
宁春4号与河东乌麦杂交F2代穗部性状分析及其重要QTL发掘
2019-09-10 07:22王掌军刘妍刘凤楼李清峰张晓岗刘生祥贾彪
南方农业学报 2019年4期
王掌军 刘妍 刘凤楼 李清峰 张晓岗 刘生祥 贾彪

摘要:【目的】分析寧春4号与河东乌麦杂交F2代的穗部性状,并利用SSR分子标记发掘其重要QTL,为宁夏小麦穗部性状改良提供理论参考。【方法】以宁春4号与河东乌麦杂交F2代的331个单株为材料,利用方差分析、相关性分析、聚类分析和单标记回归分析等方法对5个穗部性状及其重要QTL进行定位研究。【结果】5个穗部性状在F2代呈连续正态分布,符合多基因控制的数量性状的遗传特点。F2代出现许多具有超亲性状的单株,穗长、总小穗数、结实小穗数、穗粒数和穗粒重的平均值分别为8.69 cm、18.01个、16.71个、33.07粒和1.19 g,超中亲比例分别达20.24%、44.71%、41.99%、34.14%和33.84%,超高亲比例依次为6.64%、18.73%、17.22%、34.14%和26.88%。5个穗部性状间均呈极显著正相关(P<0.01),表明这些性状对产量的贡献均较大。基于穗部性状测定结果,在遗传距离为5 cM时,可将F2代331个单株分为八大类群,其中,类群Ⅰ平均穗长最长(为9.96 cm)、穗粒数最多(49.25粒)、穗粒重最重(1.61 g),类群Ⅷ平均总小穗数和结实小穗数最多,分别为21.40和19.57个,表明类群Ⅰ和Ⅷ为穗部性状优异类群。利用19个SSR分子标记共发掘出36个与穗部性状相关的QTL,其中,穗长、总小穗数、结实小穗数、穗粒数和穗粒重QTL数量分别有15、6、6、5和4个,分布在2A、4A、5A、1B、2B、3B、5B、7B、5D、6D和7D等11条染色体上,加性效应为-0.72~1.57,表型贡献率为2%~9%,LOD最大值为23.90,其中,4A染色体上检测到穗长、穗粒数和穗粒重QTL,5A染色体上检测到穗长、总小穗数和结实小穗数QTL,1B和5D染色体上均检测到穗长、总小穗数、结实小穗数、穗粒数和穗粒重QTL,2B染色体上检测到穗长、总小穗数、结实小穗数和穗粒重QTL,7B染色体上检测到穗长和总小穗数QTL,表明4A、5A、1B、2B、7B和5D等6条染色体存在QTL富集区。【结论】小麦杂交F2代遗传性状处于高度分离,蕴藏着最大的数量遗传信息,为相关穗部性状分析及QTL发掘提供了可靠的遗传群体,检测到的36个QTL可用于小麦穗部性状的遗传改良。
关键词: 小麦;杂交后代;穗部性状;分子标记;QTL
中图分类号: S512.103.51 文献标志码: A 文章编号:2095-1191(2019)04-0685-10
Abstract:【Objective】Panicle traits of F2 hybrids from Ningchun No.4 and Hedong black wheat were analyzed and their important QTLs were discovered by SSR molecular markers. It would provide reference for panicle traits improvement of Ningxia wheat. 【Method】Three hundred and thirty-one individual plants of F2 hybrids from Ningchun No.4 and Hedong black wheat were as materials. By means of variance analysis, correlation analysis, cluster analysis and single marker regression analysis, five panicle traits and their important QTLs were studied. 【Result】The five panicle traits showed continuous normal distribution in F2 hybrids,that conformed to the genetic characteristics of quantitative traits controlled by polygenes. There appeared many individual plants with superparental traits in F2 hybrids, the average values of panicle length,gross spikelet number,bearing spikelet number,kernels per spike and kernel weight per spike were 8.69 cm,18.01,16.71,33.07 and 1.19 g, respectively. The proportions of over mid-parents were 20.24%,44.71%,41.99%,34.14% and 33.84%, respectively,and their ultra-high parents were 6.64%,18.73%,17.22%,34.14% and 26.88%, respectively. The five panicle traits were in extremely significant positive correlation among each other(P<0.01), which showed that the contributions of these traits to yield were large. Based on the panicle traits,three hundred and thirty-one individual plants were divided into eight clusters at genetic distance being 5 cM. Group Ⅰ had the longest average panicle length(9.96 cm), the largest kernels per spike number(49.25 kernels) and largest kernel weight per spike(1.61 g). Group Ⅷ had the largest average gross spikelet number and bearing spikelet number, which were 21.04 and 19.57. It showed that group Ⅰ and group Ⅷ were groups with excellent panicle traits. Thirty-six panicle related QTL were discovered by nineteen SSR molecular markers. Among them, QTLs of the panicle length,gross spikelet number, bearing spikelet number, kernels per spike number and kernel weight per spike were 15, 6, 6, 5 and 4, respectively. They were located on 11 chromosomes including 2A,4A,5A,1B,2B,3B,5B,7B,5D,6D and 7D. Their additive effects were -0.72 to 1.57,the contribution rates to phenotype were 2% to 9%,maximum LOD score was 23.90. QTLs of the panicle length,kernels per spike number and kernel weight per spike were detected on chromosome 4A. QTLs of the panicle length,gross spikelet number and bearing spikelet number were detected on chromosome 5A. QTLs of the panicle length,gross spikelet number,bearing spikelet number,kernels per spike and kernel weight per spike were all detected on chromosomes 1B and 5D. QTLs of the panicle length,gross spikelet number,bearing spikelet number, kernel weight per spike were detected on chromosome 2B. QTLs of the panicle length and gross spikelet numbers were detected on chromosome 7B. The results showed that QTL enrichment regions existed in chromosomes 4A,5A,1B,2B,7B and 5D. 【Conclusion】The genetic characteristics in F2 hybrids of wheat are highly separated, and contain the maximum amount of genetic information, which provide the reliable genetic population for the panicle traits analysis and QTLs discovery. The detected 36 QTLs may be used for genetic improvement of panicle traits for wheat.
Key words: wheat(Triticum aestivum L.); hybrid progeny; panicle trait; molecular marker; QTL
0 引言
【研究意义】小麦(Triticum aestivum L.)在世界各地被广泛种植,全球35%~40%的人口以其为主粮(何中虎等,2018;李冬兵等,2019)。我国是世界上最大的小麦生产和消费国,小麦为仅次于玉米和水稻的第三大粮食作物,其生产对保障我国粮食安全具有重要意义(刘志勇等,2018;朱昀等,2019)。随着我国人口逐年增加,特别是工业化、城镇化快速发展,并受农业结构调整、生态退耕和非农建设占用土地等因素影响,全国耕地面积只减不增,提高小麦单产尤为重要,须通过改良性状来实现高产、稳产(常向楠等,2018;傅向东等,2018)。由于小麦产量与穗部性状高度相关(魏艳丽等,2015),研究小麦穗部相关性状及借助分子标记技术发掘其重要QTL对小麦遗传改良和产量提高均具有重要意义。【前人研究进展】提高小麦产量是其育种的主要目标,其中穗部性状的改良是提高产量的主要途径(Fischer et al.,1998;Ku et al.,1999;魏艳丽等,2015),尤其增加穗粒数和穗粒重具有明显的增产效果(李万昌和刘曙东,2012;Zhang et al.,2012;蔡正正等,2016)。但小麦穗部性状间的关系复杂,多个性状间存在互作效应,无法从某单一性状提高小麦产量(王瑞霞等,2008;Mir et al.,2012)。迄今,已有较多学者在不同遗传背景下对小麦产量及穗部相关性状进行QTL检测和定位研究。Schlegel和Meinel(1994)研究发现,与小穗数相关的QTL位于1BS染色体上。Li等(2007)利用小麦RIL群体对千粒重、穗粒数、单株穗數和小穗数相关的QTL进行定位,结果发现其位于小麦12条染色体上,并分别在1D、2A、6B和7D染色体上检测到相关QTL富集区。Kumar等(2007)研究发现,2个控制小麦穗长的QTL位于2B和2D染色体上,3个控制每穗小穗数的QTL位于2B、4A和6A染色体上。孙中沛等(2017)研究发现,6个与小麦穗长相关的QTL位于2A、2D、3B、4D、5A和7D染色体上,4个与小穗数相关的QTL位于1A、4A和7D染色体上,6个与穗密度相关的QTL位于4D、5A和6B染色体上。王升星等(2017)利用SSR分子标记对小麦单株产量及其他相关性状进行QTL定位分析,结果共发掘出3个与千粒重相关的QTL,分别位于1D(2个)和4B(1个)染色体上,2个与单株产量相关的QTL和2个与有效穗数相关的QTL分别位于染色体1A和5D的相同区间。孙宇慧等(2018)把小麦穗粒数和千粒重主效QTL定位于7AL染色体QC-7AL区。虽然小麦穗部相关性状的QTL研究已有许多报道,但在育种上有利用价值的相关QTL凾待进一步发掘和应用。【本研究切入点】宁夏主栽小麦品种宁春4号具有稳产、适应性广,对麦区主要病害抗性不强等特点;河东乌麦选自山西省运城,具有分蘖力强、总小穗数、穗粒数多、千粒重高,且具有高抗白粉病、中抗条锈病和叶锈病的特点。本课题组前期对宁春4号和河东乌麦的遗传性状与分子标记进行了全面分析(刘妍等,2017;王掌军等,2018,2019),但未系统对其杂交组合群体进行穗部性状分析及其重要QTL发掘。【拟解决的关键问题】以宁春4号和河东乌麦为材料进行正反交,对其F2代进行穗部性状分析及其重要QTL发掘,为宁夏小麦穗部性状的遗传改良提供QTL和育种资源。
1 材料与方法
1. 1 试验材料
供试材料为宁春4号和河东乌麦,均由宁夏大学农学院小麦育种课题组提供。主要试剂:Taq DNA聚合酶、10×Taq Buffer(含Mg2+)和dNTPs均购自康为世纪(北京)生物科技有限公司;甲叉双丙烯酰胺(C7H10N2O2)、丙烯酰胺(C3H5NO)和SDS均购自国安生物科技(西安)有限公司;其他生化试剂均购自生工生物工程(上海)股份有限公司。主要设备仪器:ChemiDoc XRS+凝胶成像系统(BIO-RAD,美国)、微量核酸蛋白检测仪(ND-2000型,美国)、PTC-200 PCR仪(BIO-RAD,美国)、移液器(Eppendorf,德国)、台式高速冷冻离心机(Neofuge 15型,上海安亭科学仪器厂)、电泳仪(DYCZ-30C型,北京市六一仪器厂)。
1. 2 组配杂交
2016年在宁夏大学教学实验农场组配宁春4号与河东乌麦正反交组合,同年将收获的杂交种子在云南元谋南繁加代,2017年将收获种子种植于宁夏大学教学实验农场,单粒点播,每行10粒,行长1.1 m,行宽0.2 m,获得包含331个单株的F2后代群体,其中正交201株(编号1~201),反交130株(编号202~331)。
1. 3 穗部性状指标测定
在成熟期进行室内考种(均为每个单株所有有效穗数在该性状的平均值):穗长采用直接测量法,用卷尺测量;总小穗数为每穗上所有的小穗数目,不孕小穗数为各小花均不结实的小穗数目,结实小穗数为每穗均结实的小穗数,均采用直接观测计数法,结实小穗数=总小穗数-不孕小穗数;穗粒数和穗粒重为每穗的粒数和粒重量。
1. 4 分子标记设计及合成
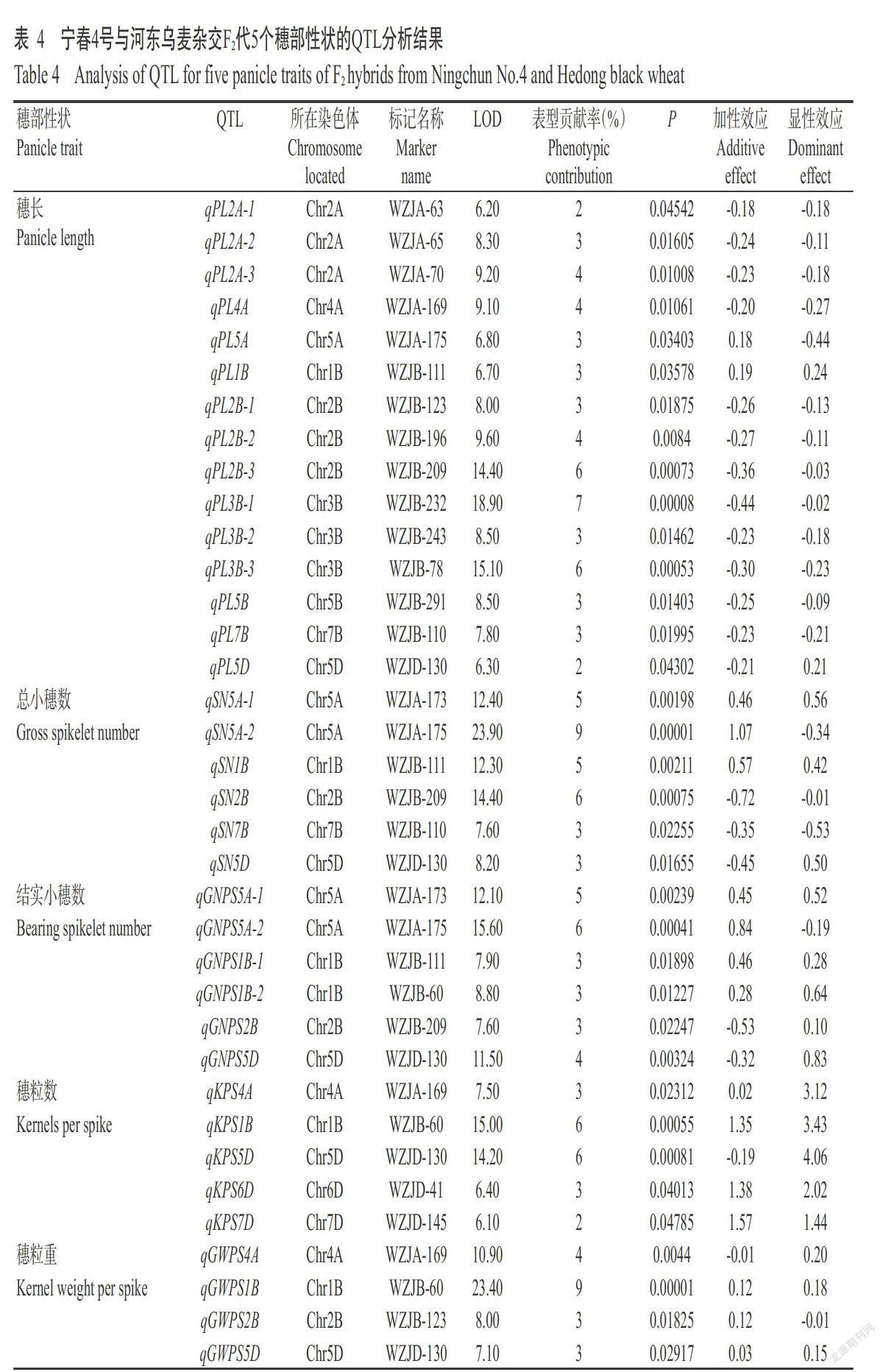
刘妍等(2017)根据小麦7个部分同源群小卫星区设计SSR分子标记,在宁春4号和河东乌麦中筛选出127个具有多态性的SSR分子标记。本研究利用在宁春4号和河东乌麦中稳定扩增的49个SSR分子标记进行分析,其中有19个SSR分子标记对穗部性状有明确QTL定位结果,其引物信息见表1。所有引物均由生工生物工程(上海)股份有限公司合成。
1. 5 PCR检测
采用SDS法提取供试材料苗期叶片的基因组DNA(Yang et al.,2006)。PCR反应体系和扩增程序参见王掌军等(2019)的方法,退火温度如表1所示。扩增产物采用8%聚丙烯酰胺凝胶电泳检测,经银染后观察拍照,并统计扩增条带。
1. 6 统计分析
采用Excel 2013对穗部性状和分子标记扩增条带进行统计。利用SPSS 20.0进行方差、相关性及聚类分析。利用Origin 8.0对各穗部性状的频率分布进行作图。利用Map Manager QTXb 20进行QTL定位,取概率值P<0.05的LOD作为判断QTL存在的阈值。参照王掌军等(2019)的方法,利用Kosambi函数中单标记回归分析方法进行QTL分析。运用Map Manager QTXb20检测F2代单株穗部性状相关QTL位点,QTL命名为:q+性状的大写字母代称+所在染色体号数,如在2A染色体上,与穗长相关的QTL位点命名为“qPL2A”。
2 结果与分析
2. 1 宁春4号与河东乌麦杂交F2代穗部性状分析结果
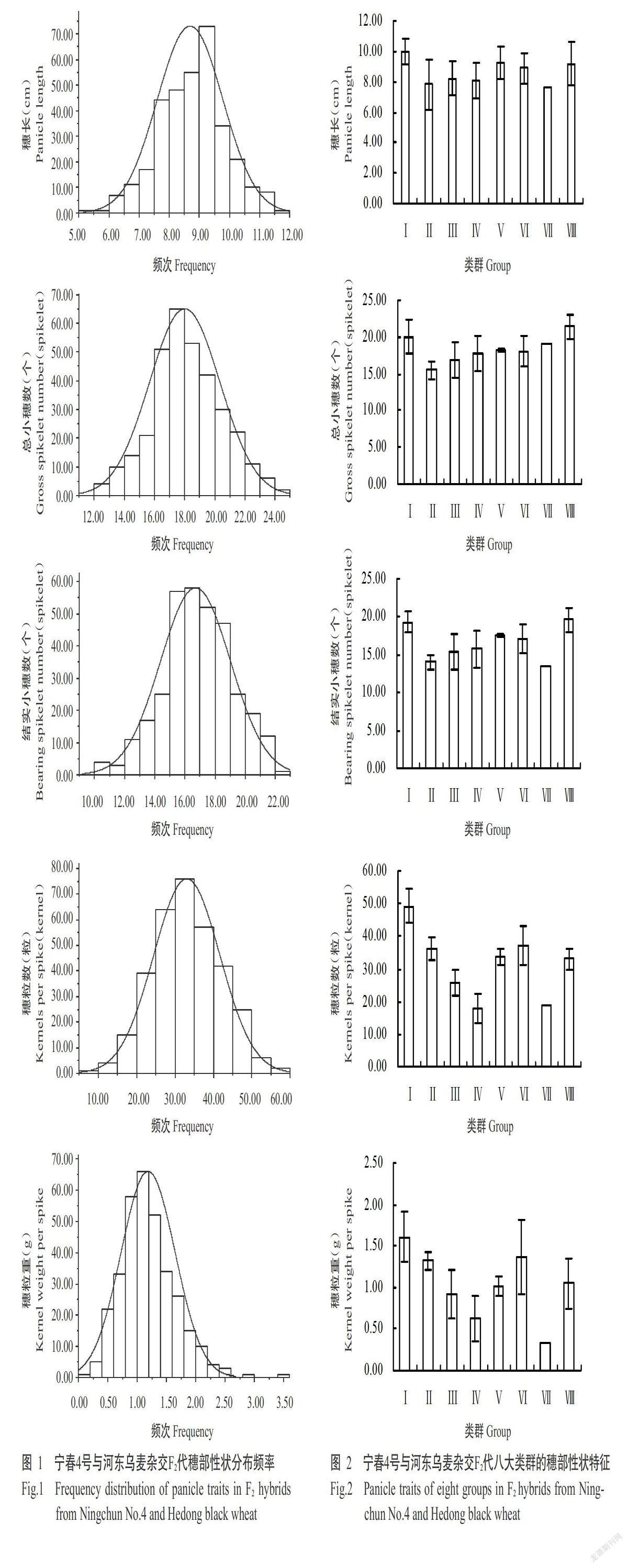
2. 1. 1 穗部性状变异性分析 由图1可知,5个穗部性状在F2代群体中均出现较大分离,分布范围均呈连续正态分布,符合多基因控制的数量性状遗传特点,且出现许多具有超亲性状的单株。对F2代群体穗部性状进行变异性分析,结果(表2)表明,总小穗数和结实小穗数的群体平均值介于高亲亲本与低亲亲本之间,穗长、穗粒数和穗粒重的群体平均值均低于低亲亲本,其中,穗长、总小穗数、结实小穗数、穗粒数和穗粒重的超中亲比例分别达20.24%、44.71%、41.99%、34.14%和33.84%,超高亲比例分别达6.64%、18.73%、17.22%、34.14%和26.88%。5个穗部性状的变异系数表现为穗粒重(38.96%)>穗粒数(26.12%)>结实小穗数(13.92%)>总小穗数(12.97%)>穗长(12.52%),平均变异系数为20.90%。综上所述,F2代群体的5个穗部性状变异较大。
2. 1. 2 基于穗部性状的聚类分析 根据穗长、总小穗数、结实小穗数、穗粒数和穗粒重5个穗部性状测定结果对F2代群体进行聚类分析,在遗传距离为5 cM时,F2代群体被分成八大类群(图2),其中,类群Ⅰ~Ⅷ的单株数分别为8、2、81、23、2、195、1和19个。同时,八大类群的穗长表现为Ⅰ>Ⅴ>Ⅷ>Ⅵ>Ⅲ>Ⅳ>Ⅱ>Ⅶ,总小穗数表现为Ⅷ>Ⅰ>Ⅶ>Ⅴ>Ⅵ>Ⅳ>Ⅲ>Ⅱ,结实小穗数表现为Ⅷ>Ⅰ>Ⅴ>Ⅵ>Ⅳ>Ⅲ>Ⅱ>Ⅶ,穗粒数表现为Ⅰ>Ⅵ>Ⅱ>Ⅴ>Ⅷ>Ⅲ>Ⅶ>Ⅳ,穗粒重表现为Ⅰ>Ⅵ>Ⅱ>Ⅷ>Ⅴ>Ⅲ>Ⅳ>Ⅶ。总体上,类群Ⅰ平均穗长最长(9.96 cm)、穗粒数最多(49.25粒/穗)、穗粒重最重(1.61 g),类群Ⅷ平均总小穗数和结实小穗数最多,分别为21.40和19.57个。综上所述,类群Ⅰ和Ⅷ为穗部性状优异类群。
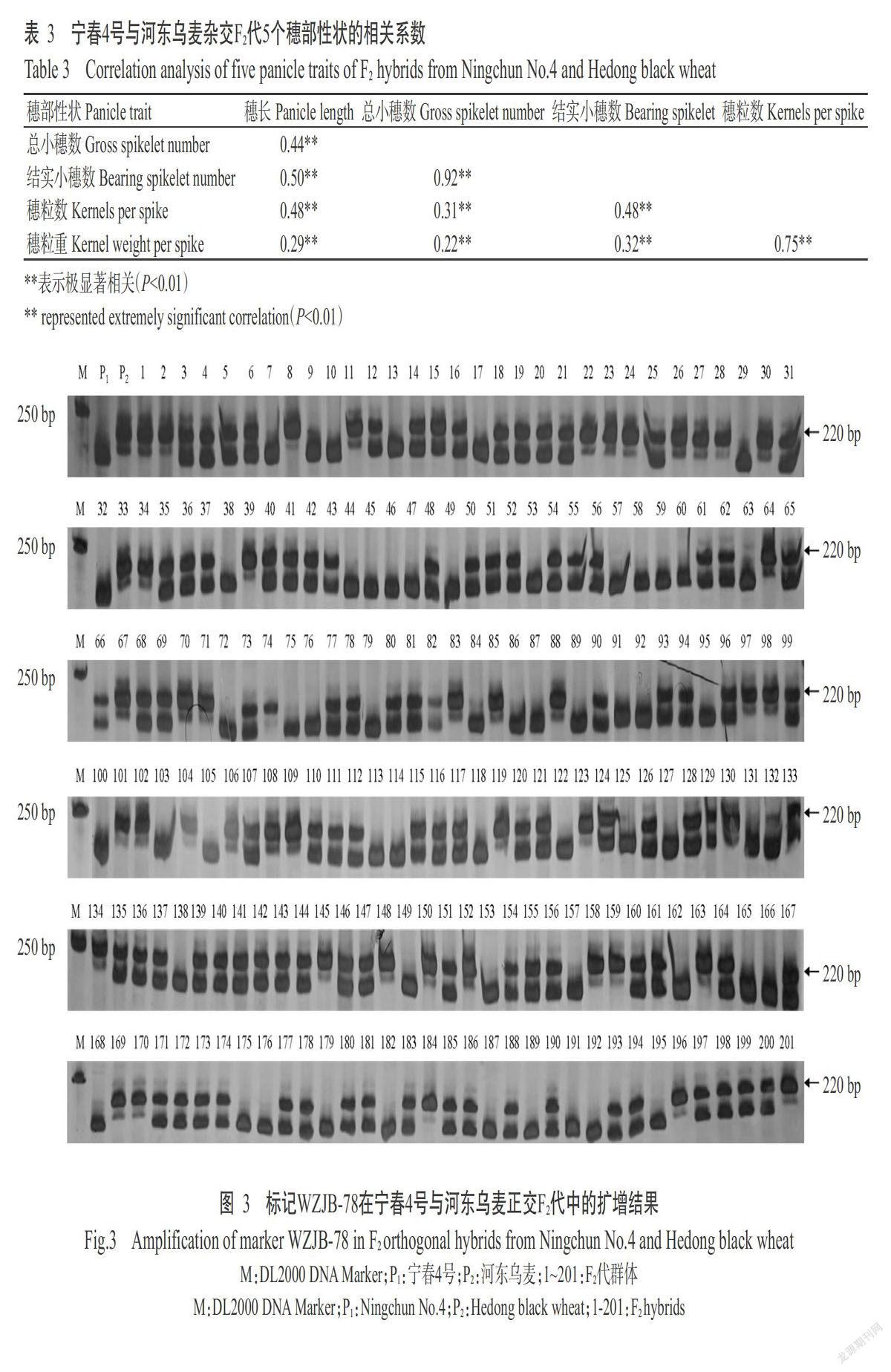
2. 1. 3 穗部性状的相关性分析 对F2代群体的5个穗部性状进行相关性分析,结果如表3所示。穗部性状间均呈极显著正相关(P<0.01),相关系数排序为总小穗数与结实小穗数>穗粒数与穗粒重>穗长与结实小穗数>穗粒数与穗长=穗粒数与结实小穗数>穗长与总小穗数>结实小穗数与穗粒重>总小穗数与穗粒数>穗长与穗粒重>总小穗数与穗粒重,表明这些性状对小麦产量贡献均较大。
2. 2 宁春4号与河东乌麦杂交F2代穗部性状的QTL分析结果
利用在宁春4号和河东乌麦中稳定扩增的49个SSR分子标记对F2代群体进行检测,部分检测结果如图3所示,其中,有19个标记对穗部性状有明确的QTL定位结果,如表4所示。19个SSR分子标记共检测到36个与穗部性状相关的QTL,加性效应为-0.72~1.57,表型贡献率为2%~9%,LOD最大值为23.90,分布在2A、4A、5A、1B、2B、3B、5B、7B、5D、6D和7D等11条染色体上。其中,与穗长相关的QTL 15个,分布在2A、4A、5A、1B、2B、3B、5B、7B和5D等9条染色体上,加性效应为-0.44~0.19,表型贡献率为2%~7%,LOD最大值为18.90;与总小穗数相关的QTL 6个,分布在5A、1B、2B、7B和5D等5条染色体上,加性效应为-0.72~1.07,表型贡献率为3%~9%,LOD最大值为23.90;与结实小穗数相关的6个QTL,分布在5A、1B、2B和5D等4条染色体上,加性效应为-0.53~0.84,表型贡献率为3%~6%,LOD最大值为15.60;与穗粒数相关的QTL 5个,分布在4A、1B、5D、6D和7D等5条染色体上,加性效应为-0.19~1.57,表型贡献率为2%~6%,LOD最大值为15.00;与穗粒重相关的QTL 4个,分布在4A、1B、2B和5D等4条染色體上,加性效应为-0.01~0.12,表型贡献率为3%~9%,LOD最大值为23.40。可见,4A染色体上可检测到穗长、穗粒数和穗粒重QTL;5A染色体上可检测到穗长、总小穗数和结实小穗数QTL;1B和5D染色体上均能检测到穗长、总小穗数、结实小穗数、穗粒数和穗粒重QTL;2B染色体上可检测到穗长、总小穗数、结实小穗数和穗粒重QTL;7B染色体上可检测到穗长和总小穗数QTL,说明4A、5A、1B、2B、7B和5D等6条染色体存在不同穗部性状的QTL富集区。
3 讨论
小麦生产在保障我国粮食安全中具有重要地位,培育高产、稳产和适应性强的新品种是目前小麦育种的主要目标。大量研究表明,禾谷类作物的穗长、小穗数、穗粒数、千粒重等遗传力较高,其遗传变异受环境影响较小,应从早期世代开始进行选择(刘广田等,1990;Ma et al.,2007;彭丁文,2011)。由于杂交F2代群体各遗传性状处于高度分离状态,能够提供最大的数量遗传信息量,是早期世代选择优系的理想群体。本研究结果也表明,穗部性状指标在F2代群体中均出现较大分离,分布范围均呈连续正态分布,属于多基因控制的数量性状遗传;穗长、总小穗数、结实小穗数、穗粒数和穗粒重的超中亲比例分别达20.24%、44.71%、41.99%、34.14%和33.84%,超高亲比例分别达6.64%、18.73%、17.22%、34.14%和26.88%,5个穗部性状的变异系数较大,表现为穗粒重>穗粒数>结实小穗数>总小穗数>穗长,与柴永峰等(2013)、王升星等(2017)的研究结果基本一致,但与王光禄等(2016)报道的部分性状的变异系数略有差异,可能是不同研究中供试的亲本不同,或群体大小、世代数高低和考察的性状数量不同所致。同时,穗长、总小穗数、结实小穗数、穗粒数和穗粒重等5个穗部性状间均呈不同程度极显著正相关,说明这些性状对小麦产量贡献均较大,与前人研究结果(Wu et al.,2014)基本一致。此外,本研究331个F2代可分为八大类群,其中,类群Ⅰ平均穗长最长(9.96 cm),穗粒数最多(49.25粒),穗粒重最重(1.61 g),类群Ⅷ平均总小穗数和结实小穗数最多(21.40和19.57个)。可见,小麦各穗部性状间存在复杂关系,在育种中不能只靠改良某一性状达到高产目标,而应兼顾各性状间的互作效应(王瑞霞等,2008;Mir et al.,2012)。
目前,SSR分子标记已广泛应用于小麦一些重要穗部性状QTL研究(张坤普等,2009;Li et al.,2010;McIntyre et al.,2010;Deng et al.,2011;Zhang et al.,2012)。本研究利用新开发19个SSR分子标记共检测到36个与穗部性状相关的QTL,LOD最大值为23.90,加性效应变幅为-0.72~1.57,表型贡献率为2%~9%,分布在2A、4A、5A、1B、2B、3B、5B、7B、5D、6D和7D等11条染色体上。与前人研究(Kumaret al.,2007;孙中沛等,2017)相比发现,本研究检测到的部分穗部性状QTL位点与前人研究发现的部分穗部性状QTL位于相同染色体上,且本研究在4A、5A、1B、2B、5B、7B、5D、6D和7D等9条染色体上新挖掘了26个QTL,可能是本研究所使用的分子标记和作图群体与前人研究不同所致。结合前人研究结果(Liu et al.,2006;Yu et al.,2014;Hu et al.,2017;Xu et al.,2017)发现,小麦21条染色体上分布重要穗部性状的QTL,其中,3D染色体上分布的穗部性状QTL最少。同时,Li等(2007)研究发现,1D、2A、6B和7D染色体上存在不同穗部性状QTL富集区。本研究在4A染色体上检测到穗长、穗粒数和穗粒重QTL,5A染色体上检测到穗长、总小穗数和结实小穗数QTL,1B染色体上检测到穗长、总小穗数、结实小穗数、穗粒数和穗粒重QTL,2B染色体上检测到穗长、总小穗数、结实小穗数和穗粒重QTL,7B染色体上检测到穗长和总小穗数QTL,5D染色体上检测到穗长、总小穗数、结实小穗数、穗粒数和穗粒重QTL。综上所述,4A、5A、1B、2B、7B和5D等6条染色体存在穗部性状QTL富集区,可作为下一步研究的重点。另外,Liu等(2018)利用SNP分子标记检测到11个与穗部性状相关的QTL,表型贡献率为5.4%~37.7%。由于本研究中可供利用的分子标记数目相对较少,在今后研究中,还需开发高效的分子标记,通过构建密度较饱和的物理图谱,进一步发掘和精细定位穗部性状QTL,尤其是主效QTL,为小麦重要穗部性状改良打下分子基础。
4 结论
小麦杂交F2代遗传性状处于高度分离,蕴藏着最大的数量遗传信息,为相关穗部性状分析及QTL发掘提供了可靠的遗传群体,检测到的QTL可用于小麦穗部性状遗传改良。
参考文献:
蔡正正,陈学群,唐唯其,李文强,吴为人,段远霖. 2016. 两种水稻短穗突变体的形态特征、遗传分析和基因定位[J]. 基因组学与应用生物学,35(5):1183-1188. [Cai Z Z,Chen X Q,Tang W Q,Li W Q,Wu W R,Duan Y L. 2016. Morphological characteristics,genetic analysis and gene location of two short panicle mutants of rice[J]. Genomics and Applied Biology,35(5):1183-1188.]
柴永峰,李秀绒,赵智勇,孙来虎,李永山,景力武. 2013. 国外小麦种质资源农艺性状及品质性状的多样性分析[J]. 农学学报,3(9):1-8. [Chai Y F,Li X R,Zhao Z Y,Sun L H,Li Y S,Jing L W. 2013. Diversity analysis on agronomic and quality characters of foreign wheat germplasm resources[J]. Journal of Agriculture,3(9):1-8.]
常向楠,陈树林,程西永,许海霞,朱小涛,李林繁,望俊森,楊秋云,詹克慧. 2018. 稀植和密植下小麦主要农艺性状的遗传差异及关系分析[J]. 河南农业大学学报,52(4):497-505. [Chang X N,Chen S L,Cheng X Y,Xu H X,Zhu X T,Li L F,Wang J S,Yang Q Y,Zhan K H. 2018. Genetic differences and correlation of main agronomic traits in wheat under low-density and high-density planting models[J]. Journal of Henan Agricultural University,52(4):497-505.]
傅向东,刘倩,李振声,张爱民,凌宏清,童依平,刘志勇. 2018. 小麦基因组研究现状与展望[J]. 中国科学院院刊,33(9):909-913. [Fu X D,Liu Q,Li Z S,Zhang A M,Ling H Q,Tong Y P,Liu Z Y. 2018. Research achievement and prospect development on wheat genome[J]. Bulletin of Chinese Academy of Sciences,33(9):909-913.]
何中虎,庄巧生,程顺和,于振文,赵振东,刘旭. 2018. 中国小麦产业发展与科技进步[J]. 农学学报,8(1):99-106. [He Z H,Zhuang Q S,Cheng S H,Yu Z W,Zhao Z D,Liu X. 2018. Wheat production and technology improvement in China[J]. Journal of Agriculture,8(1):99-106.]
李冬兵,左宁,吕桂珍,杜红旭,牛洪斌,尹钧. 2019. 不同小麦品种穗发芽特性的鉴定及TaRHA2b基因序列差异分析[J]. 河南农业科学,48(1):6-16. [Li D B,Zuo N,Lü G Z,Du H X,Niu H B,Yin J. 2019. Identification of charac-teristics of pre-harvest sprouting and differences of TaRHA2b gene sequence[J]. Journal of Henan Agricultu-ral Sciences,48(1):6-16.]
李万昌,刘曙东. 2002. 强优势杂交小麦产量结构优势间关系的研究[J]. 麦类作物学报,22(3):1-6. [Li W C,Liu S D. 2002. The relationship among yield structure heterosis of strong heterosis hybrid wheat[J]. Journal of Triticeae Crops,22(3):1-6.]
刘妍,田蓉蓉,王天佑,刘凤楼,亢玲,刘生祥,张晓岗,刘萍,王掌军. 2017. 2个小麦品种的遗传性状分析及分子标记筛选[J]. 河南农业科学,46(10):13-20. [Liu Y,Tian R R,Wang T Y,Liu F L,Kang L,Liu S X,Zhang X G,Liu P,Wang Z J. 2017. Analysis on related genetic traits and screening of molecular marker in two wheat varieties[J]. Journal of Henan Agricultural Science,46(10):13-20.]
刘广田,朱金宝,张树榛. 1990. 普通小麦籽粒品质性状研究Ⅱ.籽粒品质性状与农艺性状的相关[J]. 北京农业大学学报,16(3):255-261. [Liu G T,Zhu J B,Zhang S Z. 1990. Study on quality and agronomic characters in Triticum aestivum L.Ⅱ. correlations between quality and agronomic characters[J]. Acta Agriculture Universitatis Pekinensis,16(3):255-261.]
刘志勇,王道文,张爱民,梁翰文,吕慧颖,邓向东,葛义强,魏珣,杨维才. 2018. 小麦育种行业创新现状与发展趋势[J]. 植物遗传资源学报,19(3):430-434. [Liu Z Y,Wang D W,Zhang A M,Liang H W,Lü H Y,Deng X D,Ge Y Q,Wei X,Yang W C. 2018. Present situation and deve-lopment trend of wheat breeding industry innovation[J]. Journal of Plant Genetic Resources,19(3):430-434.]
彭丁文. 2011. 水稻粒重遺传与育种研究进展[J]. 南方农业学报,42(3):250-252. [Peng D W. 2011. Research progress on the utilization of rice grain weight traits in molecu-lar breeding[J]. Journal of Southern Agriculture,42(3):250-252.]
孙宇慧,刘天相,石善党,丁梦云,高欣,王中华,李春莲. 2018. 小麦穗粒数及千粒重主效QTL共定位区QC-7AL的精细定位及遗传效应分析[J]. 麦类作物学报,38(11):1288-1292. [Sun Y H,Liu T X,Shi S D,Ding M Y,Gao X,Wang Z H,Li C L. 2018. Fine mapping of a major QTL cluster QC-7AL and the effect on kernels number per spike and thousand kernels weight in wheat(Triticum aestivum L.)[J]. Journal of Triticeae Crops,38(11):1288-1292.]
孙中沛,刘天相,左希亚,赵琛,王中华,李春莲. 2017. 普通小麦穗部性状QTL分析[J]. 麦类作物学报,37(4):452-457. [Sun Z P,Liu T X,Zuo X Y,Zhao C,Wang Z H,Li C L. 2017. QTL mapping of spike related traits in common wheat[J]. Journal of Triticeae Crops,37(4):452-457.]
王光禄,刘志宏,程倩倩,冯盛烨,闫树平,于洋,赵杨,王怀恩,孙允超. 2016. 94份国外小麦种质材料的主要农艺性状分析[J]. 麦类作物学报,36(5):577-582. [Wang G L,Liu Z H,Cheng Q Q,Feng S Y,Yan S P,Yu Y,Zhao Y,Wang H E,Sun Y C. 2016. Analysis on main agronomic characters of 94 foreign wheat germplasms[J]. Journal of Triticeae Crops,36(5):577-582.]
王瑞霞,张秀英,伍玲,王瑞,海林,闫长生,游光霞,肖世和. 2008. 不同生态环境条件下小麦籽粒灌浆速率及千粒重QTL分析[J]. 作物学报,34(10):1750-1756. [Wang R X,Zhang X Y,Wu L,Wang R,Hai L,Yan C S,You G X,Xiao S H. 2008. QTL mapping fir grain filling rate and thousand-grain weight in different ecological environments in wheat[J]. Acta Agronomica Sinica,34(10):1750-1756.]
王升星,牛影,陈聪灵,郑乐,马欢欢,时曼丽,秦学锋,黄陈,朱玉磊,张海萍,卢杰,常成,马传喜. 2017. 小麦单株产量及相关性状的全基因组QTL分析[J]. 安徽农业大学学报,44(4):720-725. [Wang S X,Niu Y,Chen C L,Zheng L,Ma H H,Shi M L,Qin X F,Huang C,Zhu Y L,Zhang H P,Lu J,Chang C,Ma C X. 2017. Genome-wide QTL analysis for wheat grain yield per plant and other related traits[J]. Journal of Anhui Agricultural University,44(4):720-725.]
王掌军,刘妍,王姣,付青青,刘凤楼,张双喜,张文杰,张晓岗,刘生祥. 2018. 小麦种质资源农艺性状遗传分析及白粉病抗性鉴定[J]. 西南农业学报,31(7):1338-1348. [Wang Z J,Liu Y,Wang J,Fu Q Q,Liu F L,Zhang S X,Zhang W J,Zhang X G,Liu S X. 2018. Genetic analysis on agronomic traits and identification of powdery mildew resistance of wheat germplasm resources[J]. Southwest China Journal of Agricultural Science,31(7):1338-1348.]
王掌军,刘妍,李阳,杨乔乔,田蕊,亢玲,张晓岗,刘萍,刘生祥. 2019. 宁春4号与河东乌麦杂交F2品质性状及其分子标记分析[J]. 河南农业科学,48(2):17-24. [Wang Z J,Liu Y,Li Y,Yang Q Q,Tian R,Kang L,Zhang X G,Liu P,Liu S X. 2019. Analysis of quality traits and their molecular markers of F2 hybrids from Ningchun No.4 and Hedong black wheat[J]. Journal of Henan Agricultural Sciences,48(2):17-24.]
魏艷丽,王彬龙,李瑞国,蒋会利,张安静. 2015. 大穗小麦穗部性状的遗传分析[J]. 麦类作物学报,35(10):1366-1371. [Wei Y L,Wang B L,Li R G,Jiang H L,Zhang A J. 2015. Genetic analysis on spike characteristics of wheat variety with large spike[J]. Journal of Triticeae Crops,35(10):1366-1371.]
张坤普,徐宪斌,田纪春. 2009. 小麦籽粒产量及穗部相关性状的QTL定位[J]. 作物学报,35(2):270-278. [Zhang K P,Xu X B,Tian J C. 2009. QTL mapping for grain yield and spike related traits in common wheat[J]. Acta Agronomica Sinica,35(2):270-278.]
朱昀,李朝炜,刘颖,魏景芳. 2019. 小麦VP基因的克隆、生物信息学分析及功能初探[J]. 河南农业科学,48(2):8-16. [Zhu Y,Li Z W,Liu Y,Wei J F. 2019. Cloning,bioinformatics analysis and preliminary functional study of wheat VP gene[J]. Journal of Henan Agricultural Sciences,48(2):8-16.]
Deng S,Wu X,Wu Y,Zhou R,Wang H,Jia J,Liu S. 2011. Characterization and precise mapping of a QTL increa-sing spike number with pleiotropic effects in wheat[J]. Theoretical and Applied Genetics,122(2):281-289.
Fischer R A,Rees D,Sayre K D,Lu Z M,Condon A G,Saavedra L A. 1998. Wheat yield progress associated with high stomatal conductance and photosynthetic rate and cooler canopies[J]. Crop Science,278(6):1467-1475.
Hu Y S,Ren T H,Li Z,Tang Y Z,Ren Z L,Yan B J. 2017. Molecular mapping and genetic analysis of a QTL controlling spike formation rate and tiller number in wheat[J]. Gene,643:15-21.
Ku M B S,Sakae A,Mika N,Hiroshi F,Hiroko T,Kazuko H,Seichi T,Mitsue M,Makoto M. 1999. High-level expre-ssion of maize phosphoenolpy ruvate carboxylase in transgenic rice plants[J]. Nature Biotechnology,17(1):76-80.
Kumar N,Kulwal P L,Balyan H S. 2007. QTL mapping for yield and yield contributing traits in two mapping populations of bread wheat[J]. Molecular Breeding,19(2):163-177.
Li S S,Jia J Z,Wei X Y,Zhang X C,Li L Z,Chen H M,Fan H Y,Zhao X H,Lei T D,Xu Y F,Jiang F S,Wang H G,Li L H. 2007. A intervarietal genetic map and QTL analysis for yield traits in wheat[J]. Molecular Genetics and Genomics,123(20):167-178.
Li Z,Peng T,Xie Q,Han S,Tian J. 2010. Mapping of QTL for tiller number at different stages of growth in wheat using double haploid and immortalized F2 populations[J]. Journal of Genetics and Genomics,89(4):409-415.
Liu K,Sun X X,Ning T Y,Duan X X,Wang Q L,Liu T T,An Y L,Guan X,Tian J C,Chen J S. 2018. Genetic disse-ction of wheat panicle traits using linkage analysis and a genome-wide association study[J]. Theoretical and App-lied Genetics,131(5):1073-1090.
Liu S B,Zhou R H,Dong Y C,Li P,Jia J Z. 2006. Development,utilization of introgression lines using a synthetic wheat as donor[J]. Theoretical and Applied Genetics,112(7):1360-1373.
Ma Z Q,Zhao D M,Zhang C Q,Zhang Z Z,Xue S L,Lin F,Kong Z X,Tian D G,Luo Q Y. 2007. Molecular genetic analysis of five spike-related traits in wheat using RIL and immortalized F2 populations[J]. Molecular Genetics and Genomics,277(1):31-42.
McIntyre C L,Mathews K L,Rattey A,Chapman S C,Drenth J,Ghaderi M,Shorter R. 2010. Molecular detection of genomic regions associated with grain yield and yield-rela-ted components in an elite bread wheat cross evaluated underirrigated and rainfed conditions[J]. Theoretical and Applied Genetics,120(3):527-541.
Mir R R,Kumar N,Jaiswal V,Girfharwal N,Prasad M,Bal-yan H S,Gupta P K. 2012. Genetic dissection of grain weight in bread wheat through quantitative trait locus interval and association mapping[J]. Molecular Breeding,29(4):963-972.
Schlegel R,Meinel A. 1994. A quantitative trait locus(QTL) on chromosome arm 1RS of rye and its effect on yield performance of hexaploid wheat[J]. Cereal Research Communications,22(1/2):7-13.
Wu X Y,Cheng R R,Xue S L,Kong Z X,Wan H S,Li G Q,Huang Y L,Jia H Y,Jia J Z,Ma Z Q. 2014. Precise map-ping of a quantitative trait locus interval for spike length and grain weight in breed wheat(Triticum aestivum L.)[J]. Molecular Breeding,14(33):129-138.
Xu T,Bian N,Wen M,Xiao J,Yuan C,Cao A,Zhang S,Wang X,Wang H. 2017. Characterization of a common wheat(Triticum aestivum L.) high-tillering dwarf mutant[J]. Theoretical and Applied Genetics,130(3):483-494.
Yang Z J,Li G R,Chang Z J,Zhou J P,Ren Z L. 2006. Charac-terization of a partial amphiploid between Triticum aestivum cv Chinese spring and Thinopyrum intermedium ssp. Trichophorum[J]. Euphytica,149(1/2):11-17.
Yu M,Chen G Y,Zhang L Q,Liu Y X,Liu D C,Wang J R,Pu Z E,Zhang L,Lan X J,Wei Y M,Liu C J,Zheng Y L. 2014. QTL mapping for important agronomic traits in synthetic hexaploid wheat derived from Aegiliops tauschii ssp. tauschii[J]. Journal of Integrative Agriculture,13(9):1835-1844.
Zhang D L,Hao C Y,Wang L F,Zhang X Y. 2012. Identif-ying loci influencing grain number by microsatellite scree-ning in bread wheat(Triticum aestivum L.)[J]. Planta,236(5):1507-1517.
(責任编辑 陈 燕)
猜你喜欢
农民致富之友(2020年8期)2020-05-11
农民致富之友(2019年32期)2019-11-23
清明(2019年6期)2019-11-21
新农村(2017年17期)2017-08-23
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
山东农业科学(2016年12期)2017-01-21
天津农业科学(2016年12期)2017-01-11
天津农业科学(2015年5期)2015-05-30
