
长期监测与野外控制试验揭示温带典型森林结构、功能及演变规律
2019-09-02 09:36郑兴波于大炮王安志戴冠华
中国科学院院刊 2019年8期
郑兴波 于大炮 王安志 戴冠华
1 中国科学院沈阳应用生态研究所 沈阳 110016 2 中国科学院长白山森林生态系统定位研究站 沈阳 110016
长白山是我国东北地区的最高峰,位于吉林省延边朝鲜族自治州,地跨安图县、抚松县和长白县,东南与朝鲜毗邻。长白山拥有欧亚大陆从中温带到寒带主要植被类型,是研究森林生态系统对气候变化响应的天然实验室。其低海拔分布的原始阔叶红松林是我国目前温带面积最大、保护最为完整的森林生态系统,一直以来作为东北温带森林资源保护、恢复和可持续经营的重要参照系,备受国内外学者关注。
长白山是欧亚大陆北半部最具代表性的典型自然综合体,山地森林生态系统保存着最完好和最丰富的物种基因库,也是世界上同纬度地区保存最完好、面积最大的原始森林分布区。巨大的海拔差异,导致水热条件的明显不同,从而形成了长白山自上而下明显的环境梯度,造就了长白山类型多样的自然植被,构成了独特的自然景观格局。为保护典型的火山地貌景观和复杂的原始森林生态系统,保存野生动植物种质资源,保护、拯救和扩繁珍稀濒危生物物种,以及保持生态系统的自然演替过程,1960 年建立综合性的自然保护区(127°42'55″ E—128°16'48″ E,41°41'49″ N—42°51'18″ N),总面积达 19.6 万公顷。1980 年 1月,经联合国教科文组织批准,该保护区加入国际生物圈保护区网,列为世界自然保留地之一,1986 年晋升为国家级自然保护区。2003 年,长白山自然保护区被国际人与生物圈、人与地理圈、山地研究发起组织等 10 个国际组织评为全球 28 个环境监测点之一。
1979 年,中国科学院林业土壤研究所(现“中国科学院沈阳应用生态研究所”)受中国科学院委托,由著名生态学家王战先生牵头创建了中国科学院长白山森林生态系统定位研究站(以下简称“长白山站”),从此开始了以原始森林生态系统结构、功能及其演变过程为核心的长期综合监测和试验研究工作,为阐明人类活动和环境变化对森林生态系统的影响以及森林生态系统对环境的反馈作用,寻求森林资源持续发展和森林优化管理的有效途径提供理论依据。
1 温带原始阔叶红松林碳汇及其形成机制
阔叶红松林是我国东北地区东部山地主要森林植被类型,分布广、面积大,遍布于长白山、完达山和小兴安岭山地。属于温带典型的针阔混交林植被,具有涵养水源、保持水土、调节气候、维持生态平衡等多种生态功能。
20 世纪 80 年代,全球变化生态学在国内外受到了前所未有的关注,成为生态学研究的前沿和热点。而森林生态系统在调节全球碳平衡、减缓大气中 CO2浓度上升以及维护全球气候等方面发挥着不可替代的作用,因此研究森林生态系统碳循环机制及其对全球变化的响应成为森林生态学研究的重要工作内容[1]。其中,森林的源/汇功能一直是研究的热点和关键科学问题,备受国内外学者关注[2-4]。
在全球变化背景下,阔叶红松林作为温带典型的地带性顶极森林植被类型,其对全球变化的响应及碳源/汇功能成为长白山站科研人员迫切需要回答的科学问题。
1.1 原始老龄阔叶红松林仍是碳汇
依据连续 11 年 CO2通量观测数据(图 1),针对传统涡度相关方法忽略湍流方程组二阶闭合项的弊端,提出了压力脉动是造成森林生态系统长期碳收支的重要偏差源,并给出了静压脉动通量修正方法。在考虑大气静压脉动的影响后,碳通量的计算精度可提高 15%。得出长白山原始老龄阔叶红松林生态系统仍然是大气的持续碳汇,具有持续固碳能力,其碳汇强度为 170—290 g C m-2yr-1[5-7]。提出固碳潜力参照原生稳定群落固碳假设,获“973”项目“天然森林和草地土壤固碳功能与固碳潜力研究”支持。根据这些科学数据推算,我国温带森林固碳潜力可抵扣全国 1982—2003 年均碳排放总量的 9%—21%。上述研究汇总的成果“长白山老龄阔叶红松林碳汇功能研究”获吉林省自然科学成果奖。
依据长白山站在 1981 年设置的 3 个森林固定监测样地30年的连续监测数据(图 2),得出阔叶红松林生态系统碳汇为 278 g C m-2yr-1,其中活立木和土壤分别为 131 g C m-2yr-1和 150 g C m-2yr-1,固定样地的监测结果与通量计算结果相近。云冷杉林为 96 g C m-2yr-1,其中活立木为 68 g C m-2yr-1,土壤为 70 g C m-2yr-1,而粗木枝残体达到 95 g C m-2yr-1;高山岳桦林为 183 g C m-2yr-1,其中活立木为 124 g C m-2yr-1,土壤为 47 g C m-2yr-1[8]。
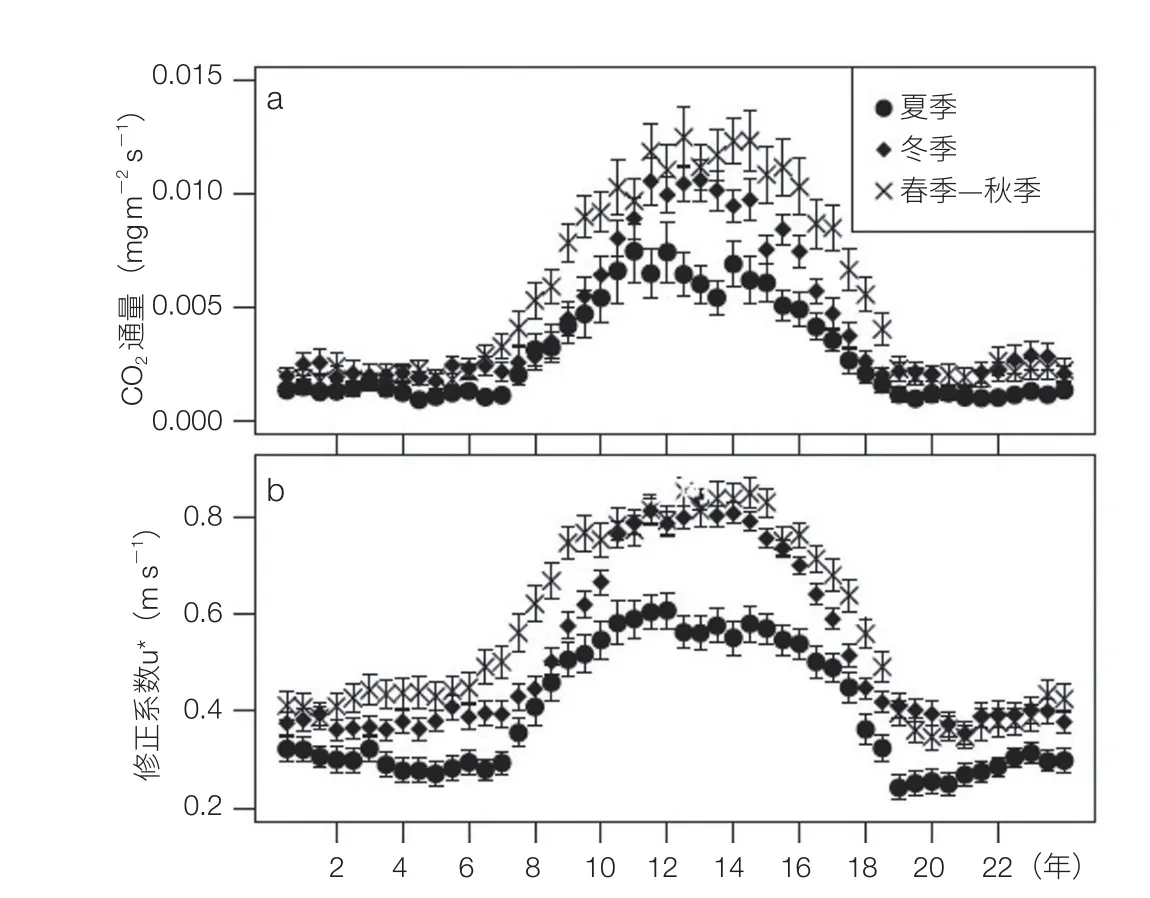
图1 CO2通量不同季节变化图

图2 不同森林类型有机碳密度
1.2 阔叶红松林生态系统生产力形成机制
森林作为陆地生物圈的主体,不仅蕴藏着丰富的生物多样性,而且具有较高的生产力。森林生产力是森林生态系统维持与发展的根本动力,探讨维持森林生产力的内在生物学要素和外在生态条件,是当今生态学和林学研究的重要内容之一。
1.2.1 大树对森林碳汇的贡献大
森林净初级生产力是森林群落在自然环境下的生产能力,也是反映森林固碳能力的重要指标。对森林植被净初级生产力的研究一直是生态学研究的一个重要方面,同时也是全球变化研究的焦点之一[9,10]。尽管我们在不同尺度上对森林生产力的理解有了很大提高[11],但在个体水平上还没有一致的结论。
目前许多研究报道树木生长速度随径级增加而下降,但这些研究通常用植物叶片光合速率来代替个体生物量的累积速度。然而,代谢生态学理论预测植物生物量的增加速率应随个体大小的增加而增加。为准确评估森林树木固碳速率随径级的变化规律,长白山站通过与美国地质调查局等全球 30 个研究机构的 38 位科学家合作,基于全球固定森林样地 403 个树种、672 127 株大径级树木的多次调查数据开展了相关研究。研究发现:全球森林中胸径大于 100 cm 的树木个体年均固碳量约 103 kg,约是胸径为 50 cm 树木个体的 3 倍,96.8% 的树种都在持续固碳,且固碳速率随径级(树龄)的增加而增加[12]。
1.2.2 植被数量是影响阔叶红松林生产力第一因素
基于长白山温带森林样地 10 年的长期监测数据,使用基于机器学习和分类回归树的推动回归树模型(BRT)分析回答了“不同因子对生产力形成的贡献”这一科学问题,量化了生物多样性(包括分类、谱系和功能多样性)、林分结构特征(包括基于胸径的香农-威纳指数和变异系数)、功能性状组成以及初始生物量和土壤养分对生产力的相对贡献。结果发现:温带典型森林生产力主要受初始生物量的影响。植被数量对温带天然林生产力的影响大于植物质量和土壤养分,性状组成和土壤养分对生产力的相对贡献随着森林群落演替的升级而降低,生物多样性和植被数量的相对贡献则随之升高(图 3)[13]。
1.2.3 水力结构研究为阐释森林生产力提供新视角
在温带森林生态系统中,树木越冬过程中频繁发生的冻融交替,可诱导木质部产生气穴化栓塞。气穴化栓塞对树木的水分传输功能造成较严重的损伤(图 4):木质部管道(导管或管胞)中的水柱在结冰过程中,溶解其中的空气离析出来形成大小不一的气泡;冰融化后水柱通常承受较大张力(负压),这时较大的气泡即可迅速扩张导致木质部管道产生气栓,使得其水力结构相关的适应性特征发生了较大改变,威胁木质部正常的水分传输功能。越冬过程中发生的水分传输功能损伤若不能及时修复,将影响树木在生长季的蒸腾用水和光合能力(潜在的生产力),严重时可造成树木衰退死亡,森林生产力降低、建群/优势树种天然更新障碍等问题。
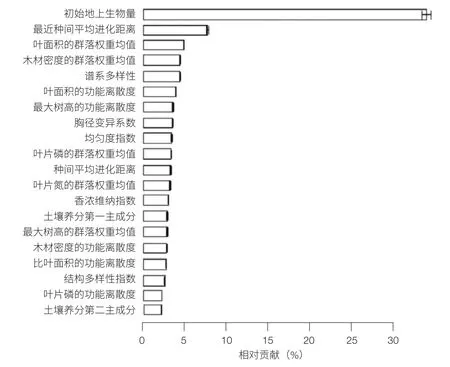
图 3 不同因子对生产力的相对贡献

图 4 长白山针阔混交林常见树种冬季枝条木质部导水率丧失的百分比
(1)植物水力学功能差异与树木生产力密切相关。温带树种除了进化出较强的木质部冻融栓塞抵抗力以外,一些树种(如桦树、槭树)还可在生长季来临前通过产生正的压力(根压或茎压),将木质部导管内的气栓排出维管系统或者溶解到树液中,从而修复冬季栓塞化导管的水分传输功能。正压机制在一些类群中进化出来以后,很可能使得其水力结构相关的适应性特征发生了较大改变,进而对温带树种的环境适应性和生产力产生重要影响。基于此,选取了长白山阔叶红松林中能够产生根压和茎压、只产生根压和两种压力均不能产生的 3 个功能类群的典型树种,开展了树木水力结构对比研究。结果表明,不同功能类群间在木质部导水率、对冻融造成的木质部功能损伤的抵抗力、对干旱诱导栓塞的抵抗力 3 个重要水力学特征上存在显著差异,而该水力学功能的差异是由于不同功能类群间木质部组织水平(光学解剖)和纹孔水平(扫描电镜)的结构特征存在显著分化(图 5)。该研究首次较系统地阐述了纬度较高环境中木质部水分传输效率、对冻融造成的木质部功能损伤的抵抗力、对干旱诱导栓塞的抵抗力三者间较复杂的权衡关系。
(2)水分传导率是树木生产力的重要影响因素。阔叶红松林中“东北三大硬阔”(胡桃楸、水曲柳和黄菠萝)都为复叶树种,而复叶特征长期以来被认为与较高的光合生产力有关。长白山站以长白山地区包括“东北三大硬阔”在内的树种为材料,研究了其木质部水力结构和光合碳同化相关的重要功能性状。结果表明,水分传导率是决定其光合能力(潜在的生产力)的重要因素,而且随着复叶面积的增大,茎水力导度和整枝水力导度都呈显著增加趋势,同时水力导度的增加也使得光合同化速率增大,即光合同化速率也与复叶面积呈显著的正相关关系(图 6)。复叶特征使得树木的叶片面积可以远远大于一般单叶树种,而大的叶片有利于树木构建更为有效的水力传输系统,从而支撑其较高的光合气体交换和较高的生产力。但是在冻融交替等胁迫因素存在的情况下,生产力和抗逆性间存在显著权衡关系,具有较大叶片、较高水力导度、较高光合速率的复叶树种的潜在生产力需要在条件比较优越的环境中才能得以较高的生产力[14]。
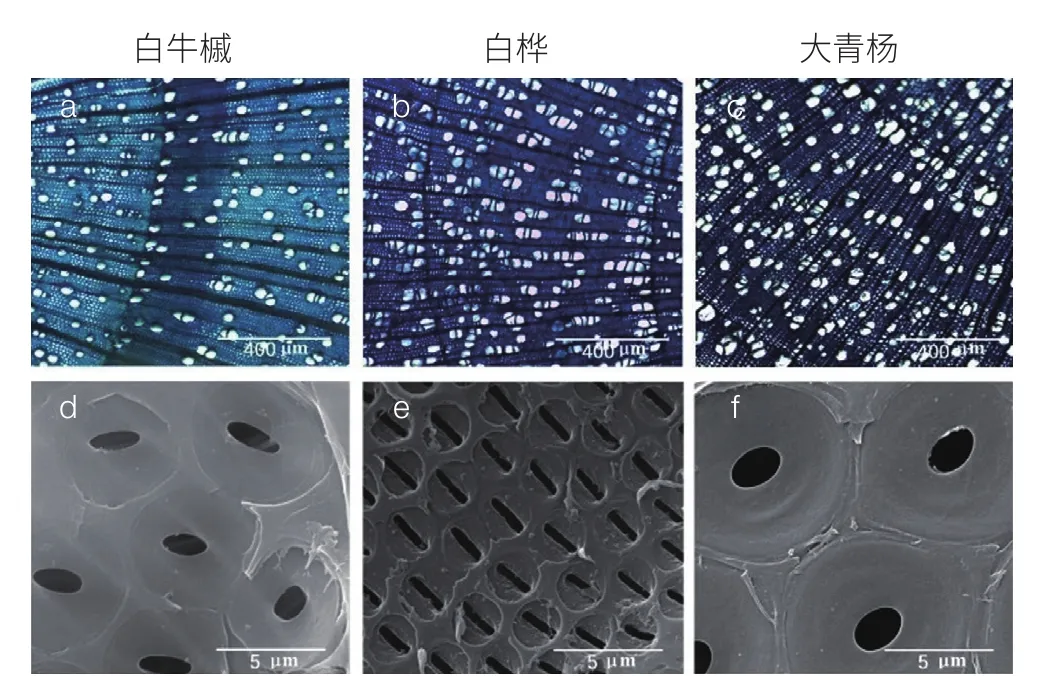
图 5 三个功能类群的代表树种的组织水平和纹孔水平木质部的解剖图
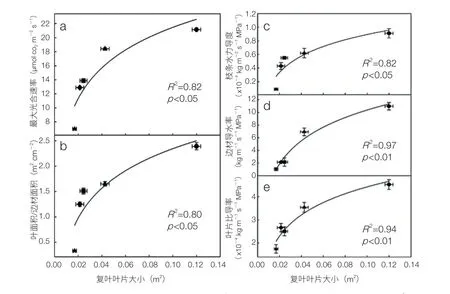
图6 长白山阔叶红松林5种复叶树种整枝导水率、最大净光合速率、枝条边材比导率、叶比导率与叶片面积间的正相关关系
2 阔叶红松林碳循环过程对 CO2 升高的响应
利用开顶箱熏蒸系统开展 CO2浓度升高对红松、长白松和蒙古栎的生长、生理生态过程影响的科学实验。该系统的独特之处在于自籽实萌芽起,在植物的生长季节坚持昼夜高浓度(500 mg/L CO2)熏蒸。该系统在承担中国科学院知识创新重大项目(2000—2005 年)和“973”项目(2003—2007 年)期间,完成了红松和蒙古栎幼苗的细根生长、死亡和快速分解等研究内容;同时,着重开展了开顶箱和林地自然条件的幼苗细根周转(包括凋落物)的互置试验。
2.1 树木叶片固碳能力和土壤呼吸通量增加
CO2浓度升高在增加树木叶片固碳能力的同时,也通过树木根系加强了 CO2排放的通量。通过对 CO2升高与对照情况下生长季呼吸通量与叶片固碳的比率比较分析,土壤呼吸量增加 21%—24.5%,而叶片固碳能力增加 28.8%,大气 CO2升高对叶片固碳能力的贡献大于对照实验[15]。
2.2 细根周转过程减缓,土壤碳素滞留时间延长
600 天的分解试验结果显示,分解初始阶段细根表现出氮(N)的固持,然后表现出矿化现象。对照箱中的细根分解表现出更高的 N 固持率,N 净固持期限也延长,超出 300天,而高浓度 CO2箱中的细根分解 N 的净固持时限较短;高浓度 CO2箱细根分解过程中 N 释放率显著高于对照箱,表明高浓度 CO2使地下 N 周转加快;高浓度 CO2箱细根木质素残留量显著低于对照箱,表明高 CO2使细根碳循环减慢,有助于碳在土壤中长期滞留,增加土壤碳库含量。在整个分解过程中,纤维素含量持续下降,CO2处理间差异显著,高浓度 CO2处理低于对照。这说明高 CO2下易变有机碳周转加快,有助于土壤微生物活性和植物营养元素供给。
CO2处理使土壤脱氢酶活性提高了19%,转化酶活性提高 9.7%,蛋白酶活性降低 1.0%,多酚氧化酶升高 9.9%,木聚糖酶活性提高 7.3%,纤维素活性提高 8.1%。土壤酶活性研究显示,高浓度 CO2提高土壤微生物活性,特别是易变碳分解微生物功能群的活性,而对 N 周转功能微生物影响并不显著,这说明土壤微生物可通过提高自身周转速率,满足植物快速生长的营养需求[16]。
2.3 阔叶红松林碳氮水多因子综合野外模拟实验
2008 年秋季,构建了长白山原始阔叶红松林大规模(占地 5 ha)野外多因子(增氮、控水、增温)模拟实验装置(图 7)。该装置现已成为我国温带森林与全球变化关系研究的综合研究场地,以及承担国家级科研任务、实现广泛国际合作和研究团队公共技术平台。
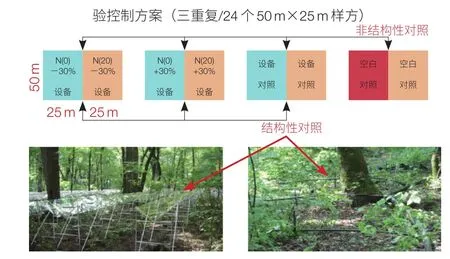
图 7 多因子野外模拟实验装置

图 8 团聚体粒径的全球增温潜势在施氮处理下的差异
在阔叶红松林和天然次生林内设立了控水施 N 试验地,参考国外同类研究及所在地 N 沉降情况,将实验地设置为 24 块 50 m×25 m 样方。施氮量 0 kg N yr-1h-1、50 kg N yr-1h-1;±30% 降水量;地表增温 <2℃。按照增 N×(+N)交叉设计,分别为对照(有无 PVC 板)、对照 +N、减水 30%、减水 30%+N、增水 30%、增水 30%+N,每种设计三重复。
±30% 降水控制,是针对控制穿透雨量进行的。采取铺设透明 PVC 板遮挡样地面积的 30% 实现降水量减少 30%;而被截留的 30% 穿透雨再通过收集系统自动灌注到毗邻的林地,实现林地降水量增加 30%。
2.3.1 N 添加增强了土壤有机碳库稳定性
通过连续 6 年野外多因子模拟实验,结合室内培养,探讨在长期施 N 的情况下土壤有机碳稳定性的变化趋势(图 8)。研究结果表明,与对照和低施 N 量处理(25 kg N ha-1yr-1)相比,长期的高 N(50 kg N ha-1yr-1)施加显著抑制土壤 CO2的排放,高 N 处理土壤 CO2排放速率较对照处理低 30%,土壤有机质的稳定性有所提高。其机制主要体现在两个方面:大团聚体对微团聚体的保护作用增强(图 9);土壤有机质本身的抗分解性提高(同位素 δ13C 作为指示指标)[17]。
2.3.2 氮添加减少了细根的量,增加了周转速率
利用多因子野外模拟实验平台,对长白山原始阔叶红松林细根动态的 6 年调查研究显示,细根生物量、生产量、死亡量和消失量均表现出明显的季节动态(图 10)。月细根量受前月气候条件(如月平均温、月降雨量)的影响,而月细根生产量则与当月的平均温关系密切;细根生产量的季节动态与森林净初级生产力(NPP)的季节动态相似,与凋落物量的季节动态相反;总细根生产量占阔叶红松林 NPP 的 25%,经由细根向土壤输入的碳量是地上凋落物的 1.2 倍。
施氮和控水双因子模拟研究表明,阔叶红松林细根量显著降低 47%,细根周转至少增加 1.4 倍;即使在0—20 cm 土壤层中,细根对氮、水的改变表现出明显不同的响应特征,土壤表层(0—10 cm)氮有效性细根形态结构的主要影响因子,而在土壤亚表层(10—20 cm)水分和养分有效性共同影响着细根的生长动态及其形态结构(图 11)。这些研究成果阐明了温度、降水和氮沉降对阔叶红松林细根动态的影响,量化了细根对土壤碳的贡献,为进一步准确评估全球变化背景下我国温带森林生态系统的固碳速率和潜力提供了数据支持[18,19]。
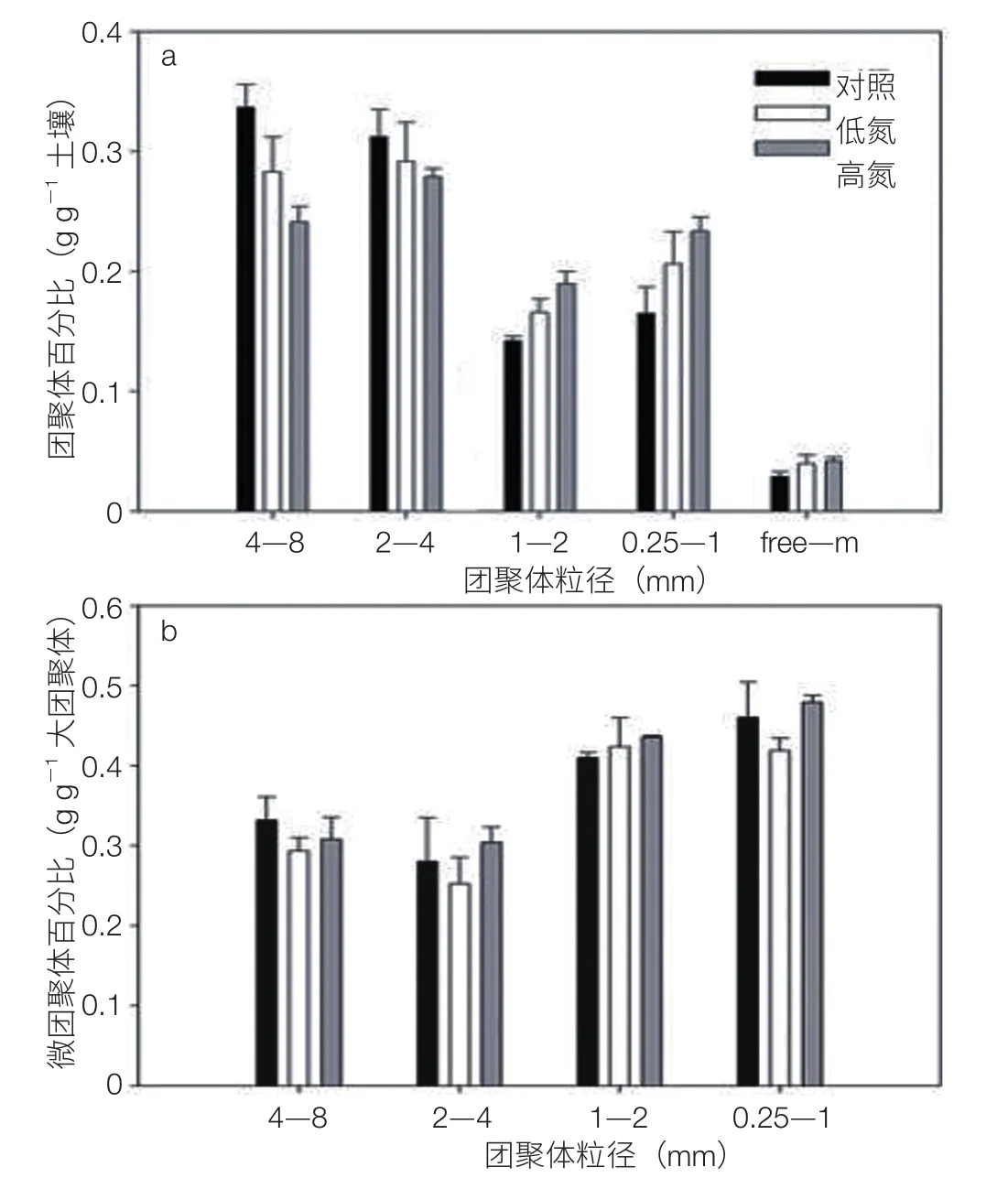
图9 土壤团聚体的分布规律(a)及大团聚体包裹的微团聚体含量变化(b)
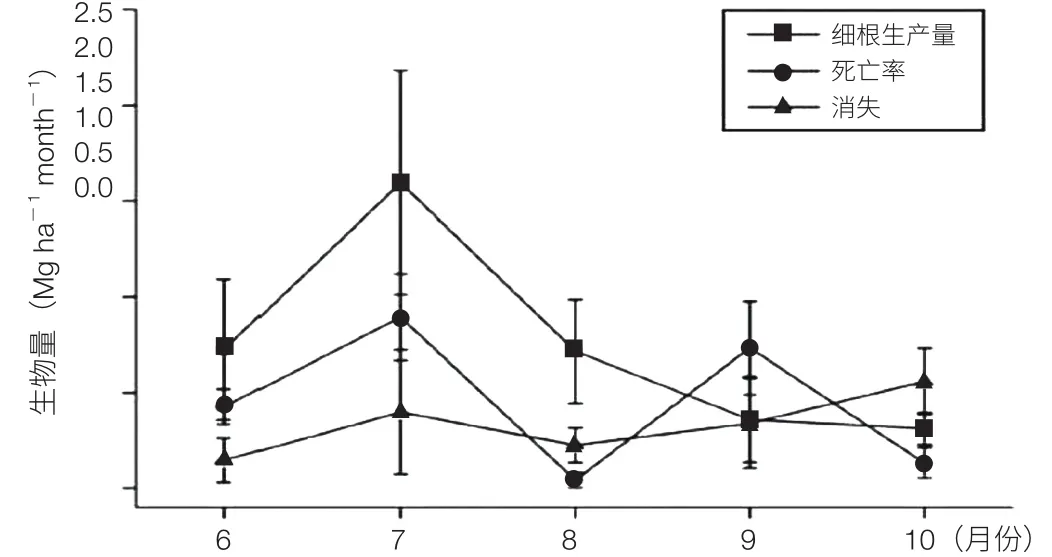
图 10 细根生产量(MP)、死亡率(MM)和消失(MD)的季节动态

图 11 土壤硝态氮和铵态氮(a)以及细根生物量和坏死量(b)
3 阔叶红松林群落结构内在机理及生物多样性维持机制研究
为揭示温带典型森林群落构建、物种多样性共存机制、物种多样性格局等国际前沿和热点科学问题,长白山站以国际温带森林生物多样性网络监测大样地为研究平台,在长白山北坡原始阔叶红松林建立了50 ha 的长期固定监测样地,开展温带典型森林物种多样性维持机制、多样性格局、群落构建等相关研究工作。
3.1 阔叶红松林群落构建过程机理研究
3.1.1 群落构建中随机性与确定性过程贡献具有尺度依赖性
在国家自然科学基金委员会和美国科学基金会双方合作交流项目的资助下,探讨群落构建中确定性过程和随机性过程在不同尺度上的相对贡献。首次在大尺度上运用新的统计推断手段——最大熵原理阐明了群落构建过程的重要性具有尺度依赖性。
结果发现:基于物种性状的环境过滤在环境变异较小的小尺度上几无作用,而在有明显环境梯度的中大尺度上作用显著。基于不同尺度物种库的集团效应在不同尺度都有较高贡献,表明区域生态过程在决定局域群落构建中具有重要作用[20]。
3.1.2 种子结实机制与时空动态对阔叶红松林群落构建的影响
以长白山阔叶红松林生物多样性长期监测样地内,连续 8 年的野外种子收集数据为研究对象,检测了群落内 20 个木本植物物种 8 年间的种子生产的变异性和同步性(图 12),并且验证了传粉效率假说以及气象因子对种子生产的直接作用,即长白山温带阔叶红松林群落水平的种子生产动态同时决定于进化中的选择压力(传粉效率假说)和气象因子的直接作用(图 13)。而捕食者饱和假说在这 8 年的研究中仅仅表现出微弱的作用[21]。
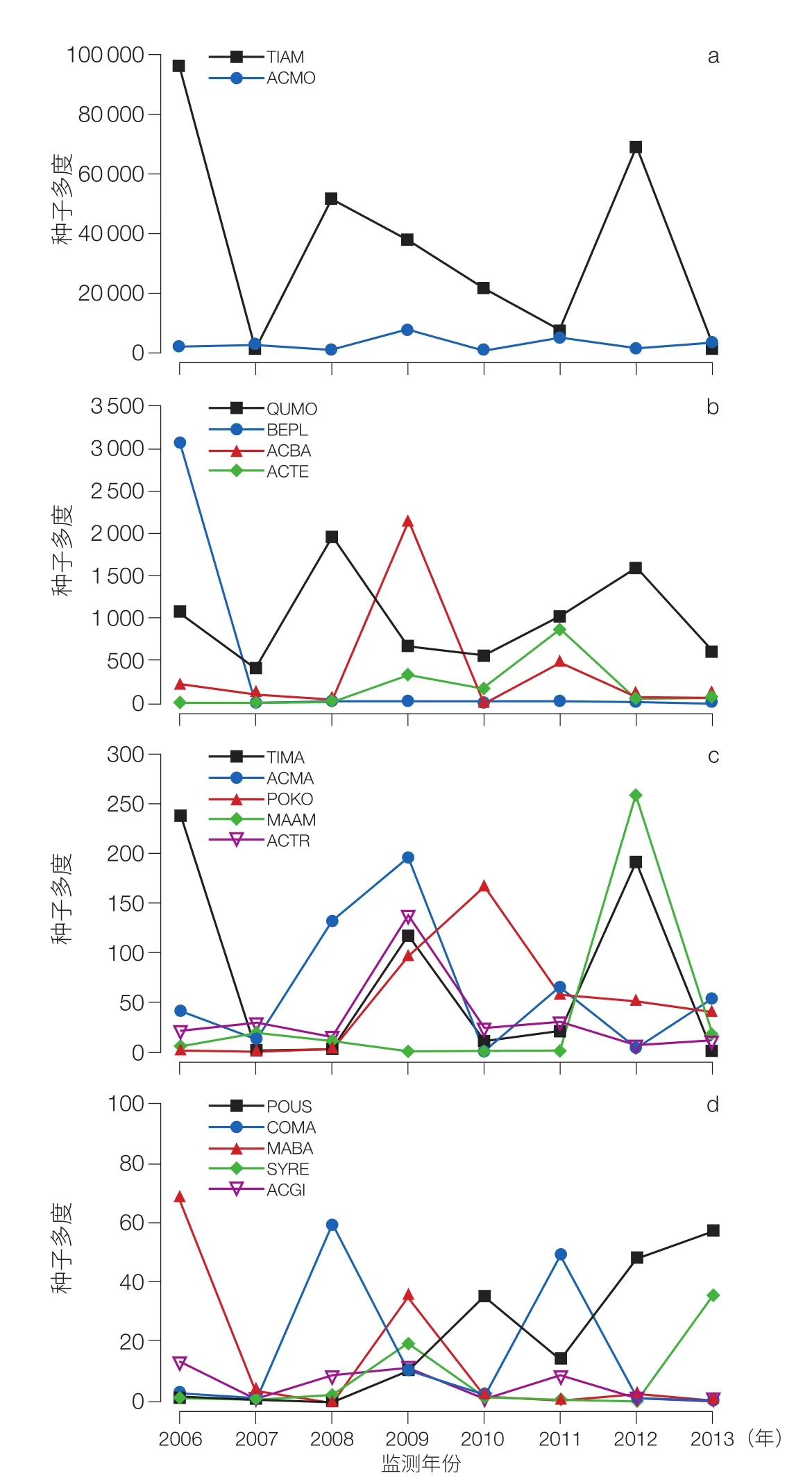
图 12 种子生产的年际变异
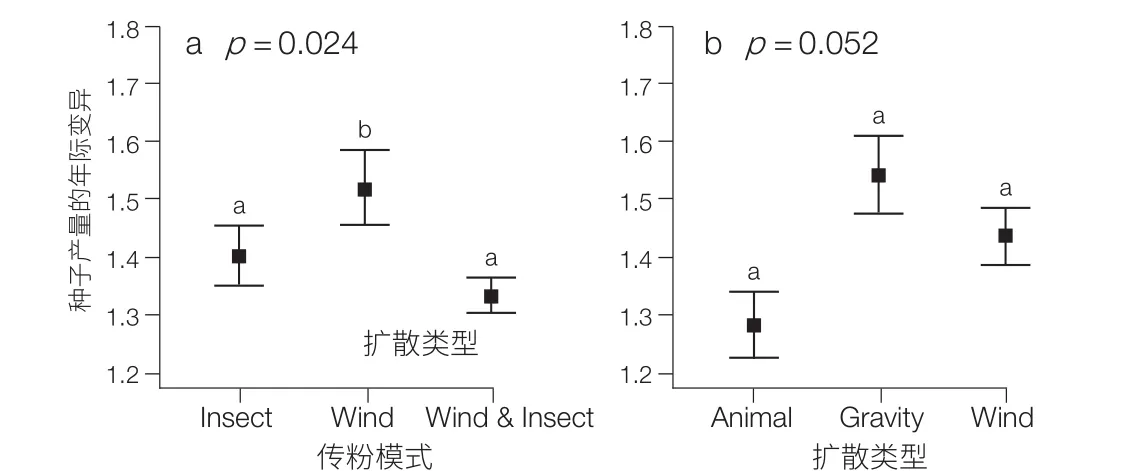
图 13 不同传粉模式(a)和扩散方式(b)的物种组间的平均年际种子产量变异
3.2 环境过滤和扩散限制是阔叶红松林 Beta 多样性格局主要驱动力
以长白山生物多样性长期固定监测样地在内的全球温带地区 8 个面积大于 20 ha 森林样地为研究平台,利用不同空间过程模型,分别模拟样地内所有胸径大于 1 cm 木本植物的空间分布格局,探讨种间关系、环境过滤和扩散限制等对温带森林 Beta 多样性格局及其组分(周转和嵌套)的影响及其相对作用。
研究表明:在 8 个温带森林样地中,物种周转格局所占的比例要高于嵌套格局。环境过滤和扩散限制共同影响 Beta 多样性、物种周转和嵌套格局,而种间关系的作用较弱。对于 Beta 多样性和物种周转格局,扩散限制的作用要强于环境过滤;而对于嵌套格局,环境过滤的作用不弱于扩散限制[22]。
同时发现,不同生态学过程的作用具有明显的尺度依赖性:在较大的空间尺度上(150—250 m)生境过滤起关键作用,在中等尺度下(20—150 m)扩散限制的作用最强,而种间关系仅在小空间尺度(小于20 m)上起作用。由此可见,生境过滤和扩散限制共同影响局域尺度上系统发育和功能 Beta 多样性格局,而以往研究发现扩散限制是局域尺度上该森林木本植物物种 Beta 多样性格局的主要驱动力,利用物种的系统发育和功能性状可进一步加深对森林群落结构内在机理的理解。此外,研究发现种间关系对系统发育和功能 Beta 多样性格局的作用较弱,这与其对系统发育和功能 Alpha 多样性格局的作用一致[23]。
3.3 群落木本植物多样性维持机制研究—稀释假说
生物多样性维持机制一直是生态学研究的热点问题之一,但存在很大争议[24-26]。例如,传统的起源于物种较少地区的生态学理论强调种间关系和环境异质性等确定性过程对于维持生物多样性的作用,然而由于其不能很好地解释物种丰富地区的生物多样性分布格局,近年来受到来自强调随机过程等中性理论的挑战。中性理论假设物种等同,即物种之间没有显著的种间关系。然而许多研究表明无论在温带森林还是在热带雨林,种间关系都显著影响植物的生长和存活,这似乎与中性理论的假设不一致。
通过长白山生物多样性长期监测样地结果分析,提出了随机稀释假说:由于各物种间的关系不一致,随着森林内物种数的增加,目标种与其领域内其他物种的关系随之稀释。这就造成了在物种较少的温带森林呈显著种间关系的物种所占的比例,要高于物种丰富的热带森林。
为了验证该假说,在全球范围基于 5 个面积大于 20 ha 森林样地的数据,利用了多变量点格局分析方法,考虑各树种间的亲缘关系和功能相似性,将树种分为促进种、抑制种和中性种,探讨各类型所占比例随物种数的变化趋势。基于随机稀释假说,中性树种所占的比例应随着树种数的增加而增加(图 14)。研究结果与该假说的预测一致,该假说得到了实测数据的支持,并逐渐被国际研究同行认可。
随机稀释假说验证了中性理论关于物种之间没有显著种间关系的假设在热带雨林的合理性,但不适应于温带森林,丰富发展了森林生物多样性维持机制理论[14]。
4 阔叶红松林演替的生态过程研究
长白山原始阔叶红松林作为我国温带地带性顶极植物群落,在诠释我国幼龄林居多为特点的温带森林的固碳潜力评估中作用不可替代,而原生稳定性森林群落在量化森林生态系统固碳潜力中的参照作用尚未引起足够的重视。同时,不同演替阶段地上—地下生物群落的生态关联,对于我们理解生态系统多样性维持机制和结构稳定性具有重要意义,因此也是生态学关注的热点问题之一[27-29]。
图 14 中性物种所占的比例随物种数的变化趋势
4.1 不同演替阶段地上—地下生态关联具有显著差异性
以长白山阔叶红松林演替前期和演替后期代表林型为研究对象,通过采集大量样方的地上植被和土壤微生物群落组成数据,系统研究了地上—地下群落多样性的生态关联,在群落水平上系统解释了地上植被和土壤微生物之间 Alpha 多样性和 Beta 多样性的生态关联。对于我们理解生态系统多样性维持机制和结构稳定性具有重要意义。
结果表明,植被—微生物之间 Alpha 多样性和Beta 多样性关系在森林不同演替阶段明显不同。对于 Alpha 多样性,演替前期无明显相关关系;而演替后期呈明显负相关,这与我们通常认为的地上植被群落多样性高必然对应较高的地下生物多样性的假设相悖。对于 Beta 多样性,演替前期地上—地下无明显生态关联,而演替后期,呈明显正相关关系,这表明在演替后期土壤细菌群落结构受地上植被群落组成的影响[30]。
4.2 不同演替阶段生物多样性格局与生产力特点
以长白山演替前期杨桦次生林和演替后期阔叶红松林样地为研究对象,利用长期连续、固定监测数据分析了地上生物量的现状和动态[31],收集了叶片磷含量、叶片氮含量、叶面积、比叶面积、木材密度和比根长等功能性状,比较各性状的聚散特征[32],计算了物种丰富度、功能多样性和谱系多样性。同时,使用回归树模型(BRT)量化了生物多样性、林分结构特征、功能性状组成以及初始生物量和土壤养分对生产力的相对贡献。
研究发现:生物多样性对群落生产力的促进作用在演替阶段后期更强。研究结果证实,随着群落的演替,温带天然林中不同的多样性指数对生态系统功能的影响亦发生变化。这说明生物多样性与生态系统功能的关系是一个动态的过程,考虑真实生态系统中多因果关系、多时空尺度和生物多样性的多维性将成为正确理解和把握生物多样性与生产力的突破口。生产力在演替前期中受与叶片磷相关的性状组成、初始生物量、土壤有机质和物种均匀度影响较大,在演替后期则主要受初始生物量的影响。土壤养分对演替前期的影响远大于演替后期;性状组成和土壤养分对生产力的相对贡献随着森林群落演替而降低,生物多样性和植被数量的相对贡献则随之升高[33]。
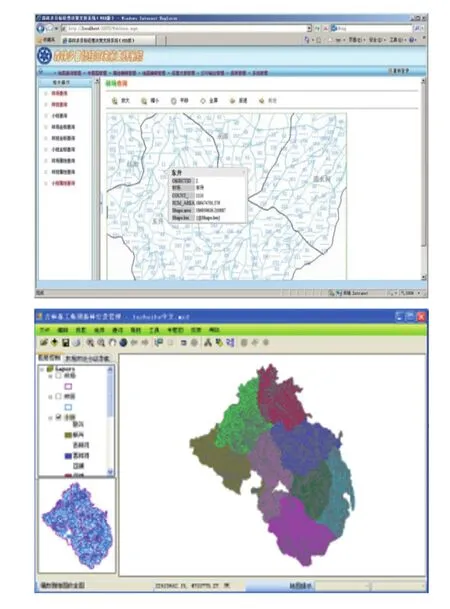
图 15 《森林生态系统管理决策支持系统》
5 服务于地方发展,为社会主义新时期生态文明建设贡献力量
5.1 积极寻求森林资源持续发展和森林优化管理的有效途径
为探求森林资源的可持续经营与管理,长白山站与露水河林业局开展合作研究、试验基于 3S(全球定位系统—GPS、地理信息系统—GIS、遥感—RS)的数字化林业信息技术管理体系。通过该体系的研究和研制,将传统基于林分尺度的森林经营技术转变为基于空间尺度的生态系统管理技术,实现了传统林业到数字林业的转变[34]。通过在不同林业管理部门的试用与完善,研制出了《森林生态系统管理决策支持系统》(图 15),推广示范面积已达 12.1 万公顷。吉林森林工业集团有限责任公司科学技术开发中心评价认为:《森林生态系统管理决策支持系统》实现了森林资源数据的空间数据和属性数据的动态更新,提高了森林经营决策精准化,节省了人力、物力和财力,为东北天然次生林的可持续经营作出了重要贡献,为同行业其他地区森林资源的数字化管理提供了借鉴。
5.2 构建首张高密度红松遗传图谱,填补相关知识空白
科技支撑与服务是野外台站服务地方的重要内容,在与吉林省露水河红松种子园合作中,以露水河红松种子园人工授粉区内 94 个子一代(F1)个体为作图群体,SRAP(相关序列扩增多态性标记)、SSR(简单重复序列)和 ISSR(内部简单重复序列) 3 种方法结合,绘制首张高密度红松遗传图谱[35,36]。
构建了红松交配系统,通过分子生物学实验分析,发现现有的种子园尚未受到周边母树林的污染,为支持世代种子园建设提供科学支持。
5.3 植物多样性调查—新物种的发现
在科技部支持下,由长白山站牵头,东北林业大学和北华大学等单位参加的“东北森林植物种质资源专项调查”于 2008 年启动,历时 5 年,2014 年 4 月以“优秀”等级通过验收。
项目取得的主要成果包括:发现植物新种 2 个,新变种 1 个,中国新记录种 1 个(图 16);获得维管束植物 136 科 645 属 1 940 种,占东北林区野生维管束植物近 150 年记录总数的 83.44%,编绘了大兴安岭、小兴安岭、长白山等重点区域和东北林区的森林植物原色图谱,并编绘了东北地区 2 800 种植物的县级产地分布图;采集了 2.5 万号(5 万份)植物标本,收集了近 2 万份种质材料;摸清了东北林木良种基地种质资源现状;提出了有关东北森林植物种质资源保护、珍稀濒危物种保护、外来物种入侵防治等系列建议。项目成果被《光明日报》《科技日报》《科学时报》等多家媒体及国家林业局先后报道。
6 结语
40 年来,长白山站以“观测、研究、示范、服务”为工作宗旨,在不同时期服务于国家科技战略需求,长期致力于林学、生态学研究探索,不断完善和提高野外科学实验的基础设施建设,积极践行开放共享的理念,取得了一大批科研成果,成为青年科学家成长的摇篮。
“十三五”期间,长白山站将聚焦全球性热点问题和国家建设生态文明的实际需求,围绕中国生态系统研究网络(CERN)2020 年战略规划、中国科学院沈阳应用生态研究所“一三五”战略定位和特色研究所建设规划,以森林生物多样性维持及生产力形成机制、森林生态系统碳平衡、森林与水关系等为研究重点,强调监测数据的时空过程与拓展、多源要素的融合与联网观测、地面与遥感等有机结合,构建智能型数据采集及管理平台建设;努力促成区域台站联合网络建设,将综合观测能力提升、数据服务平台建设与扩展和国家重大科研任务、人才团队培养等有机地结合,以揭示温带典型森林结构、功能及演变规律的研究核心方向。通过基础科学研究成果产出、有效履行国际合作的综合观测基础建设平台、稳定且具创新的人才队伍和重大科技成果示范,使长白山站发展成为国内一流,具有显著国际影响的开放型科研基地。
猜你喜欢
北京林业大学学报(社会科学版)(2022年4期)2023-01-04
草业学报(2022年9期)2022-09-16
东北林业大学学报(2022年6期)2022-07-25
——低维护的小型观赏草阔叶山麦冬
花卉(2021年21期)2021-11-11
绿色科技(2021年5期)2021-04-09
学生天地(2020年34期)2020-06-09
辽金历史与考古(2019年0期)2020-01-06
国际木业(2018年4期)2018-09-10
山东工业技术(2016年15期)2016-12-01
前卫文学(2016年3期)2016-07-01
