
微小RNA在抑郁症及其临床药物治疗中的研究进展
2018-10-19 01:43董宪喆朱维煜
解放军医学院学报 2018年9期
陈 超,董宪喆,胡 园,刘 旭,朱维煜,刘 屏
解放军总医院 临床药理研究室,北京 100853
微小RNA(microRNA,miRNA)是在真核生物中发现的一类长约22 nt,具有调控功能的内源性非编码RNA,可以通过序列互补与靶基因3'端非编码区域(3'UTRs)的miRNA反应原件(miRNA response elements,MREs)结合,使靶基因降解或抑制其翻译,进而抑制靶基因的水平。近年来的研究发现,目前超过半数的可检测miRNAs在哺乳动物神经系统中丰富表达。MiRNAs在神经元发育和分化、突触可塑性、认知和学习记忆功能等方面发挥重要作用,与多种中枢神经系统疾病(神经系统肿瘤、神经退行性疾病、精神科疾病等)的发生发展相关,在神经元的增殖、凋亡及抑郁症的发生中均扮演着重要的角色[1-4]。基于miRNAs的转录组学研究手段为发现抑郁症临床生物标记物和潜在的药物靶标提供了有力支持,本文就此进行综述。
1 MiRNAs参与抑郁症的病理生理过程
哺乳动物脑组织中的miRNA 种类及其表达水平都明显高于其他组织器官,脑中的miRNA种类约占所有已知miRNA的70%。MiRNA在神经发育过程中的神经祖细胞、神经元以及神经胶质细胞中都有特定的表达,可能参与了轴突导向、大脑突触可塑性调节等生理过程,以及神经退行性疾病和抑郁症的病理过程。尽管miRNA在神经退行性疾病中的作用已得到广泛的研究,但其在重度抑郁和其他精神疾病中作用的研究仍处于初级阶段。临床前的研究证据提示,miRNAs参与抑郁症的多个发病机制,尤其在突触可塑性、神经形成、信号传导通路关键性元件的基因表达调控等方面具有重要作用[1,5-6]。
1.1 MiRNAs与神经发生及突触可塑性 突触可塑性是学习和记忆的重要过程,其功能失调可能诱发神经系统紊乱。有多项研究显示了miRNA生物合成酶在突触可塑性中的作用,如兴奋性前脑神经元中的条件性Dicer敲除小鼠可致树突分枝细化减少和树突棘长度大幅增加[7-8]。某些在脑区内特异性富集的miRNAs也可调节突触可塑性,如miR-132、miR-134和miR-let-7影响神经元树突的形成,从而影响记忆功能和神经可塑性。MiR-124a和miR-125b则与轴突生长相关[9]。MiR-137与神经发生相关,主要调控神经干细胞生成成熟神经[10]。MiR-221和miR-222的表达被证实与细胞外调节蛋白激酶ERK1/2的激活有关[11]。
MiRNAs还可直接影响其他几种与抑郁相关的信号传导通路关键性元件的表达调控,如脑源性神经营养因子(BDNF)在抑郁症患者和应激性动物模型中减少[12],其可受miR-182、miR-30a-5p、miR-195多 个miRNAs调 控, 同时也可影响调控神经发生的miR-132的生成[13]。Smalheiser等[14]报道抑郁大鼠模型miR-22、miR-200b、miR-211、miR-300表达上调,这些miRNAs均靶向环磷腺苷效应元件结合蛋白(CREB)。MiR-124在脑中表达最为丰富,其功能失调与神经发生、神经免疫紊乱和中枢神经系统压力相关,是神经发生和其重要靶标CREB的关键调控分子[15]。
1.2 MiRNAs与单胺递质、压力性应答、糖皮质激素及昼夜节律 MiRNA可能通过调节5-羟色胺(5-HT)信号转导参与抑郁的病理生理过程。如miR-16已被证明可抑制蓝斑血清素转运蛋白(SERT)基因的表达,进而提高突触间隙内5-HT的水平[16-17];几种血清素受体也是miRNA的靶标,miR-96可增强5-HT1B受体表达和膜整合,miR-195可调节5-HT2A受体和5-HT4受体的表达水平[18-19]。
压力是抑郁症的主要危险因素之一,可导致下丘脑-垂体-肾上腺(HPA)轴活化、海马神经发生减少和海马突触可塑性受损。有研究表明,miR-218在小鼠前额叶皮质(PFC)锥体神经元中的上调与压力相关[20]。MiR-326作为一种压力诱导的神经肽urocortin1(Ucn1)的上游调节剂,其过表达可以在体外抑制Ucn1[21]。而长期和极度的压力亦可诱发糖皮质激素分泌紊乱,导致氧化应激和神经功能受损,miR-18和miR-124a均可下调糖皮质激素受体[22]。此外,miRNAs还可以调节影响情绪和诱发抑郁的昼夜节律紊乱,如miR-132可以调节活动依赖性神经可塑性和睡眠/觉醒周期,miR-182可靶向昼夜节律调节基因Clock,miR-192/194簇可以抑制PER基因家族[23-25]。
2 MiRNAs在抑郁症临床样本中的表达调控研究
已有多项研究证实了miRNAs表达水平改变与抑郁发病相关,最早的相关研究开始于2012年,主要采用基因芯片筛查加验证或候选miRNAs验证两种方法。这些研究通常采用抑郁患者尸脑组织、脑脊液、外周血、皮肤组织作为标本,可以在一定程度上揭示miRNA与抑郁之间的关系。本文对现有抑郁症临床样本的miRNA表达谱进行了总结(表1)。需要注意的是,尽管这些研究提示有大量与该疾病相关的miRNAs,但只有少数miRNAs重合,这有可能与该类精神性疾病的复杂病生理机制和基因多样性有关。此外,还要充分考虑到这些研究的局限性,包括有限的样本量,不同的临床表型和脑区的细胞异质性等。
2.1 尸脑组织相关的miRNAs研究 海马、杏仁核和前额叶皮质的整合和相互作用确保了大脑的认知、学习和记忆功能,许多临床研究和Meta分析验证了抑郁症患者大脑海马体积的减少、背外侧前额叶皮质活动的减少,额叶和其他脑区之间的突触连接受损[26]。但分子水平的证据还有限,miRNA在神经系统中的丰富表达及调控多靶点的特殊功能为研究抑郁症发病机制提供了新的切入点。
截至目前,已经有5项以重度抑郁患者尸脑组织作为研究样本的miRNA表达谱研究,这些研究通常以前额叶皮质作为起始研究材料。Smalheiser等[27-28]选取18例未服用抗抑郁药的自杀患者和17名正常对照者为研究对象,结果发现抑郁症自杀患者21种miRNAs表达下调,在这些miRNA的靶基因中,至少有3种与抑郁相关的基因(血管内皮生长因子A,B细胞淋巴瘤/白血病-2基因,DNA甲基转移酶)在前额叶皮层中被验证且共表达。该团队随后又对15例重症抑郁患者和15名正常对照者进行了芯片检测,发现miR-508-3p、miR-152-3p显著下调。Maussion等[29]以38例自杀患者(含23例重度抑郁)和17例正常对照为研究对象,发现自杀患者前额叶miR-185和miR-491-3p表达上调,并通过荧光素酶报告基因检测验证了miR-185的靶基因是酪氨酸激酶受体TrkB-T1。Lopez等[30-31]对15例抑郁自杀患者和16例对照者前额叶皮质筛选并验证了miR-34c-5p、miR-139-5p、miR-320c表达上调,通过对6个物种脑组织中miRNAs的比较,发现miR-1202是一种灵长类动物特有并富集于人类大脑的miRNA,在抑郁症患者中显著下调;同时验证了其靶基因代谢型谷氨酸受体4(GRM4)在抑郁患者的尸脑和外周血样本中均显著上调,GRM4位于突触前膜和突触后膜,具有调节谷氨酸、多巴胺、γ-氨基丁酸能和血清素等神经递质的功能。
2.2 体液相关的miRNAs研究 多项研究显示可在体液如血液和脑脊液中检测到miRNAs[32-35]。基于miRNAs的相对稳定和来源广泛的特点[36],抑郁症患者体液miRNA检测可能成为一种诊断和治疗工具。有研究显示外周血miRNAs的表达变化与神经元组织的变化相关,有报道阿尔茨海默病、亨廷顿舞蹈病、缺血性和创伤性脑损伤患者血液和脑miRNA有显著相关性[37-38]。目前在抑郁障碍方面,有7项研究使用外周血作为样本[31,39-45];1项研究以脑脊液和外周血作为样本[46];1项研究比较了脑脊液与外周血miRNAs的表达谱差异和异质性[47]。在这些研究中主要以重症抑郁(MDD)患者为研究对象,根据基因芯片和生物信息学分析,对其表达谱发生改变的miRNAs及其调控的与神经系统和脑功能相关的关键通路中的靶基因进行验证。
在基于抑郁患者外周血的miRNAs表达研究中,Wang等[44]发现抑郁症患者基线时的血浆miR-144-5p水平显著低于健康对照,且其表达水平与蒙哥马利抑郁评分呈负相关。Li等[41]则发现miR-182和miR-132随着重症抑郁患者血清中BDNF水平的降低而上调,并且自我评估抑郁量表评分与miR-132水平呈正相关,而与血清BDNF水平呈显著负相关。在32例重度抑郁障碍患者及与其相匹配的健康对照比较研究中也发现,抑郁症患者存在外周血白细胞miR-34b-5p、miR-34c-5p表达升高,并与抑郁汉密尔顿评分以及认知功能相关[45]。Belzeaux等[39]提取16例重度抑郁障碍患者和13例健康对照者外周血单核细胞进行基因芯片筛查,随访8周观测药物应答反应,结果显示重症抑郁症患者9个miRNAs表达上调,5个miRNAs表达下调,其中2个miRNAs随访8周后表现出稳定的过表达,并鉴定出可能具有预测治疗应答价值的4种基因(PPT1、TNF、IL1B和HIST1H1E)的组合。Fan等[40]同样使用外周血单核细胞研究报道了5种miRNAs的改变与重度抑郁症相关。
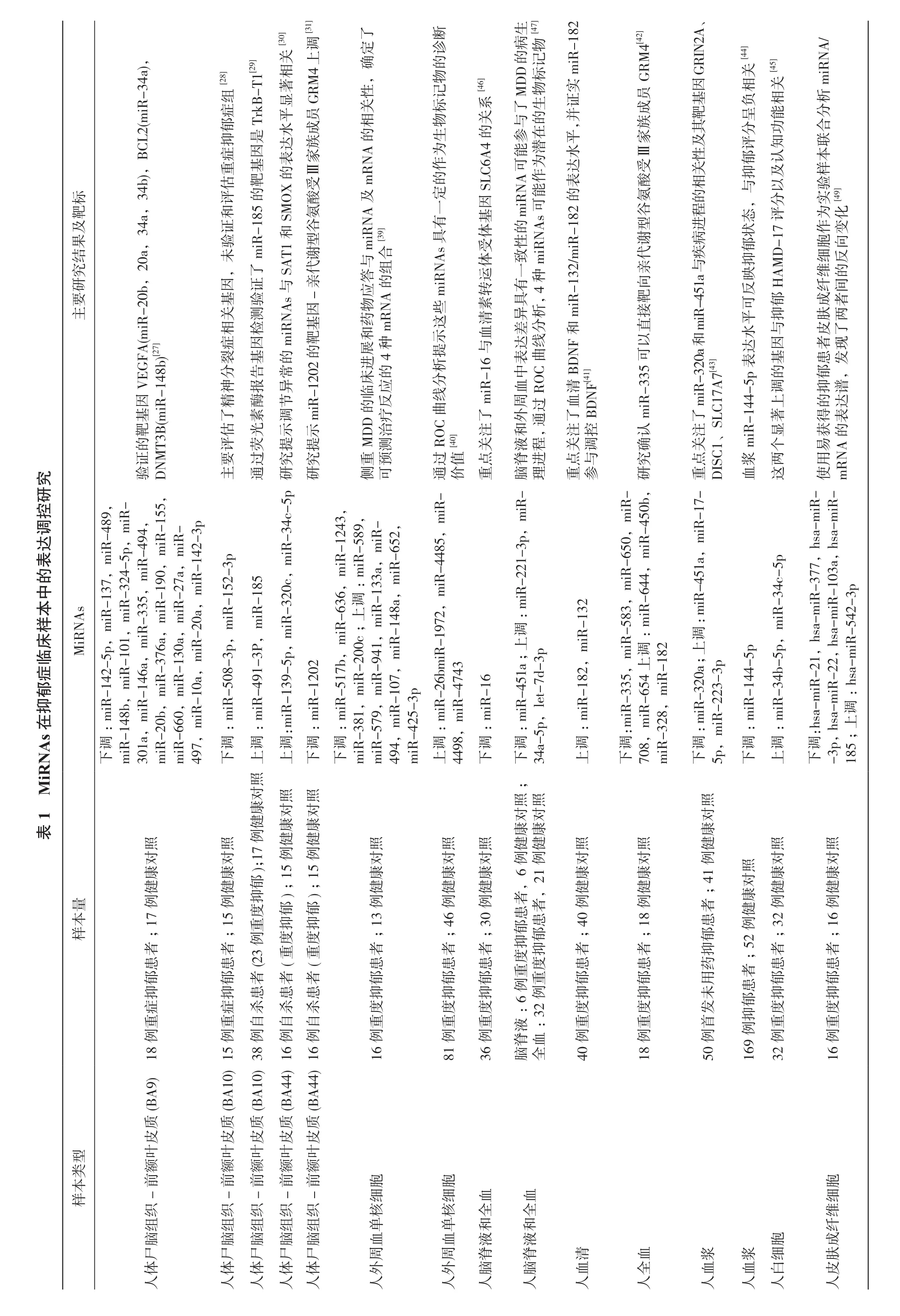
表1 M i R N A s 在抑郁症临床样本中的表达调控研究样本类型样本量M i R N A s主要研究结果及靶标人体尸脑组织-前额叶皮质(B A 9)1 8例重症抑郁患者;1 7例健康对照下m i R-调:m i R -1 4 2-5 p,m i R-i R 1 3 7,m i R-4 8 9,1 4 8 b,m i R -1 0 1 m i R-,m 3 3 5,-3 2 4-5 p,4 9 i R i R 2-m i R-3 0 m i R-1 a,m i R -1 4 6 a,m i R-4,-1 5 5,2 0 6 6 m i R-b,0,m i R-3 7 6 a,m i R i R -1 9 0,m m i R-m i R-1 3 0 a,m-2 7 a,m 1 4 4 9 7,1 0 a,m i R-2 0 a,m i R--3 p验D N 证M T 的V E G F A(m i R-2 0 b,2 0 a,3 4 a,3 4 b),B C L 2(m i R-3 4 a),靶3 B 基(m 因i R -1 4 8 b)[2 7]人体尸脑组织-前额叶皮质(B A 1 0)1 5例重症抑郁患者;1 5例健康对照下调:m i R -5 0 8-3 p,m i R-1 5 2-3 p主要评估了精神分裂症相关基因,未验证和评估重症抑郁症组[2 8]人体尸脑组织-前额叶皮质(B A 1 0)3 8例自杀患者(2 3例重度抑郁);1 7例健康对照上调:m i R -4 9 1-3 P,m i R-1 8 5通过荧光素酶报告基因检测验证了m i R-1 8 5的靶基因是T r k B -T 1[2 9]人体尸脑组织-前额叶皮质(B A 4 4)1 6例自杀患者(重度抑郁);1 5例健康对照上调:m i R -1 3 9-5 p,m i R-3 2 0 c,m i R -3 4 c-5 p研究提示调节异常的m i R N A s 与S A T 1和S M O X 的表达水平显著相关[3 0]人体尸脑组织-前额叶皮质(B A 4 4)1 6例自杀患者(重度抑郁);1 5例健康对照下调:m i R -1 2 0 2研究提示m i R-1 2 0 2的靶基因-亲代谢型谷氨酸受Ⅲ家族成员G R M 4上调[3 1]人外周血单核细胞1 6例重度抑郁患者;1 3例健康对照下m i R-调:m i R -5 1 7 b,2 0 0 c m i R-上6 3 6,:m 3 a,m m i R--5 8 9 1 2 4 3,3 8 1,m i R-;m i R-调i R m i R-5 7 9,m i R-9 4 1,1 3 i R,-4 9 4,m i R-1 0 7,m i R-1 4 8 a,m i R -6 5 2,m i R-4 2 5-3 p侧可重预M D治D疗的反临应床的进展种和物N A 应m i R N A 及m R N A 的相关性,确定了测4药m R答的与组合[3 9]人外周血单核细胞8 1例重度抑郁患者;4 6例健康对照调:m i R -2 6 b m i R-1 9 7 2,m i R -4 4 8 5,m i R-上4 4 9 8,m i R -4 7 4 3通价过值R O C 曲线分析提示这些m i R N A s 具有一定的作为生物标记物的诊断[4 0]人脑脊液和全血3 6例重度抑郁患者;3 0例健康对照下调:m i R -1 6重点关注了m i R-1 6与血清素转运体受体基因S L C 6 A 4的关系[4 6]人脑脊液和全血脑全脊血液:3:6例重抑患,6 1例例健健康康对对照照;2例重度度抑郁郁患者者,2下3 4 a-调:m i R -4 5 1 a;3 p 上调:m i R -2 2 1-3 p,m i R-5 p,l e t-7 d-脑理脊进液程和,中C 表曲达线差分异析具,4有性的A s m i R N能A作可为能潜参在与的了生M D生外通周过血R O一种致m i R N可物D标的记病物[4 7]人血清4 0例重度抑郁患者;4 0例健康对照上调:m i R -1 8 2,m i R -1 3 2重参点与关调注控了B D 血B D N F 和m i R-1 3 2/m i R-1 8 2的表达水平,并证实m i R-1 8 2清N F[4 1]人全血1 8例重度抑郁患者;1 8例健康对照下7 0 8,调:m m i R-i R -3 3 5,m i R -5 8 3,m i R -6 5 0,m 5 0 b,i R -6 5 4上1 8 2调:m i R -6 4 4,m i R -4 m i R-3 2 8,m i R-研究确认m i R-3 3 5可以直接靶向亲代谢型谷氨酸受Ⅲ家族成员G R M 4[4 2]人血浆5 0例首发未用药抑郁患者;4 1例健康对照下5 p,m 调:m i R i R -3 2 0 a;上调:m i R -4 5 1 a,m i R-1 7--2 2 3-3 p重D I S C 点关1、注m i R-3 2 0 a 和m i R-4 5 1 a 与疾病进程的相关性及其靶基因G R I N 2 A 、了S L C 1 7 A 7[4 3]人血浆1 6 9例抑郁患者;5 2例健康对照下调:m i R -1 4 4-5 p血浆m i R-1 4 4-5 p 表达水平可反映抑郁状态,与抑郁评分呈负相关[4 4]人白细胞3 2例重度抑郁患者;3 2例健康对照上调:m i R -3 4 b-5 p,m i R-3 4 c-5 p这两个显著上调的基因与抑郁H A M D -1 7评分以及认知功能相关[4 5]人皮肤成纤维细胞1 6例重度抑郁患者;1 6例健康对照下-3 p,调:h h s a-调s a-m m i R-i R -2 1,h s a-m i R--1 0 3 3 7 7,a,h s a-m i R-2 2,h s a-m i R h s a-m i R-1 8 5;上:h s a-m i R -5 4 2-3 p使m R 用N A 易实验样本联合分析m i R N A/获的得表的达抑谱郁,患发者现皮了肤两成者纤间维的细反胞向作变为化[4 9]

表2 MiRNAs在抗抑郁药物治疗临床样本中的表达调控研究抗抑郁药物 样本类型MiRnAs主要研究结果及靶标氟西汀 重度抑郁患者脑脊液(n=9) 给药上调:miR-16 MiR-16是SSRIs治疗和海马神经生成的调控分子,氟西汀给药后抑郁患者的脑脊液样本及小鼠海马脑组织中的miR-16表达水平增高 [17]艾司西酞普兰 重度抑郁患者全血(n=10)给药上调: miR-130b,miR-505, miR-29-b-2, miR-26a/b, miR-22,miR-664, miR-494,let7d/e/f/g, miR-629, miR-106b, miR-103, miR-191, miR-128,miR-502-3p, miR-374b, miR-132, miR-30d, miR-500,miR-589,miR-183, miR-574-3p,miR-140-3p, miR-335, miR-361-5p给药下调: miR-34c-5p, miR-770-5p在西酞普兰8周治疗后,28个 miRnAs上调,2个miRnAs下调,miRnA靶基因预测和功能注释分析显示在几条重要的与神经脑功能相关的通路上明显富集 [50]西酞普兰 重度抑郁患者全血(n=32) 给药上调:miR-1202抑郁患者中miR-1202表达水平降低,而在8周西酞普兰治疗后其表达水平明显上升;miR-1202调节亲代谢型谷氨酸受体4表达水平并且能够预测基线水平的药物治疗反应 [30-31]西酞普兰 重度抑郁患者全血(n=18) 给药上调:miR-335重症抑郁患者miR-335表达水平下调,在西酞普兰治疗后上调,而GRM4表达水平下降 [42]文拉法新(n=7)帕罗西汀(n=7)氟西汀(n=3)艾司西肽普兰(n=11)度洛西汀(n=1)舍曲林(n=3)米氮平(n=2)重度抑郁患者外周血单核细胞(n=32) 给药下调:miR-124 MiR-124在MDD患者中表达水平上调, 在抗抑郁药物治疗8周miR-124表达水平明显下调,尤其是在药物应答组 [52]文拉法新(n=28)艾司西肽普兰(n=27)度洛西汀(n=124)重度抑郁患者全血(n=179) 给药上调:miR-1202,miR-16,miR-135 MiR-1202在3种药物的队列中均出现应答组的基线水平降低,而治疗后表达水平升高,ROC曲线分析,对药物疗效具有较好的基线预测水平;miR-16和miR-135a的结果显示组间变异性 [53]文拉法新舍曲林米氮平抑郁患者外周血单核细胞(n=20)给药上调:miRNA-1972,miRNA-4485,miRNA-4498,miRNA-4743给药后上调的miRNAs与抑郁患者阻滞症状的改善程度呈正相关[51]
Song等[46]在对36例重症抑郁和30例健康对照者的脑脊液检测发现,重度抑郁患者脑脊液中miR-16表达水平显著低于对照组,并与汉密尔顿评分呈负相关,与脑脊液5-羟色胺水平呈正相关。Wan等[47]则分析了miRNAs在重度抑郁患者脑脊液和血清中表达的差异,并进一步选取了两种样本中具有表达差异一致性的miRNAs(3种表达上调miR-221-3p、miR-34a-5p、let-7d-3p和1种表达下调miR-451a),在另外32例抑郁患者和对照血清中进行了诊断试验验证,ROC曲线结果显示其有较高灵敏度和特异度,具有作为抑郁疾病生物标记物的潜在价值。
2.3 皮肤成纤维细胞相关的miRNAs研究 在研究情感障碍患者信号转导和分子水平的发病机制过程中发现,皮肤成纤维细胞是一种简单、与发病相关、易获得的且尚未被充分利用的模型[48]。Garbett等[49]使用微阵列技术检测16例重症抑郁患者和匹配健康对照者的皮肤成纤维细胞样本的mRNA和miRNA表达水平,初步确定MDD患者成纤维细胞的miRNA和mRNA表达谱改变可能与大脑病理变化相关,其中在MDD中显著减少的miR-122可能成为潜在的诊断生物标记物。
3 抗抑郁药物治疗对miRNAs表达调控的影响
目前涉及miRNA在抗抑郁药物治疗临床样本中表达调控的研究较少,且均集中在一线治疗药物5-羟色胺再摄取抑制剂(SSRIs)和5-羟色胺/去甲肾上腺素再摄取抑制剂(SNRIs)。其中氟西汀1项,西酞普兰2项,艾司西酞普兰1项,度洛西汀1项,联合治疗2项(含文拉法新等选择性NA再摄取抑制剂),这些药物可能通过改变miRNAs的表达来发挥治疗作用(表2)。
Launay等[17]鉴定出miR-16是5-羟色胺再摄取抑制剂在中缝核5-羟色胺能和蓝斑核去甲肾上腺素能中发挥抗抑郁作用的重要效应分子,作用于中缝核5-羟色胺能神经元的氟西汀可降低海马中miR-16的表达水平,从而增加SERT和Bcl-2蛋白的水平。氟西汀给药后抑郁患者的脑脊液样本miR-16表达水平增高,其协同作用的信号分子BDNF、Wnt2、15d-PGJ2亦均增高。另一种经典抗抑郁药物西酞普兰和艾司西肽普兰(外消旋西酞普兰的左旋对映体)也显著影响重度抑郁患者外周血miRNAs的表达。Bocchio-Chiavetto报道[50]抑郁患者应用艾司西酞普兰治疗8周后,28个miRNAs上调,2个miRNAs显著下调,其中13个miRNAs(let-7d、let-7e、miR-22、miR-26a、miR-26b、miR-34c-5p、miR-103、miR-128、miR-132、miR-183、miR-192、miR-335、miR-494)曾经被报道在大脑神经可塑性、应激反应以及其他神经疾病发生中具有重要作用,而这些miRNA的靶标包含神经营养因子BDNF、糖皮质激素受体NR3C1、一氧化氮合成酶NOS1、生长因子(IGF1、FGF1、FGFR1、VEGFa和GDNF)、钙离子通道蛋白(CACN41C、CACNB4、SLC6A12和SLC8A3)以及神经递质受体(GABRA4和5-HT4)等。Li等[42]选取18例重症抑郁患者和匹配对照者的全血样本,结果发现miR-335在MDD中下调,在西酞普兰治疗7 d后上调;与健康对照相比MDD患者中GRM4表达增加,西酞普兰治疗后GRM4的表达下调至对照水平,进一步从临床水平验证了GRM4是miR-335的靶基因。
张巧丽等[51]选取20例抑郁患者和匹配健康对照者,发现抗抑郁药(文拉法新、舍曲林、米氮平)治疗6周后单核细胞中miR-1972、miR-4485、miR-4498、miR-4743与阻滞因子的改善程度呈正相关,miR-26b与日夜变化因子的改善程度呈负相关。He等[52]选取32例MDD和30例匹配健康对照的外周血单核细胞为研究样本,结果显示MDD患者PBMC中miR-124的表达水平显著高于健康对照者,并且ROC分析的miR-124曲线下面积为0.762,灵敏度为83.33%,特异度为66.67%,而在抗抑郁药物治疗8周后,miR-124的表达水平显著下调。Fiori等[53]对179例抑郁患者按抗抑郁药物种类分为队列1(艾司西肽普兰与文拉法新)和队列2(度洛西汀)进行研究,检测了给药8周后抑郁患者外周血中miR-1202、miR-135a和miR-16的表达水平,发现基线miR-1202水平对治疗后抗抑郁药应答具有预测价值(AUC=0.812),进一步验证了miR-1202与谷氨酸系统的调控关系,而miR-135a和miR-16则存在组间差异表达的不稳定性。
Lopez等[31]基于汉密尔顿抑郁量表评分的变化,将经过西肽普兰治疗8周后的抑郁患者分为缓解组(REM)和无应答组(NRES)。有趣的是,药物治疗前REM组的miR-1202表达下调,与健康对照和NRES均存在差异,而NRES与健康对照之间没有差异,REM组在治疗8周后miR-1202水平增加,而NRES没有产生差异性变化。这些发现证实了MDD患者外周miR-1202表达与西酞普兰治疗预测之间的关系,同时miRNA-1202也是第一个被证实在抑郁患者尸脑组织和外周血中均表达异常的miRNA。上述研究表明miR-1202有潜力作为预测抗抑郁药物的反应的标记物,但引起这种疗效差异的特异性miRNAs的深层机制还不明确。
4 结语
抑郁症是一种全身性疾病,发病机制复杂,相对于目前以单胺递质为靶点的一线治疗药物来说,研发同时对多个发病机制具有治疗作用的药物是一项崭新的课题。MiRNA参与抑郁症的多个经典发病机制,通过靶向多个节点基因,从而调控关键蛋白的表达,这意味着miRNA本身具有作为抑郁症治疗靶标和前瞻性生物标记物的巨大潜能。虽然如此,有关miRNA直接介导抑郁或者抗抑郁的机制研究还比较少。总体来说,miRNA在抑郁症发病机制和治疗方面的研究仍处于初步阶段,需要更深一步的临床和临床前研究。
猜你喜欢
现代临床医学(2022年2期)2022-04-19
中老年保健(2021年5期)2021-12-02
天然产物研究与开发(2019年1期)2019-03-01
中成药(2018年4期)2018-04-26
中成药(2018年1期)2018-02-02
中国卫生标准管理(2015年5期)2016-01-14
中国卫生标准管理(2015年4期)2016-01-14
中国卫生标准管理(2015年3期)2016-01-14
中国科技信息(2015年2期)2015-11-16
中国科技信息(2015年6期)2015-11-10
