
谷子Oleosin基因家族及其对干旱响应的分析
2018-01-17 06:14蒋茂双元香梅刘晓东候蕊苏彦冰郭展于世慧李红英
山西农业大学学报(自然科学版) 2018年1期
蒋茂双,元香梅,刘晓东,候蕊,苏彦冰,郭展,于世慧,李红英,2*
(1.山西农业大学 农学院,山西 太谷 030801;2.杂粮种质资源发掘与遗传改良山西省重点实验室,山西 太谷 030801;3.山西省农业科学院 作物科学研究所 山西 太原 030031;4.山西省晋城市城区农业委员会,山西 晋城 048000)
谷子是一种耐旱,耐瘠薄作物,在干旱、半干旱地区占有重要地位[1],在节水农业和旱作农业中具有重要意义。谷子因具有基因组小(490 M)、生育期短、与其它主粮作物亲缘关系密切等特点,已逐渐成为禾本科基因组学研究的理想模式植物[2]。且谷子具有较强的适应性和非生物胁迫耐受性,其相关基因的挖掘具有很深的潜力。近年来,人们从形态学、产量、生理生化等方面对谷子进行了抗旱鉴定、筛选和抗旱育种,取得了较好的成果[3]。随着谷子全基因组测序工作的完成,测序成本的降低,人们对谷子抗旱分子机制的相关研究也取得了一定进展,为谷子分子育种提供了参考。但谷子抗旱是作物与复杂环境相互作用的结果,也是多基因网络调控的结果[4],尚有许多抗旱基因有待挖掘,谷子抗旱的调控机制还有待完善。
Oleosin最早在芥菜中发现,随后不同树木(柑橘、油棕、可可等)、油料作物(芝麻、油菜、大豆、花生、向日葵等)以及其它植物(拟南芥、玉米、水稻等)的Oleosin基因序列和氨基酸序列陆续报道[5,6]。有研究表明Oleosin基因受水分逆境、茉莉酮酸、ABA和渗透稳定剂诱导表达[5],可见该类基因对植物耐旱、抗旱具有重要意义。此外,王俊娟研究发现棉花在低温胁迫下,类似油质蛋白(15kDa Oleosin-like protein)基因上调表达,认为该基因参与增强棉花的抗逆性[7]。Milioni D等研究认为Oleosin参与植物热激响应[8]。Oleosin基因家族较小,但其生物学功能还未完全研究清楚[9],谷子的Oleosin基因研究相关报道较少,其在谷子抗旱中的功能还需进一步证明。为此,本研究通过比较谷子Oleosin基因在PEG模拟干旱胁迫下的表达差异,并进行生物信息学分析,初步探究Oleosin基因功能与谷子抗旱关系。
1 材料与方法
1.1 试验材料
抗旱谷子品种勾勾母鸡咀(GG),干旱敏感品种晋汾16(JF16),均由山西农业大学农业生物工程研究所提供。
供试材料种植于人工气候室,所用基质为营养土:蛭石=3∶1;培养条件设置为14 h光照28 ℃/10 h黑暗23 ℃。培养3周后,试验组、对照组分别采用20 % PEG-6000和蒸馏水处理0.5 h,然后2组各随机取3株(整株谷苗),立即放入液氮速冻,提取RNA并进行表达谱测序。
1.2 研究方法
1.2.1 谷子Oleosin基因在干旱胁迫下的相对表达量分析
在表达谱测序结果的数据中,干旱胁迫下Oleosin基因差异表达,根据表达情况绘制GG和JF16两个品种在干旱、对照条件下的表达柱形图。
1.2.2 谷子Oleosin家族基因基本信息及理化性质分析
利用数据库Phytozome12.1,选取物种为谷子(Setaria italica v2.2)通过关键词“Oleosin”搜索并整理谷子Oleosin基因基本信息,并结合DNAMAN软件,分析Oleosin蛋白的理化性质。
1.2.3 基因结构与进化树分析
利用GSDS在线工具(http://gsds.cbi.pku.edu.cn/)制作谷子Oleosin家族基因外显子-内含子结构示意图。
利用注释基因Seita.9G579700(Si037 924m.g)在NCBI数据库BLAST,选取拟南芥、水稻、玉米、谷子中同源性高的Oleosin基因,并获取氨基酸序列,另存为fasta格式,用MEGA7.0进行多序列比对分析,建树方法选择NJ法(Neighobr-Joining Tree),Bootstrap值设置为1000,其他参数选择默认,构建系统发育树。
1.2.4 启动子顺式作用元件分析
从基因组数据库 Phytozome12.1查找谷子Oleosin基因序列信息,选取起始密码子上游3 000 bp序列作为启动子区,在PlantCARE主页点击“Search for CARE”,按要求输入相关信息,进行启动子元件预测。
2 结果与分析
2.1 谷子Oleosin基因的表达谱分析
为了解Oleosin基因对干旱胁迫的响应,对谷子幼苗进行干旱处理,通过表达谱测序比较干旱与对照的表达量变化。如图1所示,PEG模拟干旱胁迫下,在抗旱品种GG及干旱敏感品种JF16中,Oleosin表达量均显著上调,其中该基因在GG中上调到133倍,JF16中上调到944倍,说明该基因对干旱具有明显的响应。同时,在对照组,该基因在GG与JF16中的表达量相同,均为0.01,但在干旱胁迫下,干旱敏感品种JF16中的表达量比抗旱品种GG高7倍,说明Oleosin基因对干旱比较敏感,且受品种差异的影响较大。

图1 GG与JF16品种中Oleosin基因在PEG胁迫下的表达量Fig.1 Expression of Oleosin gene in GG and JF16 under the PEG stress
2.2 谷子Oleosin基因家族成员识别及其基本信息
通过Phytozome 12.1数据库检索,谷子中的Oleosin基因家族较小,仅有6个成员。其中1个在干旱表达谱测序结果中上调表达。这些家族成员中基因序列长度相差较大,最短的441 bp,较长的2个分别是951 bp、918 bp,而它们的CDS序列长度比较相近,在441~552 bp之间。这些成员中有2个基因在9号染色体上,即Seita.9G579700与Seita.9G128500。6个Oleosin蛋白分子量均较小,在15~18 kDa,氨基酸序列长度较短,且相近,在146~183 bp之间。其等电点范围在8.20~10.45之间(表1)。
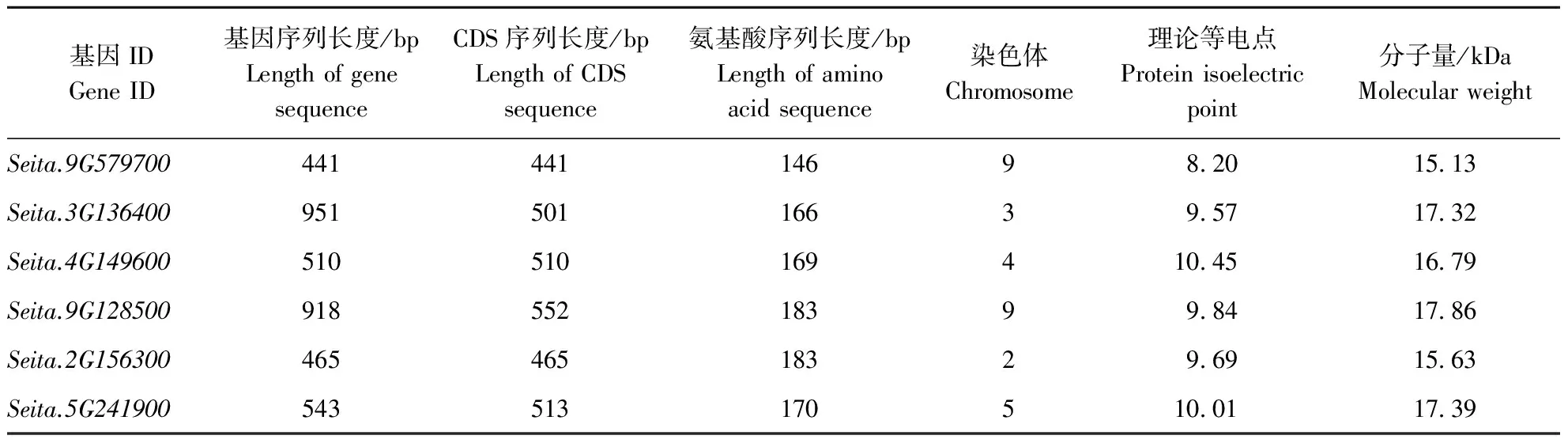
表1 谷子Oleosin基因家族的基本信息Table 1 Basic information of Oleosin gene family in foxtail millet
2.3 基因结构分析
内含子-外显子结构分析显示,仅Seita.5G241900有1个内含子,且只有该基因有2个外显子,呈现断裂基因特征;其他5个基因结构简单,均是只有1个外显子的连续编码区,表明该家族成员间基因结构差异较小。Seita.3G136400和Seita.9G128500具有非编码区(图2)。

图2 谷子Oleosin 基因内含子-外显子结构Fig.2 Intron-exon of Oleosin genes in foxtail millet
2.4 部分物种Oleosin蛋白的进化分析
将拟南芥、谷子、玉米、水稻、高粱中10个Oleosin基因进行系统进化分析,这10个基因被划分为3类,其中谷子中Seita.2G156300与水稻、玉米聚为一类,Seita.9G579700与高粱、玉米、水稻聚为一类,而拟南芥中的3个基因聚为一类,表明同是C4作物的谷子、高粱、玉米中的Oleosin基因亲缘关系较近,而与双子叶植物拟南芥亲缘关系较远。且单、双子叶植物间的亲缘关系较远。但谷子中的2个Oleosin基因却分别聚到2个不同的分支(图3)。

图3 谷子Oleosin蛋白与其它植物(拟南芥、水稻、高粱、玉米)Oleosin蛋白的系统进化树Fig.3 Phylogenetic tree of Oleosin proteins in foxtail millet and other plants (arabidopsis,rice,sorghum,maize)注:Seita,AT,LOC Os,So bic,Zm分别表示谷子、拟南芥、水稻、高粱、玉米Note:Seita,AT,LOC Os,So bic Zm respectively represent foxtail millet,arabidopsis,rice,sorghum,maize
2.5 启动子顺式作用元件分析
目前研究较多的抗旱相关顺式作用元件主要是ABRE (ABA-responsive element)、DRE (dehydration responsive element)、LTRE (low tem-perature responsive element)、MYB 和 MYC 元件。此外还有W-box、G-box中的一些启动子元件[10]、ABRE、MBS、MeJA等对植物非生物胁迫和生长发育调节均具有一定作用[11]。本研究对起始密码子上游3 000 bp的序列进行启动子元件分析(表2)。结果发现Oleosin家族基因启动子区域均含有ABA、MeJA响应元件以及MYB结合位点。其中与ABA响应相关的元件最少3个,最多的是Seita.9G579700有9个;MeJA响应相关的元件为2~14个;MYB结合位点至少1个;这些元件均与干旱、ABA诱导表达有关。6个成员也均含有较多的(13~25个)与光响应相关的元件。该家族中的Seita.3G136400、Seita.2G156300均有一个逆境与防卫相关元件TC-rich(defense and stress response)。此外,一些基因还具有赤霉素(GA)、水杨酸(SA)、低温(LTR)、热激(HSE)和根特异性(root specific)响应相关的元件。表明它们对干旱等非生物胁迫的响应具有一定作用。
3 结论与讨论
逆境诱导信号转导途径至少有3条:其中一条是依赖ABA的传导途径[12],而谷子Oleosin基因中均有与ABA响应的元件,说明Oleosin对ABA诱导抗旱具有重要作用,这与Buchanan 等对高粱中Oleosin的研究结果一致,该研究认为Oleosin能够改变膜渗透性,Oleosin蛋白对高粱响应ABA、高盐、水分等胁迫具有重要作用[13]。此外,Chen等人的研究发现用0.5 μmol·L-1ABA处理发芽的拟南芥,油质蛋白基因Ole2、Ole3均显著上调表达,证明Oleosin基因参与ABA信号通路的调节[14],也支撑了这一观点。
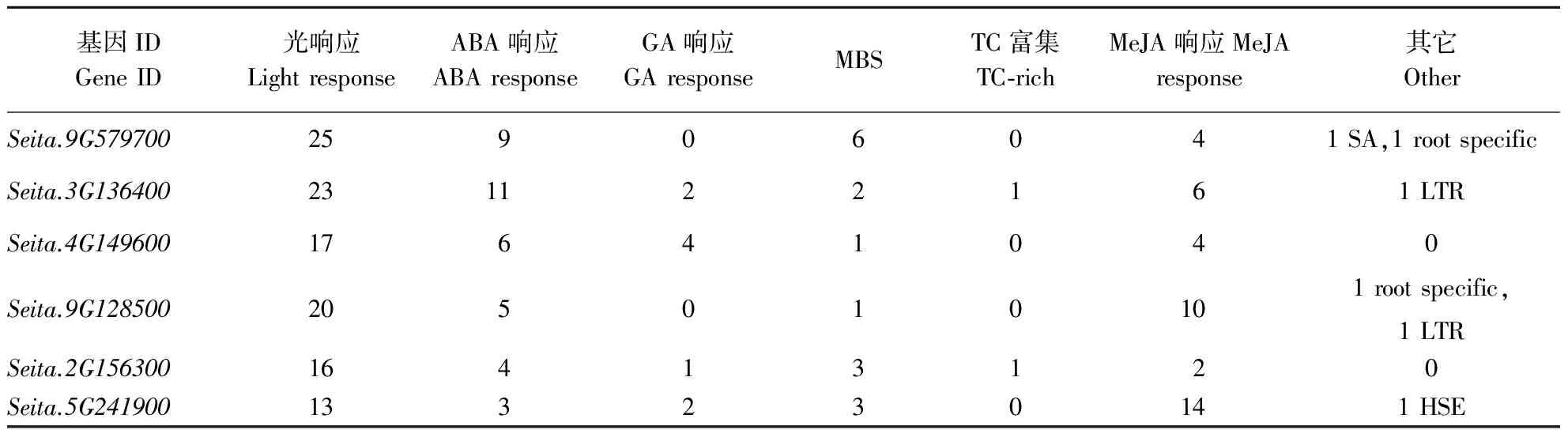
表2 谷子Oleosin基因启动子逆境相关顺式作用元件的数量分布Table 2 Distribution of cis-acting elements in foxtail millet Oleosin gene promoters
启动子元件分析发现谷子Oleosin家族的6个基因均含有MeJA响应元件,最多的是Seita.5G241900,有14个,其他最少的也有2个,而董桃杏等对水稻幼苗的抗旱生理效应研究发现茉莉酸甲酯一定程度上能缓解干旱胁迫对水稻幼苗的伤害,有效提高抗旱性[15],说明Oleosin可能受MeJA诱导参与植物抗旱调控。
有研究表明被子植物中的Oleosin基因多以基因家族的形式存在,一种植物中常有几个Oleosin异构体[6]。本研究谷子苗期干旱处理后仅发现一个Oleosin基因表达,且表达量较低,可能是由于其主要在种子中特异性表达[6,16],此次测序材料是全株幼苗,可能是根中表达的Oleosin[17]。谷子中的Oleosin基因,在进化树中被聚集到2个不同的分支,这可能是因为被子植物Oleosin蛋白具有2种明显不同的异构体[18],异构体的结构差异导致进化过程中亲缘关系较远,有待进一步研究证明。
综上分析,本研究初步确定Oleosin基因参与了谷子干旱胁迫的响应,此过程可能受ABA、MeJA的诱导。本文结果为谷子抗旱分子机制的研究和抗旱育种提供了理论依据,也为Oleosin基因的功能研究提供参考。
[1] 岳安良.谷子的抗旱性及其在旱作中的地位[J].河南农业科学,1987(5):3-4.
[2] 尹恒.谷子干旱下的转录分析及耐旱基因的发掘[D].大连:大连理工大学,2014.
[3] 张文英,黄建明,尉文彬.我国谷子抗旱性研究进展[J].安徽农业科学,2013,41(4):1469-1470.
[4] Lata C,Gupta S,Prasad M.Foxtail millet:a model crop for genetic and genomic studies in bioenergy grasses[J].Critical Reviews in Biotechnology,2013,33(3):328-343.
[5] Cao H.Genome-wide analysis ofoleosingene family in 22 tree species:an accelerator for metabolic engineering of bioFuel crops and agrigenomics industrial applications?[J].Omics A Journal of Integrative Biology,2015,19(9):521-541.
[6] 刘昱辉,贾士荣.植物油体表达体系的研究进展[J].农业生物技术学报,2003,11(5):531-537.
[7] 王俊娟.棉花抗冷性鉴定及相关基因的表达研究[D].北京:中国农业科学院,2016.
[8] Milioni D,Franz G,Sung R,et al.Gene expression during heat-shock in embryogenic carrot cell lines[J].Plant Cell Tissue and Organ Culture,2001,65(3):221-228.
[9] 眭顺照,祝钦泷,郑丽,等.植物蛋白Oleosin及其在植物基因工程中的应用[J].中国生物工程杂志,2003,23(6):17-21.
[10] 郭晋艳,郑晓瑜,邹翠霞,等.植物非生物胁迫诱导启动子顺式元件及转录因子研究进展[J].生物技术通报,2011(4):16-20,30.
[11] 元香梅,禾璐,张凯烨,等.谷子XTH基因家族与抗旱相关基因的分析[J].山西农业大学学报(自然科学版)2017,37(1):1-6.
[12] 康宗利,杨玉红,张立军.植物响应干旱胁迫的分子机制[J].玉米科学,2006,14(2):96-100.
[13] Buchanan C D,Lim S,Salzman R A,et al.Sorghum bicolor’s,transcriptome response to dehydration,high salinity and ABA[J].Plant Molecular Biology,2005,58(5):699-720.
[14] Chen C,Wu C,Miao J,et al.Arabidopsis SAG protein containing the MDN1 domain participates in seed germination and seedling development by negatively regulating ABI3 and ABI5[J].Journal of Experimental Botany,2014,65(1):35.
[15] 董桃杏,蔡昆争,张景欣,等.茉莉酸甲酯(MeJA)对水稻幼苗的抗旱生理效应[J].生态环境学报,2007,16(4):1261-1265.
[16] 鲁亚萍.花生Oleosin启动子和Arah1启动子功能鉴定及转录因子WRI1基因的in silico分析[D].泰安:山东农业大学,2012.
[17] Monica,Schmidt,Herman.Suppression of Soybean Oleosin Produces Micro-Oil Bodies that Aggregate into Oil Body/ER Complexes[J].Molecular plant,2008,1(6):910-924.
[18] Tzen J T C,Chuang R L C,Chen J C F,et al.Coexistence of both Oleosin isoforms on the surface of seed oil bodies and their individual stabilization to the organelles[J].Journal of Biochemistry,1998,123(2):318.
猜你喜欢
当代陕西(2021年21期)2022-01-19
今日农业(2021年14期)2021-11-25
山西师范大学学报(自然科学版)(2021年3期)2021-11-02
园艺与种苗(2020年12期)2021-01-08
今日农业(2020年13期)2020-12-15
河南水利年鉴(2020年0期)2020-06-09
建材发展导向(2019年11期)2019-08-24
延河(2018年6期)2018-06-11
红领巾·探索(2015年9期)2015-09-10
恋爱婚姻家庭·养生版(2011年8期)2011-05-14
