
猪源粪肠球菌菌毛亚单位蛋白多抗对其生物被膜形成及黏附侵袭作用的影响
2020-03-25 09:33王芳冯艳红董家君王亚宾段志刚
河南农业大学学报 2020年1期
王芳, 冯艳红, 董家君, 王亚宾, 段志刚
(1. 河南农业大学牧医工程学院,河南 郑州 450002;2.郑州市动物卫生监督所,河南 郑州 450007)
肠球菌(Enterococcus)是革兰氏阳性、兼性厌氧球菌,是人体肠道内的常在菌, 也是医院内的感染主要机会致病菌[1-3]。在肠球菌的感染中,粪肠球菌占据了80%左右[4],而屎肠球菌和其他肠球菌仅占20%左右。由于肠球菌具有天然和获得性抗性能力,对现有的抗生素均产生了耐药性[5]。如果再考虑到肠球菌对热和消毒药物的抗性[6],这就使得寻求肠球菌治疗新的选项和预防变得越来越重要,而发现在体内表达的重要毒力因子的蛋白就成为免疫治疗和免疫预防的重要目标[7]。由于对肠球菌主要毒力因子和分子致病机制研究的有限性,限制了对肠球菌免疫的治疗和免疫预防[6]。肠球菌的Cyl、gelE、As、ace、efaA、sprE、esp和性信息素等均已被公认为毒力因子,它们在对宿主细胞吸附、抵抗吞噬细胞杀死、损伤组织细胞等方面发挥着不同的作用,但由于它们的携带率或遗传结构的多样性等问题限制了它们的应用。BAKSHI等[8]利用比较基因组学方法评估粪肠球菌菌株的潜在致病性,并确定了一个粪肠球菌毒力岛富集模块。研究表明,粪肠球菌毒力因子编码的Ebp、Esp、GelE、AS、Ace等均与生物被膜有关[9]。粪肠球菌 ebp PGCs由4个完整的开放阅读框组成,编码Ebp菌毛的因位于染色体中,分布广泛,保守性高[10],分别编码ebpA、ebpB、ebpC菌毛亚单位和srtC分拣酶[11]。EbpA、EbpB、EbpC作为粪肠球菌菌毛相关操纵子,与其菌毛的形成、黏附侵入机体和生物被膜的形成有关[12]。细菌菌毛在黏附定植宿主上皮细胞,在引发机体感染等过程中发挥着重要的作用,并且菌毛蛋白作为一种细菌表面蛋白结构具有较好的免疫原性,能够刺激机体产生相对应的特异性抗体[10,13-14]。对临床分离菌株菌毛的研究发现,粪肠球菌Ebp菌毛蛋白能介导对胶原蛋白、纤维蛋白原和血小板的黏附,进而引起心内膜炎,且在生物膜的形成过程中也扮演着重要作用[15]。因此,Ebp菌毛蛋白有望成为一个良好的免疫原。
粪肠球菌主要通过黏附、侵袭和细胞损伤导致感染发生。其中,黏附是感染的关键一步,而且, 粪肠球菌在感染的过程中可以形成生物被膜,生物被膜给粪肠球菌提供了生长与繁殖的天然的屏障与保护作用,并能增强其致病性[16]。作者在前期研究中,对猪源粪肠球菌Ebp菌毛进行了克隆和序列分析,发现EbpA、EbpB和EbpC有6个与免疫相关的抗原表位,将这6个抗原表位分别进行克隆、表达、纯化和免疫新西兰大白兔后,初步验证了这6个Ebp菌毛抗原亚单位蛋白的免疫源性。为了进一步验证Ebp菌毛亚单位片段的对粪肠球菌的免疫保护作用,将这6个Ebp菌毛亚单位多抗血清和纯化的IgG分别进行粪肠球菌生物膜形成、黏附侵入阻断试验,以筛选出对粪肠球菌致病作用有明显影响的菌毛亚单位蛋白片段,从而为粪肠球菌的治疗和免疫预防以及感染诊断奠定基础。
1 材料与方法
1.1 菌株、细胞、试验动物与菌毛蛋白多克隆抗血清
粪肠球菌N9和N41菌株(分离自河南地区送检病死猪内脏,并经Vitek-32全自动细菌鉴定系统及16S rRNA测序鉴定到种水平的菌株,-80 ℃下保存在50%甘油中)、猪肠道上皮细胞(IPEC-J2)均为河南省动物性食品安全重点实验室保存。动物源性粪肠球菌Ebp菌毛蛋白多克隆抗血清为河南省动物性食品安全重点实验室原核表达菌毛蛋白EbpA片段(EbpA1、EbpA2和EbpA3)、EbpB片段(EF1092A)、EbpC片段(EbpC1、EbpC2)、免疫清洁级新西兰大白兔制备,经双向琼脂扩散试验检测蛋白EbpA1、EbpA2、EbpA3、 EF1092A、EbpC1和EbpC2的抗血清效价分别高达1∶16、1∶8、1∶32、1∶64、1∶32和1∶64,通过Western Blot检测多抗血清具有很好的特异性,收集的血清加入防腐剂分装后-80 ℃保存。
1.2 多克隆抗体血清的纯化
将多克隆抗血清用饱和硫酸铵盐析方法粗提取的IgG,加入到经预处理的DEAE-52纤维素层析柱中,然后用洗脱液(pH=6.4的0.017 5 mol·L-1PBS)进行洗脱并收集。用以ND-1000型微量紫外分光光度计测定其含量,-80 ℃保存。
1.3 生物被膜形成和阻断试验
生物被膜形成试验参照文献[17]略有改进。将保存的菌株在BHI液体培养基培养并调整菌液浓度为1. 5×108CFU·mL-1。用TSB+0.25%葡萄糖培养基将菌液稀释100倍后加入到96孔聚苯乙烯板中,100 μL·孔-1,5株·孔-1,在36 h时用PBS缓冲液洗板3次,晾干并用甲醇固定。用1%结晶紫染色定量生物膜的产生并用分光光度计(570 nm)测定其吸光度。而对生物被膜阻断试验基本同生物被膜形成试验,仅在加入TSB+0.25%葡萄糖培养的菌株同时,加入优化的100 μL·孔-1的多抗血清或IgG充分混匀,余下同生物被膜形成试验。并用含有TSB+0.25%葡萄糖培养基作为空白对照试验。
1.4 兔多抗对细胞黏附、侵袭的影响
1.4.1 菌株、细胞的培养 将N9、N41菌种划线于BHI琼脂平板上,37 ℃温箱培养24~48 h。挑取单个菌落接种于BHI培养基中培养至OD600 nm值为0.8。黏附、侵袭试验前,将细菌培养至对数中期,8 000 r·min-1离心5 min,PBS液洗3次,用无血清无双抗的RPMI-1640培养基稀释细菌至1×108CFU·mL-1备用。使用IPEC-J2细胞生长至单层细胞时用于细菌的黏附与侵袭的培养。
1.4.2 细菌的黏附、侵袭细胞试验 细菌的黏附、侵袭试验按文献[18-19]的方法稍加改动。待生长至单层的细胞加入108cfu·mL-1菌悬液100 μL(MOI:100∶1),置37 ℃、5% CO2培养箱中孵育1.5 h后,弃培养基,D-HanK’s液洗涤3次,随后进行黏附、侵袭试验。在黏附试验中,用1%的Triton X-100溶解细胞,室温静置10 min,并用PBS混匀制成悬液做10倍倍比稀释,接种到BHI琼脂平板,37 ℃培养24 h后,计算同一稀释度平板的平均CFU值。在进行侵袭试验时,每孔加入含庆大霉素 (100 mg· L-1)和万古霉素(10 mg·L-1)的RPMI-1640培养液1 mL,继续孵育1.5 h,再用D-HanK’s液洗5次,随后的侵袭率计算同黏附试验。
1.4.3 多抗血清和IgG对细胞黏附和侵袭的影响 多抗血清和IgG对细胞黏附和侵袭的影响基本同细菌的黏附、侵袭试验。按不同浓度加入多抗血清和IgG进行优化,然后用优化的等体积的多抗血清和IgG与细菌菌株在37 ℃下孵育1 h,将细菌与血清或IgG混合液接种于24孔细胞板,其他条件和操作同1.4.2细菌的黏附和侵袭细胞试验。
2 结果与分析
2.1 多克隆抗体纯化结果
粗提取并纯化的兔多克隆抗体EbpA1、EbpA2、EbpA3、EF1092A、EbpC1、EbpC2和阴性,经SDS-PAGE电泳鉴定纯化后的IgG见图1。从图1可以看出,纯化后的IgG有明显的2条带,这是因为β-巯基乙醇可以还原免疫球蛋白重链与轻链间的二硫键,从而使两者分离开;其中,,1条约50 kD为IgG的重链,另1条为轻链,约25 kD,且轻链一般比较弥散。

M:蛋白质分子量标准;1~7分别表示兔抗EbpA1,EbpA2,EbpA3,EF1092A,EbpC1,EbpC2,阴性多克隆抗体IgG纯化结果。M: protein molecular weight standard; 1 to 7 indicate rabbit anti-EbpA1, EbpA2, EbpA3, EF1092A, EbpC1, EbpC2, negative polyclonal antibody IgG purification results. A:粗提取的IgG条带。B:纯化后的IgG条带。A:Crude extracted IgG band. B:Purified IgG band.图1 粗提、纯化IgG SDS-PAGE鉴定Fig.1 The SDS-PAGE identification of purificated IgG
2.2 多抗血清及IgG对粪肠球菌生物被膜形成的影响比较
粪肠球菌菌毛亚单位蛋白EbpA1、EbpA2、EbpA3、EF1092A、EbpC1和EbpC2多抗血清的不同效价均对粪肠球菌的生物被膜形成有阻断作用,其最佳效价分别为:1∶8、1∶8、1∶16、1∶64、1∶16和1∶64。以最佳效价菌毛亚单位蛋白多抗血清与粪肠球菌N41和N9菌株作用后对其生物被膜形成的影响如表1所示。菌毛蛋白多抗血清EbpA1和EbpC1对N9菌株生物被膜的形成有阻断作用,但仅有EbpA1与阴性对照相比差异显著(P<0.05)。而对于N41菌株EbpA1,EbpA2,EbpA3,EF1092A,EbpC1和EbpC2对N41菌株生物被膜的形成均有阻断作用,但仅有多抗血清EbpA1与空白对照相比有显著的阻断作用(P<0.05)。

表1 ebp菌毛蛋白多抗血清对粪肠球菌生物被膜形成的影响Table 1 The influence of ebp pili protein antiserum on E.faecalis biofilm formation
注:a代表与阴性对照组相比差异显著;b代表与空白对照相比差异显著。
Note: a represents a significant difference compared to the negative control group; b represents a significant difference compared to the blank control.
粪肠球菌菌毛亚单位蛋白EbpA1、EbpA2、EbpA3、EF1092A、EbpC1、EbpC2多抗IgG浓度不同对N9、N41菌株生物被膜形成的影响有差异。其中,仅有多抗IgG EbpA1、EbpA3和EbpC1对粪肠球菌生物被膜的形成有阻断作用。EbpA1和EbpC1菌毛亚单位蛋白多抗IgG均在质量浓度为0.375 mg·mL-1时对细菌生物被膜形成的阻断作用最强,而EbpA3菌毛蛋白多抗IgG在质量浓度为0.75 mg·mL-1时对细菌生物被膜形成的阻断作用最强,如表2所示。除了EbpA3和EbpC1对N9菌株生物被膜形成与阴性对照相比有显著差异外(P<0.05),EbpA1对N9、N41菌株和EbpA3、EbpC1对N41菌株生物被膜形成与阴性和空白对照相比均有显著阻断作用(P<0.05)。
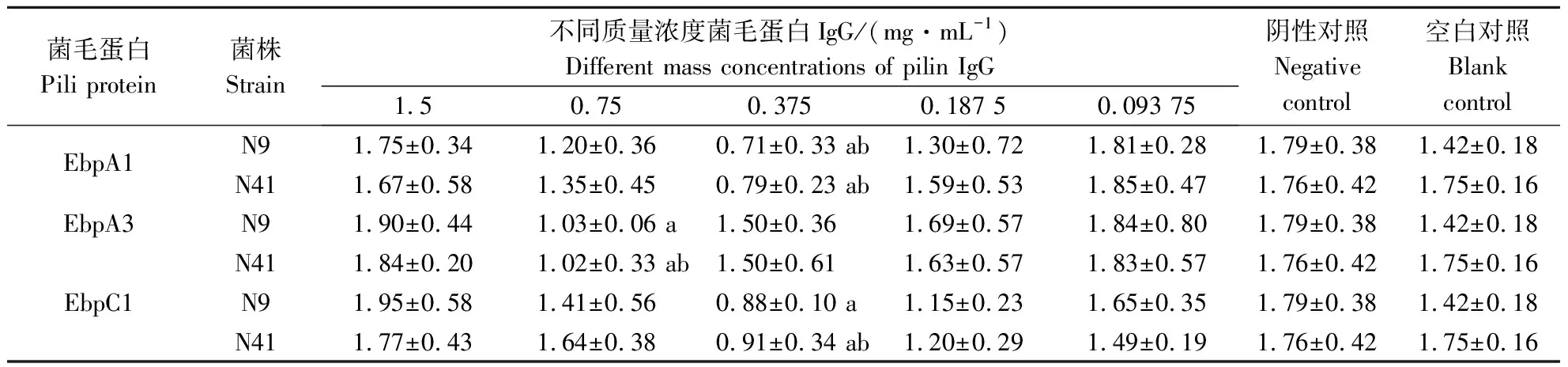
表2 不同质量浓度菌毛蛋白多抗IgG对粪肠球菌生物被膜形成的影响
注:a代表与阴性对照组相比差异显著;b代表与空白对照相比差异显著。
Note: a represents a significant difference compared to the negative control group; b represents a significant difference compared to the blank control.
2.3 兔多抗对细菌黏附、侵袭细胞的影响
2.3.1 最佳多抗血清效价及纯化IgG的浓度优化 通过细菌黏附阻断预试验发现EbpA1、EbpA2、EbpA3、EF1092A、EbpC1和EbpC2多抗血清阻断细菌黏附细胞的最佳血清效价分别是1∶8、1∶8、1∶16、1∶8、1∶32、1∶64;IgG最佳质量浓度分别为0.75、0.75、1.5、0.75、0.75、1.5 mg·mL-1。
2.3.2 Ebp菌毛蛋白多克隆血清对细菌黏附、侵袭细胞的影响 多抗血清以最佳效价与细菌作用1 h,其混合物黏附细胞1.5 h后,细菌黏附细胞的结果如图2。6个多抗血清都对N9、 N41菌株黏附细胞有阻断作用,但EbpA1和EbpC1对N9、N41菌株的黏附阻断作用最强(P<0.05);EbpA3和EbpC2虽对N9菌株黏附有明显的阻断作用,但在对N41的黏附阻断作用中,仅与空白对照相比有显著差异(P<0.05)。另外,在多抗血清作用下,N41菌株的黏附率更低,但在阴性血清对照中,两菌株则出现了明显的差异,N9菌株阴性对照黏附作用更强,而N41菌株则较低,甚至低于EF1092A。
多抗血清对细菌侵袭细胞阻断程度基本同黏附的阻断程度,结果如图3。6个多抗血清对N9、N41菌株侵袭细胞均具有一定的阻断作用,但EbpA1和EbpC1对N9、N41菌株的侵袭阻断作用更强,且与阴性对照和空白对照相比差异极显著(P<0.01);其他各多抗血清也具有明显的侵袭阻断作用(P<0.05,N41菌株的阴性对照除外)。从两菌株的侵袭率分析,N41菌株较N9菌株更低;但在阴性对照中,同样出现了黏附阻断作用现象,即N9菌株的阴性对照侵袭率最高,而N41菌株则低于EF1092A。

图2 菌毛蛋白多抗血清对细菌黏附IPEC-J2细胞的影响Fig.2 The pili protein polyclonal antisera effects on bacterial adhesion IPEC-J2 cells
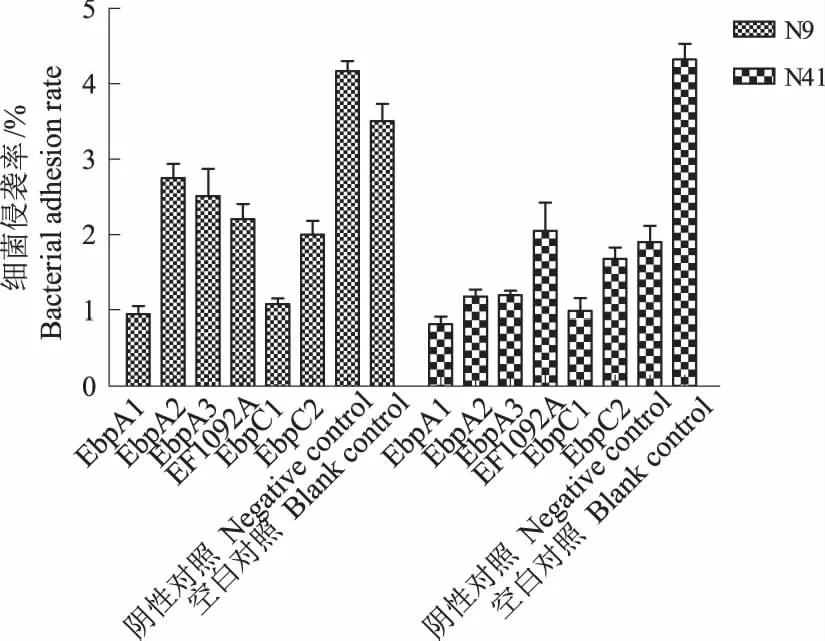
图3 菌毛蛋白多抗血清对细菌侵袭IPEC-J2细胞的影响Fig.3 The pili protein polyclonal antisera effects on bacterial invasion IPEC-J2 cells
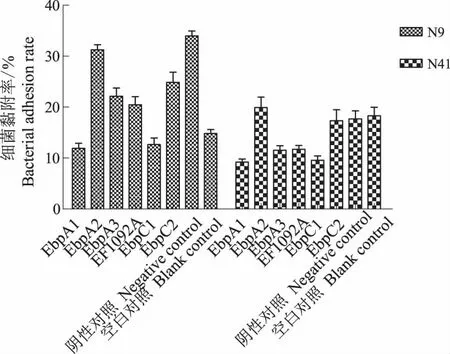
图4 菌毛蛋白多抗IgG对细菌黏附IPEC-J2细胞的影响Fig.4 The pili protein t polyclonal IgG effects on bacterial adhesion IPEC-J2 cells

图5 菌毛蛋白多抗IgG对细菌侵袭IPEC-J2细胞的影响Fig.5 The pili protein t polyclonal IgG effects on bacterial invasion IPEC-J2 cells
2.3.3 Ebp菌毛蛋白多克隆IgG对细菌黏附、侵袭细胞的影响 多抗IgG以最佳浓度与细菌作用1 h,其混合物黏附细胞1.5 h后,细菌黏附细胞的结果如图4。由图4可见,EbpA1和EbpC1多抗IgG对N41菌株和N9菌株均具有黏附阻断作用,但对N41菌株(P<0.01)明显强于N9菌株(P<0.05)有黏附阻断作用;其余4种多抗IgG则作用不同, EbpA2、EbpA3、EF1092A和EbpC2对N9菌株与阴性对照相比有一定的阻断作用,但与空白对照相比,尤其是EbpA2却具有促进黏附作用(P<0.01);EbpA3、EF1092A对N41菌株具有明显的阻断作用,而EbpA2、EbpC2则具有促进黏附作用。
多抗IgG对细菌侵袭细胞作用基本同黏附作用,结果如图5所示。多抗IgG EbpA1、EbpC1的侵袭阻断作用对N41菌株(P<0.01)强于对N9菌株(P<0.05);多抗IgG EbpA3、EF1092A和EbpC2对N9菌株与阴性相比有一定的阻断作用,而多抗IgG EbpA3、EF1092A对N41菌株则有一定的抑制侵袭作用(P<0.05)。
3 讨论
Ebp菌毛是粪肠球菌的表面菌毛,在肠球菌中尤其在粪肠球菌和临床菌株中广泛分布且具有较好的保守性[7],同时也在人的心内膜炎、尿道感染和生物膜形成中发挥重要作用并具有良好的免疫原性[10,13-15]。在粪肠球菌中还发现了能够介导血小板、胶原和纤维蛋白原的黏附,从而引起心内膜炎的ebp菌毛相关毒力基因,这些基因可以编码菌毛蛋白,调节菌毛生物作用[20]。研究证明,ebp菌毛基因缺失突变株对血小板、胶原和纤维蛋白原的黏附能力均显著降低[10]。因此,进一步研究动物感染中Ebp菌毛的致病作用和免疫原性,将可以为肠球菌尤其是粪肠球菌免疫预防、治疗以及诊断奠定坚实的基础。由于Ebp菌毛蛋白分别由EbpA(1033aa)、EbpB(476aa)和EbpC(625aa)3个较大的蛋白组成[21],故寻求它们的抗原表位建立亚单位蛋白抗原,可以更加有效地研究其致病性和免疫保护作用。
本研究使用6个Ebp菌毛亚单位蛋白抗体来研究动物源粪肠球菌Ebp菌毛蛋白的致病性,仅有EbpA1和EbpC1在细菌的生物膜形成和猪肠上皮细胞IPEC-J2的黏附及侵入具有明显的阻断作用,这一结果与NALLAPAREDDY等[10]和PINKSTON等[22]报道的结果相一致。这不但表明构建的EbpA1和EbpC1的正确性,也说明Ebp菌毛蛋白在人源和猪源的致病作用和可能的免疫治疗作用。
本研究使用粪肠球菌N9和N41两株菌株在体外进行Ebp亚单位蛋白的致病性研究,主要是为了验证Ebp亚单位蛋白抗体在不同的粪肠球菌中作用。由于N9和N41菌株在主要的表面黏附毒力因子的不同(因为N9菌株缺少ace毒力基因),也可以检验不同毒力因子对其黏附的贡献。试验结果表明,EbpA1和EbpC1对N41和N9菌株的黏附均具有阻断作用,但对N41的阻断能力强于N9菌株。Ebp和Ace均为细菌表面主要黏附素,N9菌株由于缺少ace基因,EbpA1和EbpC1应该对N9菌株的黏附阻断作用强于N41菌株,出现这一现象的原因可能是Ace的受体宿主细胞外基质蛋白在完整的上皮细胞不易暴露,从而不能被Ace识别;另一种可能是Ebp表达效率,由于Ebp表达需要双稳定表达模式,不同的粪肠球菌Ebp表达范围在30%~72%之间[23]。
猜你喜欢
中国感染与化疗杂志(2022年3期)2022-06-20
安徽农业科学(2022年9期)2022-05-17
中国典型病例大全(2022年7期)2022-04-22
中国农业大学学报(2022年2期)2022-01-05
昆明医科大学学报(2021年3期)2021-07-22
昆明医科大学学报(2020年12期)2021-01-26
山东畜牧兽医(2020年4期)2020-12-29
中华养生保健(2020年1期)2020-11-16
中国感染控制杂志(2019年11期)2019-12-03
医学研究杂志(2015年12期)2015-06-10
