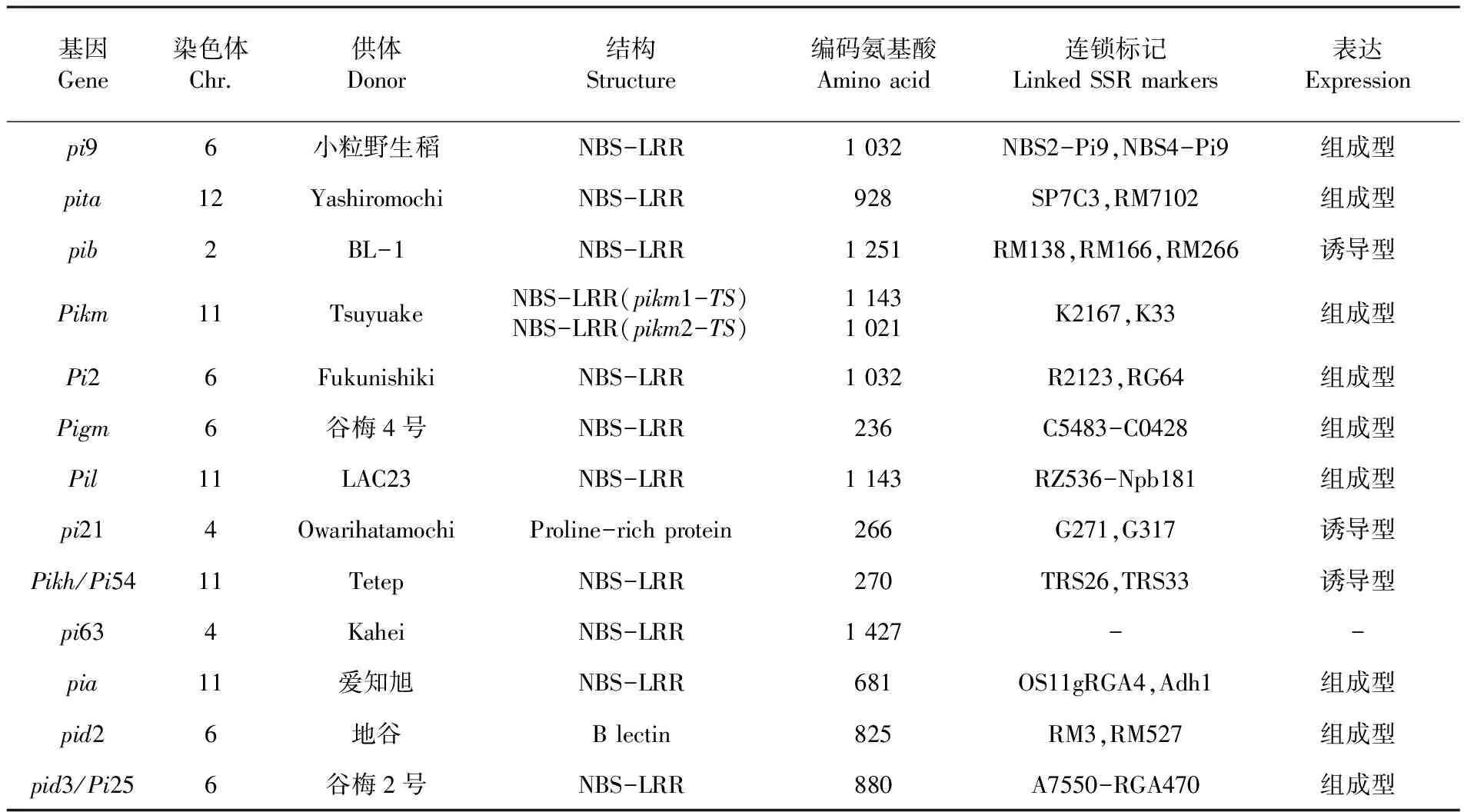
基因Gene染色体Chr.供体Donor结构Structure编码氨基酸Aminoacid连锁标记LinkedSSRmarkers表达Expressionpi96小粒野生稻NBS-LRR1032NBS2-Pi9,NBS4-Pi9组成型pita12YashiromochiNBS-LRR928SP7C3,RM7102组成型pib2BL-1NBS-LRR1251RM138,RM166,RM266诱导型Pikm11TsuyuakeNBS-LRR(pikm1-TS)NBS-LRR(pikm2-TS)11431021K2167,K33组成型Pi26FukunishikiNBS-LRR1032R2123,RG64组成型Pigm6谷梅4号NBS-LRR236C5483-C0428组成型Pil11LAC23NBS-LRR1143RZ536-Npb181组成型pi214OwarihatamochiProline-richprotein266G271,G317诱导型Pikh/Pi5411TetepNBS-LRR270TRS26,TRS33诱导型pi634KaheiNBS-LRR1427--pia11爱知旭NBS-LRR681OS11gRGA4,Adh1组成型pid26地谷Blectin825RM3,RM527组成型pid3/Pi256谷梅2号NBS-LRR880A7550-RGA470组成型
2 结果与分析
2.1 13个抗性基因的分子鉴定
利用13个抗性基因对应的特异性分子标记分别对31份供试材料进行PCR检测,以能扩增出相应目标片段大小的品种为含有该抗性基因,统计扩增条带。因Pikm基因由紧密连锁具有独立功能的两个NBS-LRR类基因组成,分别用引物DKm1和DKm2检测这两个基因,以引物DKm1和DKm2都能检测到相应目的片段为含pikm基因。研究表明,全部供试材料中均无Pi21基因。Pi9、Pita、Pib和Pikm基因所占比例偏少,共检测到3个品种含pi9基因,分别为新稻21号、新稻23号和新稻24号,占全部供试材料的9.68%;7个品种含Pita基因,分别为新稻23号、新稻36号、新稻39号、新稻44号、伊粳8号、伊粳12号、阿稻1号,占全部供试材料的22.58%;2个品种含pib基因,分别为新稻1号、新稻23号,占全部供试材料的6.45%;4个品种含pikm基因,分别为新稻11号、新稻23号、新稻39号、伊粳13号,占全部供试材料的12.90%。全部供试材料中Pi2、Pigm、Pil、Pikh/Pi54、Pi63、Pia、Pid2、Pid3/Pi25基因广泛存在。共检测到21个品种含有Pi2基因,占全部供试材料的67.74%;24个品种含有Pigm基因,占全部供试材料的77.42%;28个品种含有Pil基因,占全部供试材料的90.32%;24个品种含有Pikh/Pi54基因,占全部供试材料的77.42%;30个品种含有Pi63基因,占全部供试材料的96.77%;31个品种含有Pia基因,占全部供试材料的100%;29个品种含有Pid2基因,占全部供试材料的93.55%;31个品种含有Pid3/Pi25基因,占全部供试材料的100%。结果表明,新疆部分自育品种表现具有不同的广谱抗性基因外,同时都具有生理小种特异性抗性基因。图1,表4
表3 引物名称及相关序列
Table 3 Name and sequences of specific primers used for PCR
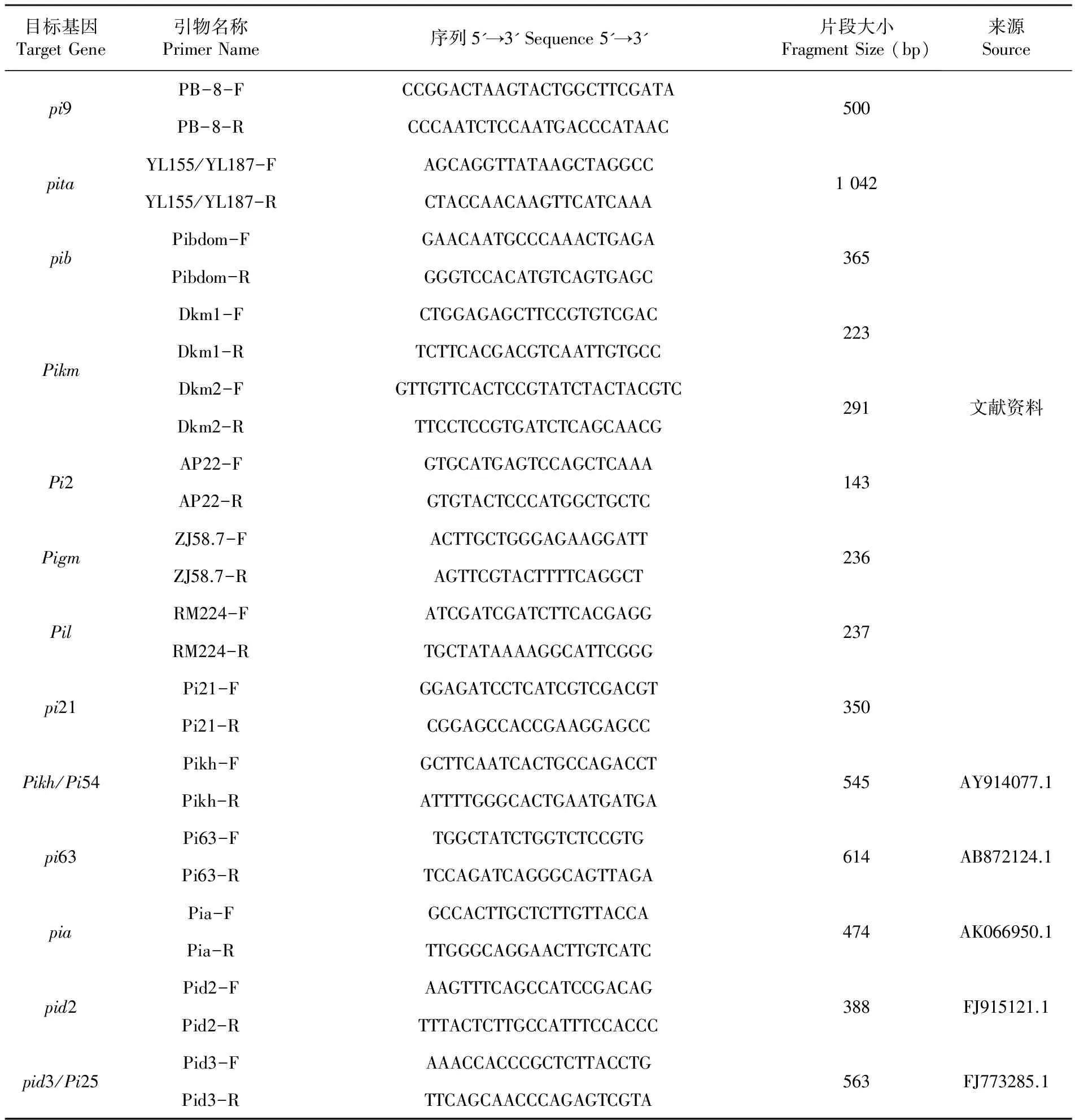
目标基因TargetGene引物名称PrimerName序列5'→3'Sequence5'→3'片段大小FragmentSize(bp)来源Sourcepi9pitapibPikmPi2PigmPilpi21PB-8-FCCGGACTAAGTACTGGCTTCGATAPB-8-RCCCAATCTCCAATGACCCATAACYL155/YL187-FAGCAGGTTATAAGCTAGGCCYL155/YL187-RCTACCAACAAGTTCATCAAAPibdom-FGAACAATGCCCAAACTGAGAPibdom-RGGGTCCACATGTCAGTGAGCDkm1-FCTGGAGAGCTTCCGTGTCGACDkm1-RTCTTCACGACGTCAATTGTGCCDkm2-FGTTGTTCACTCCGTATCTACTACGTCDkm2-RTTCCTCCGTGATCTCAGCAACGAP22-FGTGCATGAGTCCAGCTCAAAAP22-RGTGTACTCCCATGGCTGCTCZJ58.7-FACTTGCTGGGAGAAGGATTZJ58.7-RAGTTCGTACTTTTCAGGCTRM224-FATCGATCGATCTTCACGAGGRM224-RTGCTATAAAAGGCATTCGGGPi21-FGGAGATCCTCATCGTCGACGTPi21-RCGGAGCCACCGAAGGAGCC5001042365223291143236237350文献资料Pikh/Pi54Pikh-FGCTTCAATCACTGCCAGACCTPikh-RATTTTGGGCACTGAATGATGA545AY914077.1pi63Pi63-FTGGCTATCTGGTCTCCGTGPi63-RTCCAGATCAGGGCAGTTAGA614AB872124.1piaPia-FGCCACTTGCTCTTGTTACCAPia-RTTGGGCAGGAACTTGTCATC474AK066950.1pid2Pid2-FAAGTTTCAGCCATCCGACAGPid2-RTTTACTCTTGCCATTTCCACCC388FJ915121.1pid3/Pi25Pid3-FAAACCACCCGCTCTTACCTGPid3-RTTCAGCAACCCAGAGTCGTA563FJ773285.1
2.2 供试材料分子标记检测
31份供试材料不同品种抗性基因数量分布在5~11个,含5个抗性基因的为新稻12号;含6个抗性基因的为新稻19号、新稻21号、新稻30号、新稻35号和新稻42号;含7个抗性基因的为新稻10号、新稻14号、新稻17号、新稻24号、新稻27号、新稻34号、新稻41号、A稻7号和A稻8号;含8个抗性基因的为新稻11号、新稻29号、新稻33号、新稻44号、新稻45号、A稻4号、伊粳12号、伊粳13号、阿稻1号、粮粳5号和粮香5号;含9个抗性基因的为新稻1号、新稻36号、伊粳8号;含10个抗性基因的为新稻39号;含11个抗性基因的为新稻23号。表4
表4 供试材料分子标记检测结果
Table 4 Results of molecular marker analysis of plant materials in the study

编号No.品种VarietiesPi9PitaPibPikmPi2PigmPilPi21Pikh/Pi54Pi63PiaPid2Pid3/Pi25共计Total1新稻1号--+-+++-+++++92新稻10号----+-+-+++++73新稻11号---++-+-+++++84新稻12号------+--++++55新稻14号-----++-+++++76新稻17号----+++-+++-+77新稻19号------+-+++++68新稻21号+-----+-+++++69新稻23号+++++++-+++-+1110新稻24号+---+-+--++++711新稻27号----+-+-+++++712新稻29号----+++-+++++813新稻30号-----++--++++614新稻33号----+++-+++++815新稻34号-----++-+++++716新稻35号----++---++++617新稻36号-+--+++-+++++918新稻39号-+-++++-+++++1019新稻41号-----++-+++++720新稻42号----+++---+++621新稻44号-+--+++--++++822新稻45号----+++-+++++823A稻4号----+++-+++++824A稻7号----++--+++++725A稻8号-----+--+++++726伊粳8号-+--+++-+++++927伊粳12号-+---++-+++++828伊粳13号---+-++-+++++829阿稻1号-+--+++--++++830粮粳5号----+++-+++++831粮香5号----+++-+++++8
注:“+”表示含该基因;“-”表示不含该基因
Note:“+”show that the gene was contained; “-”show that the gene was not contained
注:I:Pi9基因扩增结果;II:Pid2基因扩增结果;III:Pid3/Pi25基因扩增结果。M:Markers;1:新稻1号;2:新稻10号;3:新稻11号;4:新稻12号;5:新稻14号;6:新稻17号;7:新稻19号;8:新稻21号;9:新稻23号;10:新稻24号;11:新稻27号;12:新稻29号;13:新稻30号;14:新稻33号;15:新稻34号;16:新稻35号;17:新稻36号;18:新稻39号;19:新稻41号;20:新稻42号;21:新稻44号;22:新稻45号;23:A稻4号;24:A稻7号;25:A稻8号;26:伊粳8号;27:伊粳12号;28:伊粳13号;29:阿稻1号;30:粮粳5号;31:粮香5号
Note: I: PCR amplification ofPi9 gene; II: PCR amplification ofPid2 gene;III: PCR amplification ofPid3/Pi25 gene. M: Markers; 1: Xindao1; 2: Xindao10; 3: Xindao11; 4: Xindao12; 5: Xindao14; 6: Xindao17; 7: Xindao19; 8: Xindao21; 9: Xindao23; 10: Xindao24; 11: Xindao27; 12: Xindao29; 13: Xindao30; 14: Xindao33; 15: Xindao34; 16: Xindao35; 17: Xindao36; 18: Xindao39; 19: Xindao41; 20: Xindao42; 21: Xindao44; 22: Xindao45; 23: Adao4; 24: Adao7; 25: Adao8; 26: Yijing8; 27: Yijing12; 28: Yijing13; 29: Adao1; 30: Liangjing5; 31: Liangxiang5
图1 水稻品种部分抗性基因扩增结果
Fig.1 PCR amplification of part of resistance genes in rice cultivars
3 讨 论
因稻瘟病兼具分布广泛、发病频率高、危害巨大及生理小种复杂等特点,世界各主要产稻国家一直十分重视发掘抗谱广、抗性强而持久的种质资源。日本完成2 878个主栽品种的抗性基因型分析鉴定工作,在商业品种中导入不同主要抗性基因后,携带多个不同抗性基因的多系品种也已育成,为持续有效的控制日本稻瘟病的流行发生奠定了良好基础[24]。我国自20世纪80年代以来,也开展了这方面研究,大致经历了系统育种、系统育种与杂交育种、多种途径并行及常规育种与分子辅助育种相结合的四个抗病育种阶段[25]。但目前还是多限于一个或几个抗源品种的研究,大量主栽品种的抗性基因型分析涉及程度仍不够深入全面,致使国内主栽品种的利用与布局依旧存在较大的盲目性。
抗病基因的发掘与利用是抗病育种的基础和核心。只有了解品种本身的抗性基因型,才能有针对性的改良品种抗性,进而在时间和空间上合理的布局抗病良种。依靠表型选择的传统抗性追踪方法,需大量的接种鉴定、抗性遗传和基因等位性分析工作,且在不同稻区及不同年代,病原菌优势小种组成常发生动态变化,品种的稻瘟病抗性往往出现较大的差异性,所以常因小种专化性和发病条件不充分等因素影响而产生选择不确定性,很难满足现代抗病育种的需要。随着DNA分子标记技术的发展,分子标记辅助选择育种的有效利用,尤其是抗性基因本身序列分子标记进行辅助选择,大幅提高了抗源筛选、抗性基因鉴定及育种选择的效率和准确性。可缩短育种时间并降低成本,是快速、精确选择抗性基因的有效途径[15]。
Pi9、Pita、Pib、Pikm、Pi2、Pigm、Pil、Pikh/Pi54基因具有广谱抗性,在水稻稻瘟病抗性育种中应用广泛[5]。结果表明,新疆选育供试材料中Pi9、Pita、Pib和Pikm基因所占比例偏少。Pi9在新疆选育水稻种质资源中所占比例很少的现象与张羽等[19]发现陕西省主要水稻种质资源中Pi9所占比例较少的现象相似,可能缘于Pi9基因本身来源于野生稻。共检测到3个品种含pi9基因,7个品种含pita基因,2个品种含pib基因,4个品种含pikm基因。全部新疆选育供试材料中均无Pi21基因存在。Pi2、Pigm、Pil、Pikh/Pi54、Pi63基因分布广泛,依次检测到的品种数为21、24、28、24和30个。表现为生理小种特异性抗性的Pid2、Pia、Pid3/Pi25基因,在新疆选育水稻种质资源中依次检测到的品种数为29(除新稻17号和新稻23号)、31和31个,分布范围达到93.5%~100%,初步鉴定结果表明,新疆选育水稻种质资源中对无毒菌株(小种)ZB15、B90002和Zhong-10-8-14的特异性抗性水平很高。全部供试材料不同品种抗性基因数量分布在5~11个,其中含11个抗性基因的新稻23号,其聚合了除Pi21和Pid2外其余所有抗性基因,生产实践中是属抗病品种,具有一定的光谱抗性,因此,在培育抗病新品种上可以加以利用。
目前所定位或克隆的抗性基因多源自野生稻资源或地方品种,育种实践中不利基因连锁累赘的弊端,致使这些抗性基因很难被直接利用,无法满足不同生态区域抗病育种的目标要求。通过杂交聚合手段、花药培养早代纯合方法和分子标记辅助选择来选育并聚合多个抗病基因于同一品种,是减缓和规避稻种抗性丧失的最佳途径[13,26-27]。同一稻种经多个抗性基因聚合后,抗谱被拓宽且提升了对一些生理小种的抗性水平,该手段并不是单个抗病基因抗谱间的简单累加,而是表现为抗性基因之间发生极显著的基因互作关系[28-30]。进行抗性基因聚合育种的前提是明确抗性资源的基因型背景,关键是提高选择效率。
4 结 论
新疆主要育成的31份水稻品种中均无Pi21基因存在,表现为广谱抗性的Pi9、Pita、Pib和Pikm基因所占比例偏少,分别占9.68%、22.58%、6.45%、12.90%;表现为广谱抗性的Pi2、Pigm、Pil、Pikh/Pi54、Pi63和表现为生理小种特异性抗性的Pia、Pid2、Pid3/Pi25基因广泛存在,所占比率67.74%~100%;不同水稻品种抗性基因数量分布在5~11个,含11个抗性基因的品种是新稻23号,其聚合了除Pi21和Pid2外其余所有抗性基因。新疆主要水稻育成品种中广泛存在具有光谱抗性的Pi2、Pigm、Pil、Pikh/Pi54、Pi63和表现为生理小种特异性抗性的Pia、Pid2、Pid3/Pi25基因,利用这些基因的分子标记可以有效快速鉴定品种中的抗性基因的存在和分布情况。
References)
[1] 温小红,谢明杰,姜健,等.水稻稻瘟病防治方法研究进展[J].中国农学通报,2013,29(3):190-195.
WEN Xiao-hong,XIE Ming -jie,JIANG Jian,et al. (2013). Advances in research on control method of rice blast [J].ChineseAgriculturalScienceBulletin,29(3):190-195. (in Chinese)
[2] 黄富,刘成元,谢戎,等.水稻种质资源抗稻瘟病及其恢保特性评价[J].中国农业科学,2005,38 (4):831-836.
HUANG Fu,LIU Cheng -yuan,XIE Rong,et al. (2005) .Evaluation of blast resistance and restoring or remaining characteristics of rice germplasm resources [J].ScientiaAgriculturaSinica,38 (4):831-836. (in Chinese)
[3] 孟军,赵明富,何月秋.稻瘟病菌重组研究概要[J].江西农业大学学报,2005,27 (3):422-424.
MENG Jun,ZHAO Ming-fu,HE Yue-qiu. (2005) . Brief review of the recombination in magnaporthe grisea [J].ActaAgriculturaeUniversitatisJiangxiensis,27 (3):422-424. (in Chinese)
[4] 袁筱萍,魏兴华,余汉勇,等.部分中国栽培稻资源对稻瘟病的抗性分析[J].植物保护,2005,31(3):27-31.
YUAN Xiao- ping,WEI Xing -hua,YU Han-yong,et al. (2005) .Analysis of the resistance reaction of some Chinese rice germplasms(OryzasativaL.) to rice blast[J].PlantProtection,31 (3):27-31. (in Chinese)
[5] 何秀英,王玲,吴伟怀,等.水稻稻瘟病抗性基因的定位、克隆及育种应用研究进展[J]. 中国农学通报,2014,30(6):1-12.
HE Xiu - ying,WANG Ling,WU Wei-huai,et al. (2014) .The progress of mapping,isolation of the genes resisting to blast andtheir breeding application in rice[J].ChineseAgriculturalScienceBulletin,30(6):1-12. (in Chinese)
[6] Deng, Y., Zhu, X., Xu, J., Chen, H., & He, Z. (2009).Map-BasedCloningandBreedingApplicationofaBroad-SpectrumResistanceGenePigmtoRiceBlast.AdvancesinGenetics,GenomicsandControlofRiceBlastDisease.
[7] Qu, S., Liu, G. B., Bellizzi, M., Zeng, L., Dai, L., & Han, B., et al. (2006). The broad-spectrum blast resistance gene pi9 encodes a nucleotide-binding site-leucine-rich repeat protein and is a member of a multigene family in rice.Genetics, 172(3):1,901-1,914.
[8] Ashikawa, I., Hayashi, N., Yamane, H., Kanamori, H., Wu, J., & Matsumoto, T., et al. (2008). Two adjacent nucleotide-binding site-leucine-rich repeat class genes are required to confer Pikm-specific rice blast resistance.Genetics, 180(4):2,267-2,276.
[9] Sharma, T. R., Madhav, M. S., Singh, B. K., Shanker, P., Jana, T. K., & Dalal, V., et al. (2005). High-resolution mapping, cloning and molecular characterization of the pi - k, h, gene of rice, which confers resistance to magnaporthe grisea.MolecularGenetics&Genomics, 274(6): 569-578.
[10] Jiang, H., Feng, Y., Bao, L., Li, X., Gao, G., & Zhang, Q., et al. (2012). Improving blast resistance of Jin 23B and its hybrid rice by marker-assisted gene pyramiding.MolecularBreeding,30(4):1,679-1,688.
[11] Hua, L., Wu, J., Chen, C., Wu, W., He, X., & Lin, F., et al. (2012). The isolation ofpi1, an allele at the pik locus which confers broad spectrum resistance to rice blast.Tag.theoretical&AppliedGenetics.theoretischeUndAngewandteGenetik, 125(5):1,047-1,055.
[12] Liang-Ying, Xiong-Lun, Ying-Hui, Xiao, Guo-Liang, & Wang. (2007). Recent advances in cloning and characterization of disease resistance genes in rice.JournalofIntegrativePlantBiology, 49(1):112-119.
[13] 陈学伟,李仕贵,马玉清,等.水稻抗稻瘟病基因pi-d(t)1、Pi-b、Pi-ta2的聚合及分子标记选择[J]. 生物工程学报,2004,20 (5):708-714.
CHEN Xue-wei,LI Shi - gui,MA Yu-qing,et al. (2004) .Marker-assisted selection and pyramiding for three blast resistance genespi-d(t)1、Pi-b、Pi-ta2,in rice [J].ChineseJournalofBiotechnology,20(5):708-714. (in Chinese)
[14] 姚姝,刘燕清,张亚东,等. 水稻抗稻瘟病基因Pi-ta和Pi-b多重PCR体系的构建与应用[J].作物学报,2014,40(9):2 225-2 230.
YAO Shu,LIU Yan-qing,ZHANG Ya-dong,et al. (2014) .Establishment and application of multiplex PCR system for blast resistance genesPi-taandPi-bin rice[J].ActaAgronomicaSinica,40(9):2,225-2,230.(in Chinese)
[15] 时克,雷财林,程治军,等. 稻瘟病抗性基因Pita和Pib在我国水稻主栽品种中的分布[J].植物遗传资源学报,2009,10 (1):21-26.
SHI Ke,LEI Cai -lin,CHENG Zhi-jun,et al. (2009). Distribution of two blast resistance genesPitaandPibin major rice cultivars in China [J].JournalofPlantGeneticResources, 10(1):21-26. (in Chinese)
[16] 杨杰,杨金欢,王军,等. 稻瘟病抗病基因Pita和Pib在中国水稻地方品种中的分布[J].华北农学报,2011,26(3):1-6.
YANG Jie,YANG Jin - huan,WANG Jun,et al. (2011) . Distribution of two blast resistant genesPitaandPibin landrace rice in China[J].ActaAgricBorealiSin,26(3):1-6. (in Chinese)
[17] 王忠华,贾育林,吴殿星,等.水稻抗稻瘟病基因Pi-ta的分子标记辅助选择[J].作物学报,2004,30 (12):1 259-1 265.
WANG Zhong -hua,JIA Yu-lin,WU Dian-xing,et al. (2004) .Molecular markers-assisted selection of the rice blast resistance genePi-ta[J].ActaAgronomicaSinica,30(12):1,259-1,265.(in Chinese)
[18] 刘洋,徐培洲,张红宇,等.水稻抗稻瘟病Pib基因的分子标记辅助选择与应用[J] .中国农业科学,2008,41(1):9-14.
LIU Yang,XU Pei-zhou,ZHANG Hong-yu,et al. (2008) .Marker-assisted selection and application of blast resistant genePibin rice [J].ScientiaAgriculturaSinica,41(1):9-14.(in Chinese)
[19] 张羽,冯志峰,张晗,等.陕西省水稻种质资源中Pi9基因的分布状况[J] .四川农业大学学报,2013,31(2):115-121.
ZHANG Yu,FENG Zhi-feng,ZHANG Han,et al. (2013) . Distribution of the blast resistance genePi9 in rice germplasm resources in Shanxi Province [J] .JournalofSichuanAgriculturalUniversity,31(2):115-121.(in Chinese)
[20] 李进斌,王甜,许明辉. 云南地方稻种抗稻瘟病基因Pi-ta和Pi-b的鉴定[J]. 中国水稻科学,2012,26(5):593-599.
LI Jin-bin,WANG Tian,XU Ming-hui. (2012) .Identification ofPi-taandPi-bgenes for rice blast resistance of rice landraces from Yunnan Province[J].ChineseJournalofRiceScience,26(5):593-599. (in Chinese)
[21] Jia, Y. L., Wang, Z. H., & Singh, P. (2003). Development of dominant rice blast pi-ta resistance gene markers. crop sci 42: 2145-2149.CropScience, 42(6): 2,145-2,149.
[22] Jia, Y., Wang, Z., Fjellstrom, R. G., Moldenhauer, K. A., Azam, M. A., & Correll, J., et al. (2004). Rice pi-ta gene confers resistance to the major pathotypes of the rice blast fungus in the united states.Phytopathology,94(3): 296-301.
[23] Fjellstrom, R., Conawaybormans, C. A., Mcclung, A. M., Marchetti, M. A., Shank, A. R., & Park, W. D. (2004). Development of dna markers sui
Table for marker assisted selection of three pi genes conferring resistance to multiple pyricularia grisea pathotypes.Cropence, 44(5):1,790-1,798.
[24] Koizumi, S., Ashizawa, T., & Zenbayashi, K. S. (2004).DurableControlofRiceBlastDiseasewithMultilines.RiceBlast:InteractionwithRiceandControl. Springer Netherlands.
[25] Liang-Ying, Xiong-Lun, Ying-Hui, Xiao, Guo-Liang, & Wang. (2007). Recent advances in cloning and characterization of disease resistance genes in rice.JournalofIntegrativePlantBiology, 49(1):112-119.
[26] 张祎颖,洪汝科,张锦文,等. 花药培养获得优质抗稻瘟病水稻[J]. 西南农业学报,2008,21 (1):75-79.
ZHANG Yi-ying,HONG Ru-ke,ZHANG Jin - wen,et al. (2008) .High quality and blast resistance DH lines via anther culture[J].SouthwestChinaJournalofAgriculturalSciences,21(1):75-79. (in Chinese)
[27] 孙海波,邹美智,任洪岩,等. 花药培养快速培育聚合抗3种水稻病害基因的新种质研究[J]. 华北农学报,2015,30(2):116 -123.
SUN Hai-bo,ZOU Mei-zhi,REN Hong-yan,et al. (2015) .Study of quick cultivation of new seeds with three diseases resistance by anther culture method[J].ActaAgriculturaeBoreali-Sinica,30(2):116-123. (in Chinese)
[28] 马军韬,张国民,辛爱华,等. 水稻品种抗稻瘟病分析及基因聚合抗性改良[J]. 植物保护学报, 2016,43(2): 177-183.
MA Jun-tao,ZHANG Guo-min,XIN Ai-hua,et al. (2016) .Resistance analysis and improvement of rice varieties by gene pyramiding[J].JournalofPlantProtection,43(2):177-183. (in Chinese)
[29] Hur, Y. J., Cho, J. H., Park, H. S., Noh, T. H., Park, D. S., & Ji, Y. L., et al. (2016). Pyramiding of two rice bacterial blight resistance genes, xa3, and xa4, and a closely linked cold-tolerance qtl on chromosome 11.Theoretical&AppliedGenetics, 129(10):1,861-1,871.
[30] Korinsak, S., Siangliw, M., Kotcharerk, J., Jairin, J., Siangliw, J. L., & Jongdee, B., et al. (2016). Improvement of the submergence tolerance and the brown planthopper resistance of the thai jasmine rice cultivar kdml105 by pyramiding sub1, and qbph12.FieldCropsResearch, 188(8):105-112.
Abstract:【Objective】 To make clear the distribution situation of rice blast resistance genes from the main bred rice varieties in Xinjiang and provide the basis for rice blast resistance breeding.【Method】In this study, 31 Xinjiang bred rice varieties were taken as materials, specific molecular markers of 13 rice blast resistance genes were used to carry out PCR identification, thus preliminarily establishing the distribution situation of resistance genes from Xinjiang main bred varieties.【Result】There were noPi21 genes in all the test materials; The proportion ofPi9,Pita,PibandPikmwere less, which belong to broad spectrum resistance genes, accounting for 9.68%, 22.58%, 6.45% and 12.90%, respectively. However,Pi2,Pigm,Pil,Pikh/Pi54 andPi63 that belong to broad spectrum resistance genes andPia,Pid2,Pid3/Pi25 that belong to physiological races specific resistance genes, all existed widely between 67.74% and 100%. The number of blast resistance genes in different varieties were between 5 and 11. Xindao 23 contained 11blast resistance genes that polymerized all blast resistance genes except forPi21 andPid2.【Conclusion】Pi2,Pigm,Pil,Pikh/Pi54 andPi63 that belong to broad spectrum blast resistance genes andPia,Pid2 andPid3/Pi25 that belong to physiological races specific resistance genes were widely spread in partial bred rice varieties in Xinjiang. The presence and distribution situation of blast resistance genes in varieties can be identified rapidly and efficiently by using specific molecular markers.
Keywords: rice; bred varieties; rice blast; resistance genes
Distributionof13RiceBlastResistanceGenesfromBredVarietiesinXinjiang
ZHANG Yan-hong1, JIA Chun-ping1, WEN Xiao-rong2, TANG Fu-sen2,WANG Feng-bin1, ZHU Xiao-xia1,2
(1.InstituteofNuclearandBiologicalTechnologies,XinjiangAcademyofAgriculturalSciences,Urumqi830091,China; 2.RiceExperimentStationinWensu,XinjiangAcademyofAgriculturalSciences,WensuXinjiang8430091,China)
10.6048/j.issn.1001-4330.2017.09.004
S511
A
1001-4330(2017)09-1595-11
2017-07-17
科技部“十三五”七大农作物育种项目课题(2017YFD0100505);新疆农科院青年基金项目(xjnkq-2015034)
张燕红(1982-),女,助理研究员,硕士,研究方向为水稻遗传育种及栽培,(E-mail)zhangyanhong9527@163.com
王奉斌(1968-),男,研究员,研究方向为水稻遗传育种及栽培,(E-mail)xjnkywfb@163.com
Supported by: The Ministry of Science and Technology of China, "Seven Crops Breeding Project during "the13th Five-year Plan" Period(2017YFD0100505)and The Youth Fund Project of Xinjiang Academy of Agricultural Sciences (xjnkq-2015034)
Corresponding author:WANG Feng-bin(1968-),male, researcher. Research area: Genetic breeding and cultivation techniques of rice. (E-mail) xjnkywfb@163.com
猜你喜欢
智慧农业导刊(2022年21期)2022-11-03
河北科技师范学院学报(2022年2期)2022-08-26
今日农业(2022年4期)2022-06-01
作物学报(2022年6期)2022-04-08
空间科学学报(2021年1期)2021-05-22
农民文摘(2019年7期)2019-01-14
浙江农业学报(2017年1期)2017-05-17
上海农业学报(2017年3期)2017-04-10
中国果菜(2016年9期)2016-03-01
中国蔬菜(2015年9期)2015-12-21
