
p300/p53/Smad3通路参与人心房成纤维细胞衰老相关心房纤维化*
2021-02-05 01:02赖颖瑜高小燕周慧珊王钊煜彭德威邓春玉
中国病理生理杂志 2021年1期
赖颖瑜 , 高小燕 , 周慧珊 , 李 昕 , 王钊煜 ,彭德威 , 饶 芳 △, 邓春玉 ,3△
(1南方医科大学药学院,广东广州510515;2广东省心血管病研究所心内科,广东省人民医院医学研究部,广东省医学科学院,广东广州510080;3华南理工大学医学院,广东广州510006)
衰老是随着寿命增长所发生的一个不可避免的过程,而心房纤颤(简称房颤,atrial fibrillation,AF)是临床上最常见的心律失常之一,也是一种“老年病”,其发病率随年龄增长而增加,但目前治疗效果仍不理想,其发病机制需要深入研究。心房结构重塑是AF 的主要发病机制之一,心房纤维化是其主要特征。研究表明衰老与心房纤维化密切相关,与年龄较大者(>70岁)相比,年龄较小者(<50岁)右心耳纤维化明显减少,年龄是影响纤维化发生的主要因素之一[1]。在快速起搏诱发AF 的动物模型中发现,与成年组相比,老年犬左心房纤维化更严重[2],但衰老相关心房纤维化的具体分子机制尚未完全阐明。
转录辅激活因子p300 是一种细胞内普遍存在的核磷酸蛋白,具有内在的乙酰转移酶活性,在细胞增殖,凋亡和胚胎发育中都发挥关键作用[3]。研究发现,p300参与纤维化,如在特发性肺纤维化患者的肺成纤维细胞中,p300 明显上调,而抑制p300 能减轻纤维化[4-5];p300 还影响细胞衰老,研究显示在微血管内皮细胞中,高糖诱导的沉默信息调节因子1(silent information regulator 1,Sirt1)下调,p300 表达上调,导致细胞快速衰老[6]。我们的前期研究结果也显示,p300 随着体外培养的小鼠心脏成纤维细胞衰老而升高,伴随有纤维化指标的增加[7]。因此,p300可能参与心脏衰老相关纤维化的病理过程。
Smads 是纤维化通路的重要信号分子[8],衰老通路中的重要因子p53 也与纤维化相关[9-10]。研究发现,在人肾小管上皮细胞株HK-2 和角化细胞株Ha-CaT中,转化生长因子β1(transforming growth factor-β 1,TGF-β1)通过调控p53 活性进而促进 p53 与Smads相互作用,随后结合到纤溶酶原激活物抑制因子1(plasminogen activator inhibitor-1,PAI-1)启动子上,导致纤维化[11]。而 p300 可通过乙酰化调控 p53[12]。这些研究提示,p300 可通过调控p53,促进p53-Smad3相互作用进而参与衰老相关纤维化。
因此,本研究采用人心房成纤维细胞(human atrial fibroblasts,HAFs)作为研究对象,通过传代建立细胞衰老模型,比较不同代数的细胞中p300、p53、Smad3 和其他衰老及纤维化相关因子的变化,并通过干预p300 的表达,观察其对p53 和Smad3 等衰老和纤维化指标的影响,探索p300 在衰老相关纤维化中的作用及其可能机制,以期发现治疗衰老相关心房纤维化的新靶点。
材 料 和 方 法
1 患者组织标本
本研究收集患者于心脏体外循环手术中或经胸腔镜AF 外科消融手术中需切除的心耳组织,分离培养原代心房成纤维细胞并进行传代培养。所有患者均签署知情同意书,并获广东省人民医院(广东省医学科学院)伦理委员会批准,批准号为No.GDREC2016128H。有肺炎或者其他感染性疾病的患者不入选。
2 试剂
特级澳洲胎牛血清和0.25% 胰蛋白酶-EDTA(Gibco);成纤维细胞基础培养基(fibroblast basal medium,FBM)(Lonza);p300 小发卡(small-hairpin,sh)RNA 质粒和 p300 过表达质粒 pCMV p300 CHA(吉凯基因);Lipofectamine 3000 转染试剂(Invitrogen);姜黄素(Cayman);蛋白 Marker(Fermentas);4×蛋白上样缓冲液(BIO-RAD);RIPA 裂解液(强)(Beyotime);抗p300 抗体(Millipore);抗I 型胶原蛋白α1 链(collagen type I α1 chain,Col1A1)抗体和抗Ⅲ型胶原蛋白α1 链(collagen type Ⅲ α1 chain,Col3A1)抗体(Abcam);抗基质金属蛋白酶2/9(matrix metalloproteinase-2/9,MMP-2/9),p21 和 p16 抗体(Santa Cruz);抗p53,Smad3,p-Smad3,GAPDH,PAI-1 和 TGF-β 抗体及衰老相关β-半乳糖苷酶(senescence-associated βgalactosidase,SA-β-Gal)染色试剂盒(Cell Signaling Technology);其他生化试剂均为进口分装或国产分析纯。
3 方法
3.1 细胞培养及质粒转染 人心耳组织经PBS 漂洗后去除多余脂肪组织和内外膜,加入几滴FBM 培养基(含10%胎牛血清),细剪成约1 mm ×1 mm ×1 mm 小块,用吸管将组织块均匀涂布于25 cm²培养瓶培养面上,静置约15 min,轻翻培养瓶从侧面加入4 mL FBM 培养基(含10%胎牛血清),培养面朝上于37 ℃,5%CO₂的培养箱中静置2 h,组织块贴牢后,翻转培养瓶,培养面朝下放置使培养基浸过所有组织块,继续培养。每隔5 d换液,待细胞游离、生长至基本融合成片达瓶底80%后,进行消化传代。传代至P3 和P7 的细胞接种于6 孔细胞培养板中,待细胞密度达70%~80%,使用Lipofectamine 3000 转染试剂盒按说明操作进行质粒转染。质粒稀释液:125 μL Op-ti-MEM 培养基稀释 2.5 μg 质粒和 5 μL P3000 试剂,轻轻混匀;脂质体稀释液:125 μL Opti-MEM 培养基稀释 6 μL Lipofectamine 3000 试剂,高速震荡 3 s 混匀。然后将这两份分别含有质粒和脂质体的溶液混合,轻轻混匀,室温孵育15 min,均匀滴加到细胞新鲜更换的1 mL FBM 培养基(含10%胎牛血清)中,轻轻摇匀,于培养箱中继续培养2~3 d。
3.2 SA-β-Gal 染色 β-半乳糖苷酶是衰老细胞的标志物。将细胞接种于6 孔细胞培养板中,待密度达到50%~70%,按试剂盒操作说明进行SA-β-Gal 染色。弃培养基后用PBS 漂洗1 次。加入固定液室温固定15 min,去除固定液后PBS 漂洗2次。按照说明配制染色液,调节pH 值为6,每孔加1 mL 染色液,密封,于37 ℃,无CO2的干燥恒温箱孵育过夜。在显微镜下观察,核周蓝染的即为衰老细胞。
3.3 Western blot实验 细胞弃去培养基,PBS洗3次后加入适量含蛋白酶抑制剂的RIPA 裂解液(强),冰上裂解5 min,用刮刀刮细胞,用移液枪收集细胞混合物至 EP 管中,冰上继续裂解 20 min。4 ℃,12 000 r/min 离心15 min,取上清分装至EP 管,储存于-80 ℃。BCA法测定蛋白浓度。加入4×蛋白上样缓冲液稀释30 μg 蛋白,55 ℃煮 10 min。行 8%~10% SDS-PAGE分离蛋白样品,转移到PVDF 膜上。蛋白面向上用5%脱脂牛奶于室温封闭1 h。TBST 洗3 次,每次5 min。加入稀释好的Ⅰ抗4 ℃孵育过夜,TBST 洗3次,每次5 min。加入稀释好的Ⅱ抗室温孵育1 h,TBST 洗3 次,每次5 min。蛋白面朝上均匀滴加ECL孵育30 s,平铺在托盘上送入机器自动曝光。
3.4 细胞免疫荧光染色 待细胞密度达到70%~80%时,用预温PBS 洗2 次。4%多聚甲醛固定15 min,PBS 浸洗 3 次,每次 10 min。0.2% TritonX-100通透20 min,4%BSA 封闭30 min,去除封闭液直接滴加稀释好的Ⅰ抗放入湿盒,4 ℃孵育过夜。PBS 浸洗3 次,每次10 min。滴加稀释好的荧光Ⅱ抗,湿盒中避光室温孵育45 min。PBS 浸洗3 次,每次10 min。避光滴加封片剂(含DAPI)后在荧光显微镜下观察。
4 统计学处理
采用统计软件SPSS 20.0 分析处理数据。数据以均数±标准误(mean±SEM)表示。两组间比较用t检验。多组间比较用单因素方差分析,各组均数的两两比较采用 LSD-t或 SNK-q检验,以P<0.05 为差异有统计学意义。
结 果
1 不同代数人心房成纤维细胞SA-β-Gal 染色结果以及p300、p53 和Smad3 等衰老和纤维化相关蛋白水平的变化
体外培养的HAFs 连续传代至P11 代,行Western blot 检测不同代数(P3、P7 和P11)细胞的 p300、p53和Smad3等衰老和纤维化相关因子的蛋白水平,结果如图1A、1B所示,随着细胞代数增加,p300和衰老相关因子p53表达水平逐渐升高,以P11代细胞最高(P<0.05;P<0.01)。与P3 代细胞相比,P7 代细胞的p21 和p16 表达水平都明显升高(P<0.01),且p16在P11 代达最高水平(P<0.01)。Smad3 是公认的促纤维化因子,随着细胞传代,总Smad3 有下降趋势,但无统计学意义,p-Smad3 的蛋白水平在P7 和P11代细胞中均明显增高(P<0.05),其他纤维化指标MMP-2/9 和PAI-1 表达均随着代数逐渐增加,以P11代细胞最高(P<0.01),但Col1A1和Col3A1的表达在各组间无明显差异,TGF-β 在P11 代细胞中表达较P3 和P7 有所下降,但无统计学差异。另外,分别取P3、P7和P11代细胞进行SA-β-Gal染色,观察细胞衰老情况,结果如图1C 所示,随着传代代数增加,核周蓝染的衰老细胞逐渐增加(P<0.01)。由此可见,随着 HAFs 衰老,p300 和 p53 表达增加,Smad3 被激活,一些纤维化相关因子表达增加。
2 姜黄素处理高代数人心房成纤维细胞对衰老和纤维化相关蛋白水平的影响
为了进一步探索p300 在HAFs 衰老以及衰老相关的纤维化中的作用,我们给予不同浓度(6、9 和12 μmol/L)的 p300 抑制剂姜黄素处理高代数(P7)HAFs,使用 Western blot 检测细胞中 p300、p53 和Smad3 以及其他衰老和纤维化相关蛋白水平的变化。结果如图2 所示,不同浓度姜黄素处理后,P7 代细胞的p300 水平呈现浓度依赖性下降(P<0.05),以12 μmol/L 浓度处理组达最低水平(P<0.01);衰老相关指标p53、p21 及p16 表达水平也呈逐渐下降趋势,Smad3 及p-Smad3 的蛋白水平明显下降,以高浓度处理组下降最为明显(P<0.05);其他纤维化指标Col1A1、MMP-2、TGF-β 和PAI-1 在高浓度姜黄素(12 μmol/L)处理后,亦明显下降(P<0.05;P<0.01);而Col3A1 和MMP-9 的蛋白水平也有下降趋势,但差异无统计学显著性。因此,p300 抑制剂姜黄素处理抑制高代数HAFs的p300,可明显降低衰老和部分纤维化因子的蛋白水平,提示p300 在HAFs 衰老和纤维化中发挥着重要作用。
3 敲减p300 的表达对高代数人心房成纤维细胞衰老和纤维化相关蛋白水平的影响
为了确定p300 在HAFs 衰老相关的纤维化中发挥的关键作用,我们对高代数(P7)细胞进行p300 shRNA 质粒转染敲减p300的表达,观察衰老和纤维化相关因子的变化,结果如图3 所示。质粒转染后p300 的表达水平下降(P<0.05),伴随p53、Smad3 及p-Smad3 的蛋白水平显著降低(P<0.01),同时其他纤维化相关指标如Col1A1、Col3A1 和MMP-2 的蛋白水平也降低(P<0.05;P<0.05),MMP-9、TGF-β 和PAI-1 水平也有下降趋势,但差异无统计学显著性。由此说明,敲减HAFs 的p300表达可以抑制衰老及纤维化相关因子的分泌。
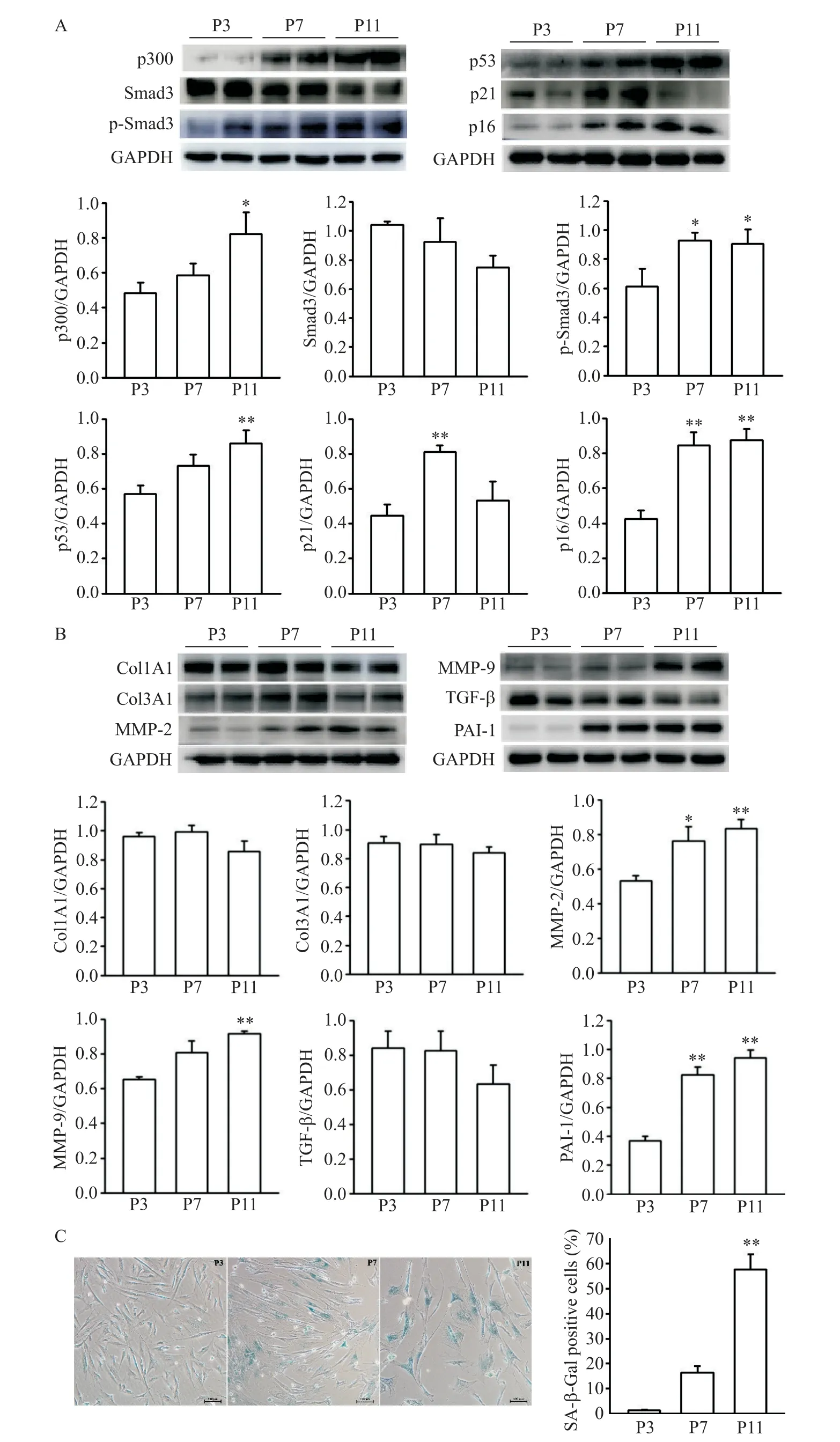
Figure 1.The results of SA-β-Gal staining and the protein expression levels of p300,senescence and fibrosis associated proteins in human atrial fibroblasts of different passages.A:the protein levels of p300,Smad3/p-Smad3,p53/p21 and p16;B:the protein expression of Col1A1/3A1,MMP-2/9,TGF-β and PAI-1;C:the images of SA-β-Gal staining(×100)and SA-β-Gal positive rate of HAFs at passage 3,passage 7 and passage 11,respectively.Mean±SEM. n=6.*P<0.05,**P<0.01 vs P3 group.图1 不同代数人心房成纤维细胞SA-β-Gal染色结果及p300、衰老和纤维化相关蛋白的表达水平

Figure 2.Effect of curcumin on the expression of p300,senescence and fibrosis associated proteins in human atrial fibroblasts of passage 7.A:the protein levels of p300,Smad3/p-Smad3,p53/p21 and p16 before and after treated with curcumin of different concentrations(6,9 and 12 μmol/L);B:the protein expression of Col1A1/3A1,MMP-2/9,TGF-β and PAI-1 before and after treated with curcumin.Mean±SEM. n=4.*P<0.05,**P<0.01 vs DMSO group.图2 姜黄素处理对人心房成纤维细胞p300及衰老和纤维化相关蛋白表达的影响
4 人心房成纤维细胞过表达p300 对衰老和纤维化相关蛋白水平的影响
对低代数(P3)HAFs进行质粒转染过表达p300,观察衰老和纤维化相关指标的变化,结果如图4 所示,转染后p300 的表达显著增加(P<0.05),p53 和Smad3 的水平也明显升高(P<0.05),纤维化指标MMP-2的水平也随p300而增加(P<0.05),而Col1A1无明显变化。此部分研究结果显示,低代数的HAFs过表达p300,可促进衰老和纤维化相关因子的分泌。
讨 论
以上研究结果表明,随着HAFs 衰老,p300 及其他衰老和纤维化相关因子水平升高;姜黄素处理或p300 shRNA质粒转染敲减p300的表达可降低p53和Smad3 的表达,抑制细胞衰老和纤维化相关因子分泌;而过表达p300 则可促进细胞衰老和纤维化相关因子分泌,说明p300 在HAFs 衰老相关纤维化中起重要作用,可能通过调控p53/Smad3通路参与其相关病理机制。
心脏纤维化的发生发展是一个复杂的过程,相关病理因素包括,遗传因素,代谢障碍,肥胖和高血压等。近年来研究表明,除却病理因素,衰老亦会导致心肌细胞外基质的改变,如心脏成纤维细胞的增殖和胶原蛋白含量增加,交联增强,导致心脏顺应性下降[13]。AF 的主要病理机制包括心房肌细胞的电重塑和结构重塑,心房纤维化是结构重塑的主要表现。而进行性心房纤维化亦与心脏衰老相关[14],与年幼大鼠相比,老年大鼠的左房纤维化增加,且具有更高的 AF 诱发性[15]。因此,AF 的发生率随年龄的增长而增加,与增龄相关的心房纤维化有关,心房组织呈现年龄依赖的间质纤维化,心肌细胞间的电通讯受阻,异位或折返活动的可能性增加,使老年人容易发生AF[15]。目前虽然衰老相关的心房纤维化研究已获关注,但其相关机制尚未得到很好阐明。因此,本研究以HAFs 为研究对象通过传代建立复制性细胞衰老模型,探究其相关机制。
转录辅激活因子p300 是体内重要的表观遗传调控分子,具有内的在乙酰转移酶活性,可通过乙酰化组蛋白和非组蛋白参与调控多种基因的表达[3,16]。研究表明,p300 可调控胶原蛋白表达,促进纤维化,如在糖尿病大鼠心肌肥厚模型中,p300 通过调控 Smad2 乙酰化,增加 TGF-β 活性,促进胶原合成,导致心肌纤维化和肥厚[17];p300 HAT 特异性抑制剂—L002 在体内和体外均能有效抑制纤维化反应,如减弱体外培养的人心脏成纤维细胞分化,增殖,迁移及胶原合成能力,逆转高血压诱导的小鼠心肌纤维化和肥大[18-19]。虽然,直接关注p300 在细胞衰老中作用的研究不多,但在人原代包皮成纤维细胞中发现,p300 对原癌基因Ras诱导的细胞早衰至关重要,抑制p300 活性可阻止p53 活化,有助于肿瘤转移中的衰老逃逸[20]。而在人脐带来源的间充质基质细胞中,敲减p300的表达或抑制其乙酰转移酶活性将诱导细胞早衰并降低增殖潜能,提示p300 通过激活p53/p21 信号途径,在诱导细胞衰老过程中起重要作用[21]。本研究发现,p300 的表达水平随着HAFs 传代代数的增加而升高,同时伴随其他衰老和纤维化相关因子增加。姜黄素处理或p300 shRNA 质粒转染敲减p300的表达可抑制衰老信号通路和纤维化因子分泌,而过表达p300 则相反。由此证实p300 在HAFs 衰老相关纤维化中发挥关键作用。
本研究还对p300 参与衰老相关纤维化的分子机制进行初步探索。p53 是一种序列特异性DNA 结合蛋白,是调控细胞增殖,凋亡和衰老的重要转录激活子[22]。乙酰化是调控p53 功能的关键共价修饰,乙酰转移酶p300 介导的p53 乙酰化不仅可增加其蛋白稳定性和转录活性,还可以增强其反应性基因,如细胞周期依赖性激酶抑制剂p21等的启动子激活促进细胞周期停滞或衰老[23],这可能是p300 参与细胞衰老的重要途径。有研究发现,高糖刺激可通过Sirt1/p300/p53/p21 途径诱导持续的内皮细胞衰老[24]。同时,p53与Smad3都是纤维化反应的效应因子[25],在 TGF-β1 刺激肾纤维化中 p53 与 Smad3 有明显协同作用,p53 通过与受体激活的Smads 结合而充当TGF-β1 诱导的信号传导途径中的辅助因子和TGF-β1 纤维化反应基因的转录共调节因子[26],促进肾纤维化疾病进展。本研究中也发现,p300 随着细胞传代而增加,且在过表达p300 时,p53 和Smad3 的活性随之增加,并伴随纤维化因子增加;降低p300水平后,p53 和Smad3 的水平也下降,同时纤维化因子分泌减少。因此,p300可能通过调控p53和Smad3参与衰老相关纤维化。

Figure 3.The protein levels of senescence and fibrosis associated proteins in human atrial fibroblasts of passage 7 after p300 shRNA transfection.The protein levels of p300,p53,Smad3/p-Smad3,Col1A1/3A1,MMP-2/9,TGF-β and PAI-1 in passage 7 HAFs after transfected with p300 shRNA plasmid were determined by Western blot.Mean±SEM. n=4.*P<0.05,**P<0.01 vs control group.图3 高代数(P7)人心房成纤维细胞敲减p300的表达对衰老和纤维化相关蛋白表达的影响

Figure 4.The protein levels of senescence and fibrosis associated proteins in human atrial fibroblasts of passage 3 after p300 over-expression.The protein levels of p300,p53,Smad3,Col1A1 and MMP-2 in passage 3 HAFs after transfected with p300 over-expression plasmid were determined by Western blot.Mean±SEM.n=4.*P<0.05 vs vector control group.图4 低代数(P3)人心房成纤维细胞过表达p300对衰老和纤维化相关因子表达的影响
此外,本研究具有以下局限性:虽然我们发现敲减p300的表达可明显降低纤维化相关因子,如Col 1A1/3A1 和 MMP-2 的 表 达 ,同 时 p53 和 Smad3/p-Smad3 的水平亦明显降低,而p300 过表达后p53 和Smad3 水平明显升高,纤维化指标MMP-2 的表达亦明显增加,但对Col 1A1 的表达无明显影响。其原因可能是因为本研究中的HAFs来源于人体心房组织,均为接受心脏手术的患者,其心房可能均有一定程度的病理改变,在这个基础上分离的心房成纤维细胞可能已有一定程度的纤维化,导致其胶原的表达已经比较高,故过表达p300 不能使胶原合成进一步增加。
综上所述,本研究初步证实p300 可能通过激活p53/Smad3 通路参与人心房成纤维细胞衰老相关心房纤维化,最终导致AF的发生和发展。
猜你喜欢
昆明医科大学学报(2021年8期)2021-08-13
云南医药(2021年3期)2021-07-21
河北理科教学研究(2021年4期)2021-04-19
数学年刊A辑(中文版)(2021年4期)2021-02-12
科学(2020年1期)2020-08-24
Medical Data Mining(2019年2期)2019-07-16
中成药(2018年2期)2018-05-09
中成药(2018年3期)2018-05-07
中国现代医学杂志(2015年26期)2015-12-23
应用数学与计算数学学报(2015年1期)2015-07-20
