
杜鹃花品种间杂交F1代主要形态性状变异1)
2021-05-26 07:56苏鸣夏溪张春英奉树成倪穗
东北林业大学学报 2021年5期
苏鸣 夏溪 张春英 奉树成 倪穗
(宁波大学,宁波,315800)(上海植物园)(宁波大学)
杜鹃花是杜鹃花科(Ericaceae)杜鹃花属(Rhododendron)植物的总称,是著名观赏植物类群、我国十大名花之一[1]。杜鹃花资源丰富,观赏价值高,是城市绿化、庭院美化的重要植物类群。据统计,全世界杜鹃花属植物约967种,其中562种原产中国[2],我国西南山区是杜鹃花资源最重要的发源地和集聚中心[3]。根据英国皇家园艺学会(RHS)统计,全世界登录的杜鹃花品种已经超过3万种,但是绝大多数杜鹃花品种却源自欧美和日本[4-6]。虽然近年来我国陆续有杜鹃花新品种推出[7-9],但是截至2020年国家林业和草原局植物新品种保护办公室审定的杜鹃花品种数量刚刚突破百种。
随着我国经济水平的提高、生态文明和美丽中国建设的推进,人们对观赏植物品种多样性的需求不断增加。我国长江流域及以南地区园林绿地中应用最多的杜鹃花是锦绣杜鹃(Rhododendronpulchrum)类品种[10],抗性强,栽培容易,在绿化市场备受青睐。由于国内露地栽培杜鹃花抗逆好的品种少,可用于绿地栽培的杜鹃花品种十分单调,无法满足绿地品种多样性的需求,因此如何快速培育特色各异的抗逆品种成为杜鹃花产业发展亟待解决的问题。目前,杂交育种是国内外杜鹃花新品种培育中最有效、最常用的方法之一[10]。杂交后代主要性状变异规律的预测是亲本选配和提高育种效率的关键。常见观赏植物如菊花(Chrysanthemumspp.)[12]、紫薇(Lagerstroemiaspp.)[13]、月季(Rosaspp.)[14]、百合(Liliumspp.)[15]等杂交子代性状变异规律均有大量研究,关于杜鹃花杂交后代性状变异分析的文献却很少,仅见刘晓青等[16]对23个杂交组合F1代主要观赏性状的遗传表现进行了简单分析,马绍宇等[17]对高山杜鹃1个杂交群体F1代主要观赏性状的遗传分析,de Keyser et al.[18]对4个常绿杜鹃杂交组合F1代的花及叶片特征进行了QTL定位图像分析。因此,本研究选择了抗逆性较好的杜鹃花品种作为亲本进行杂交,并获得杂交子代开花植株。对3组杜鹃花品种间杂交F1代主要形态性状进行观测,分析杂交后代主要形态性状变异特点及规律,为杜鹃花新品种选育和杂交亲本的合理选配提供参考。
1 材料与方法
1.1 试验材料
杜鹃花品种‘紫鹤’(Rhododendronpulcherum‘Zihe’)与‘红苹果’(Rh. ‘Red Apple’)、‘白鹤’(Rh. pulcherum ‘Baihe’)与‘粉红泡泡’(Rh. ‘Pink Bubble’)、‘紫波’(Rh. ‘Zibo’)与‘麒麟’(Rh. ‘Kirin’)3组杂交组合的亲本及稳定开花的杂交F1子代植株。杂交于2011年在浙江省宁波市北仑万景杜鹃良种园进行,杂交子代在2015年3月开花。杂交亲本的主要形态特征见图1和表1。

a.‘紫鹤’;b.‘红苹果’;c.‘白鹤’;d.‘粉红泡泡’;e.‘紫波’;f.‘麒麟’。
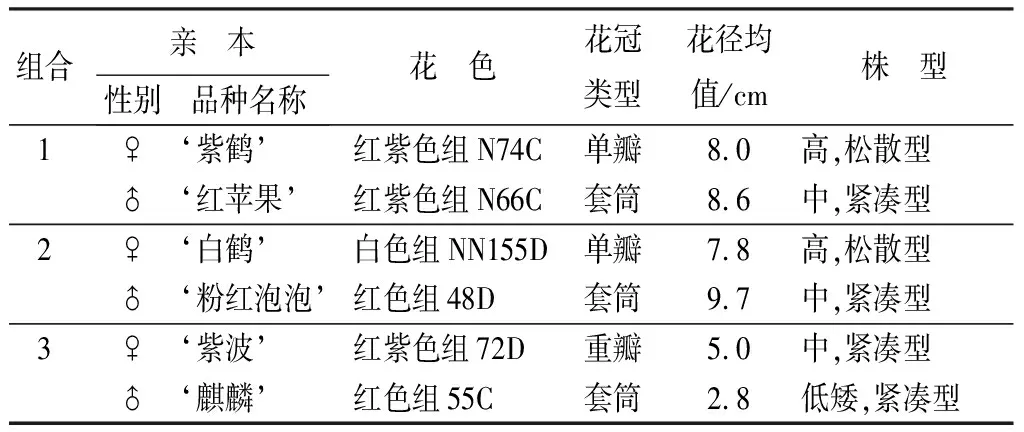
表1 杜鹃品种间杂交亲本的主要形态特征
1.2 试验方法
表型性状的选取:2018—2019年对3组杂交组合的亲本及部分F1子代的表型性状进行调查和统计。根据中华人民共和国林业行业标准LY/T 1852—2009《植物新品种特异性、一致性、稳定性测试指南 杜鹃花属 映山红亚属和羊踯躅亚属》中相关内容选取5个数量性状,4个质量性状,共计9个性状进行统计,其中花性状6个,叶性状3个(见表2)。
数据采集方法:数量性状采用直尺直接测量法,并重复3次取平均值。统计花相关数据时,选择同一植株不同位置的3朵花进行测量记录;统计叶相关数据时,选择同一植株上3片从上向下数第3层成熟无病害叶片进行测量记录。质量性状采取观察分类法,其中花色以《英国皇家园艺协会的色谱》为标准,分为白色、紫粉、红粉、红色、紫红、紫色、复色共7类,花瓣形状和花冠类型以中华人民共和国林业行业标准LY/T 1852—2009《植物新品种特异性、一致性、稳定性测试指南 杜鹃花属 映山红亚属和羊踯躅亚属》进行分类和数量化处理,数据数量化编码见表2。

表2 杜鹃花品种表型性状分类及编码
1.3 数据分析
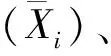
中亲值:MPS=(P1+P2)/2,P1、P2为亲本各表型性状实际测量值。
2 结果与分析
2.1 花性状变异
2.1.1 花色
3个杂交组合的花色变异系数平均值为38.65%,达到较大变异水平,不同组合之间差异大(见表3)。其中‘紫波’与‘麒麟’杂交子代的花色变异系数最大,为52.71%,双亲花色分别是紫红和红粉,子代花色分离广泛,出现了白色、紫粉、红粉、红色、紫红、复色(整体白色,瓣尖淡粉色)共计6种花色,所占比例分别是6.50%、25.80%、29.00%、12.90%、6.50%、19.40%,与父本花色相近的红粉色花数量最多。‘白鹤’与‘粉红泡泡’杂交子代的花色变异系数为36.06%,属于较大变异。双亲花色为白色和红粉,子代花色为白色、紫粉和红粉。子代花色中占比最多的是和父本花色相同的红粉,为48.40%,新花色“紫粉”占子代总数的29.00%。‘紫鹤’与‘红苹果’杂交子代的花色变异系数为27.17%,属于中等变异,亲本花色都是紫红色,子代花色为紫粉、紫红和紫色,其中以与亲本相同的紫红最多,占所有子代总数的66.70%。
2.1.2 花冠裂片内纹饰形态
花冠裂片内纹饰形态是杜鹃花的重要观赏性状之一,3个杂交组合变异程度差异较大(见表4)。其中,‘紫鹤’与‘红苹果’和‘白鹤’与‘粉红泡泡’2个组合的亲本花冠裂片内均有斑点,但是只有‘白鹤’与‘粉红泡泡’杂交组合出现少量无斑点子代,占总数的6.45%,变异系数为12.90%。父本‘麒麟’花冠裂片内无斑点,母本‘紫波’花冠裂片内有斑点,子代无斑点和有斑点的植株分别占总数的38.71%和61.29%,多数子代具有与母本性状类似的花冠裂片有斑点的特性。
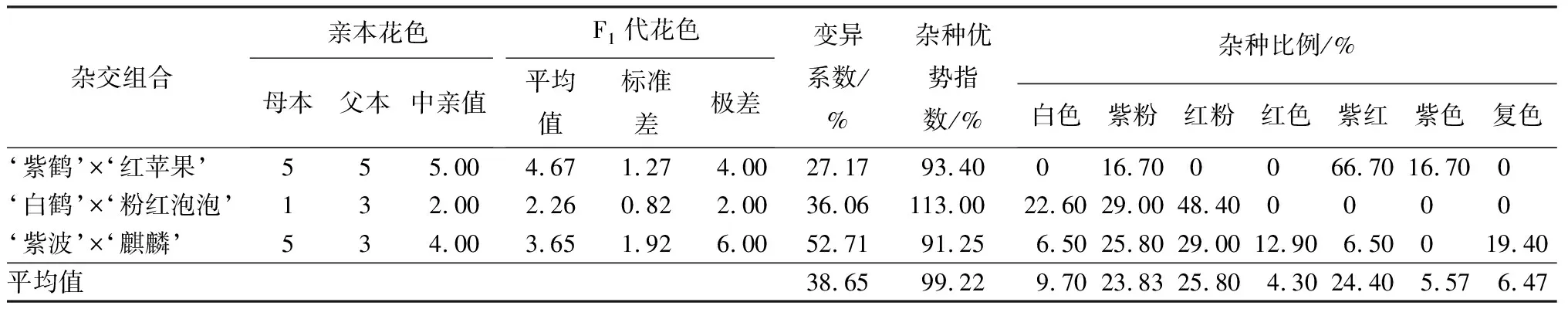
表3 杜鹃花品种间杂交F1代花色变异情况
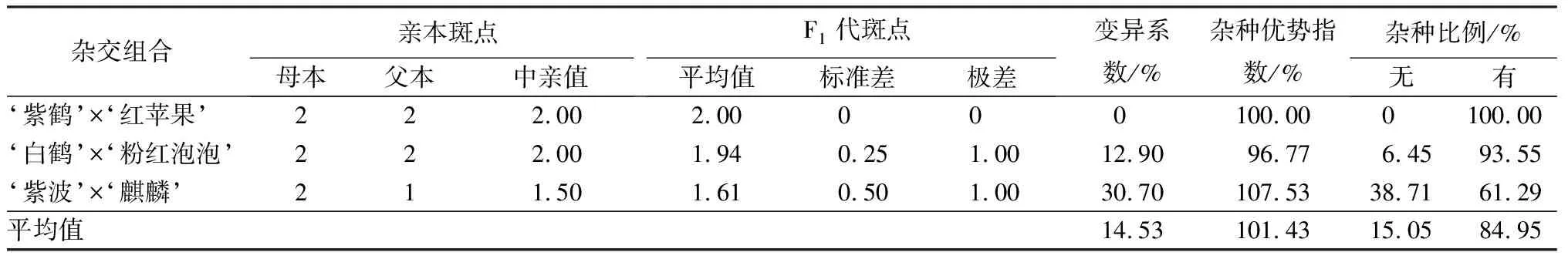
表4 杜鹃花品种间杂交F1代花冠裂片内斑点变异情况
2.1.3 花冠类型
3个杂交组合的花冠类型变异系数差别较大(见表5)。‘紫鹤’与‘红苹果’和‘白鹤’与‘粉红泡泡’2个组合的母本均为单瓣,父本均为套筒,但子代花冠类型的分离情况完全不同。‘紫鹤’与‘红苹果’子代全部为与父本相同的套筒花,变异系数为0,而‘白鹤’与‘粉红泡泡’组合的变异系数为35.70%,子代中单瓣和套筒占总数的比例分别是61.29%和38.71%,多数子代表现为与母本相同的单瓣花冠类型。‘紫波’与‘麒麟’组合的母本花冠类型为重瓣,父本为套筒,子代花冠类型中单瓣、套筒、半重瓣套筒和重瓣的比例分别是16.13%、35.48%、38.71%、9.68%,其中单瓣和半重瓣套筒是与亲本不同的花冠类型,共占子代总数的54.84%,变异系数为36.78%,属于较大变异。
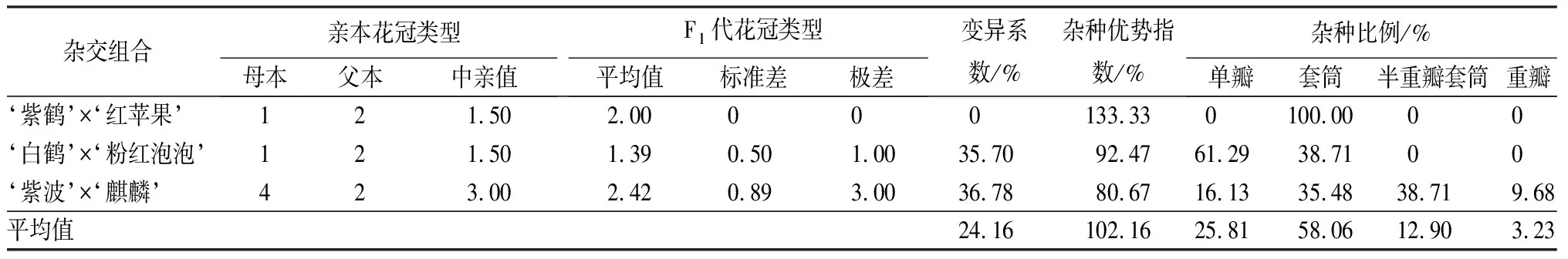
表5 杜鹃花品种间杂交F1代花冠类型变异情况
2.1.4 花瓣形状
3个杂交组合亲本均为椭圆形花瓣,子代中椭圆形花瓣的数量占比最大,是77.42%。但是不同杂交组合的子代变异系数差别较大(见表6)。其中,‘紫鹤’与‘红苹果’组合的变异系数为0,子代均为椭圆形花瓣;‘白鹤’与‘粉红泡泡’组合的变异系数为16.01%,属于中等变异,并出现了新的阔椭圆形花瓣性状,占总数的12.90%;‘紫波’与‘麒麟’组合子代性状中窄椭圆形、椭圆形、阔椭圆形和卵形分别占总数的3.23%、45.16%、29.03%和22.58%,新出现的花瓣性状占总数的54.84%,变异系数为31.73%,属于较大变异。
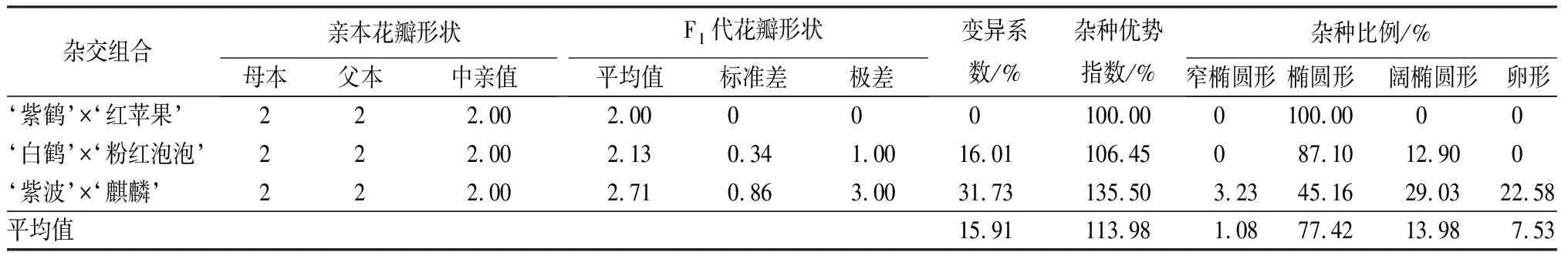
表6 杜鹃花品种间杂交F1代花瓣形状变异情况
2.1.5 花径
3个杂交组合花径的变异系数较小且相近,为9.58%~11.41%,但是杂种优势度各不相同(见表7)。‘紫波’与‘麒麟’杂交子代的杂种优势指数最大,为125.90%,呈现明显的超亲本现象,高亲比例达35.48%,没有出现低于双亲的个体;其次‘紫鹤’与‘红苹果’组合杂种优势指数为104.98%,超亲本子代占60.00%,,其中高亲比例达46.67%;‘白鹤’与‘粉红泡泡’组合杂种优势指数为92.98%,子代花径介于双亲的为61.29%,低于双亲的为35.48%,高于双亲为3.23%。从子代花径分布情况可见,不同杂交组合分布各不相同,超亲子代较多,但是没有明显偏向于父本或母本。
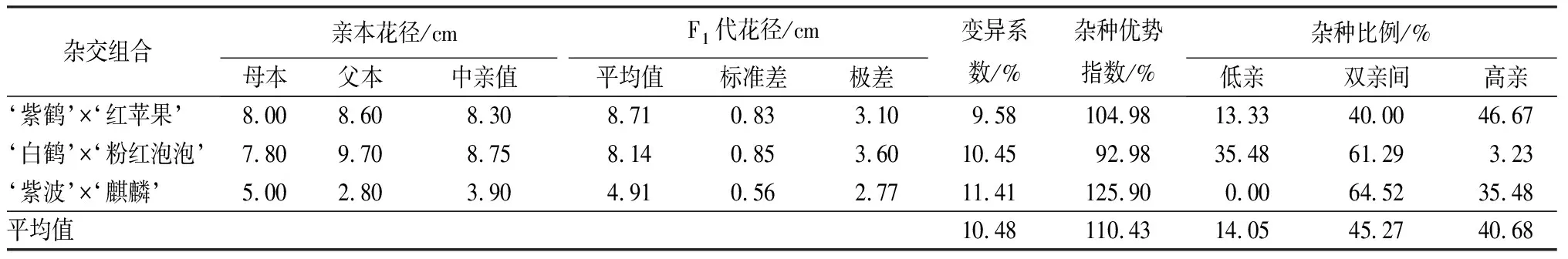
表7 杜鹃花品种间杂交F1代花径变异情况
2.1.6 花梗长度
3个杂交组合花梗长度的变异系数差异不大,平均变异系数为17.59%,达到了中等变异水平(见表8)。但是3个组合的杂种优势指数差异较大,‘紫鹤’与‘红苹果’组合的杂种优势指数最大,为140.14%,组合中高亲子代占子代总数的86.67%;‘紫波’与‘麒麟’组合的杂种优势指数最小,为86.15%,组合中96.77%的子代介于亲本之间;‘白鹤’与‘粉红泡泡’组合杂种优势指数为129.89%,但是这个组合子代高亲比例最高,为90.91%。从子代花梗长度分布情况可见,亲本花梗长度差异越小,子代高亲数量越多;亲本花梗长度差异较大时,子代多介于亲本之间。
2.2 叶部性状的遗传
叶部性状包括叶长、叶宽和长宽比3个性状(见表9和表10)。‘紫鹤’与‘红苹果’、‘紫波’与‘麒麟’2组杂交组合的子代叶片生长情况类似,表现为叶长、叶宽的杂种优势指数呈下降趋势,长宽比的杂种优势指数呈上升趋势,表明这2个杂交组合子代的叶片整体变小,变得更加窄长。杂交组合‘白鹤’与‘粉红泡泡’与其他2组相反,叶长和叶宽的杂种优势指数呈上升趋势,长宽比呈下降趋势,表明这组杂交组合的子代叶片整体变大,变得更加宽短。从叶长、叶宽和叶长宽比子代分布情况看,叶部性状没有偏向于亲本中的任何一个。
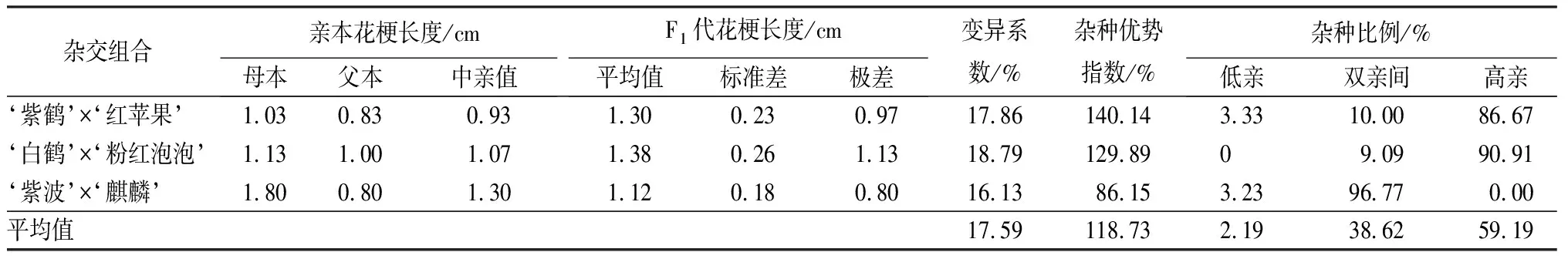
表8 杜鹃花品种间杂交F1代花梗长度变异情况
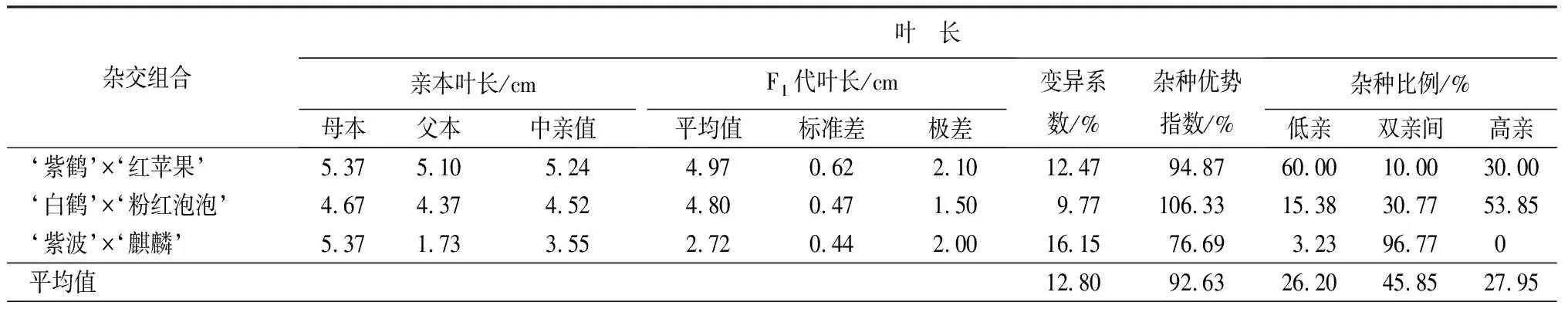
表9 杜鹃花品种间杂交F1代叶片性状变异情况
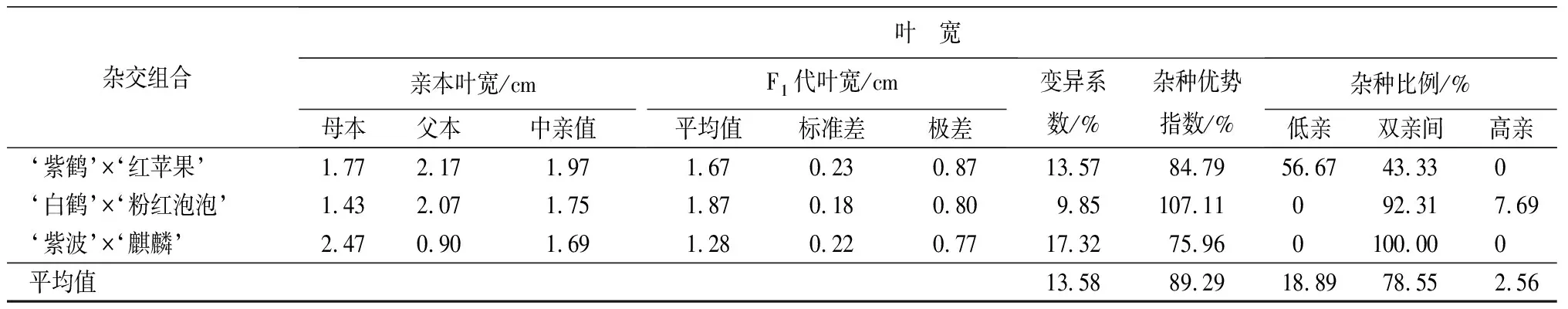
杂交组合叶 宽亲本叶宽/cm母本父本中亲值F1代叶宽/cm平均值标准差极差变异系数/%杂种优势指数/%杂种比例/%低亲双亲间高亲‘紫鹤’ב红苹果’1.772.171.971.670.230.8713.5784.7956.6743.330 ‘白鹤’ב粉红泡泡’1.432.071.751.870.180.809.85107.11092.317.69‘紫波’ב麒麟’2.470.901.691.280.220.7717.3275.960100.000平均值13.5889.2918.8978.552.56
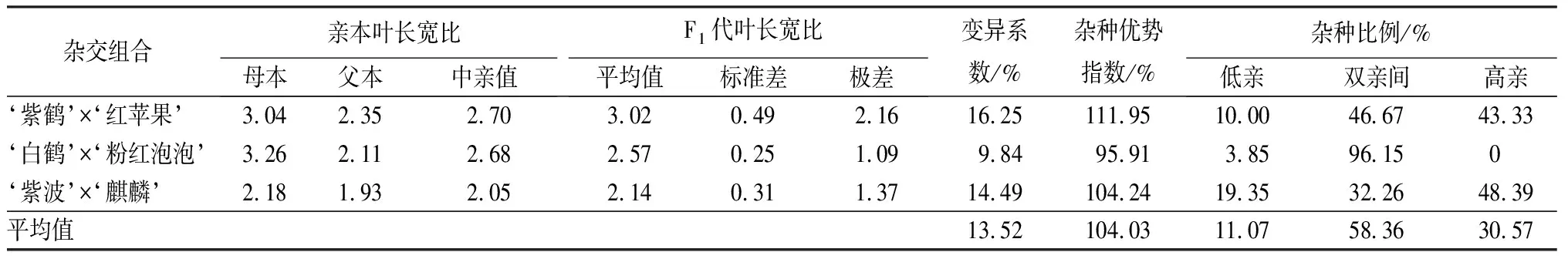
表10 杜鹃花品种间杂交F1代叶长宽比变异情况
2.3 杂交组合F1代数量性状间的相关性
据相关性分析结果(见表11),3个杂交组的数量性状之间具有较强的相关性,营养生长和生殖生长密切相关。花径与花梗长度、叶长、叶宽、叶长宽比都呈极显著相关,其中与叶长的相关性最强,相关系数为0.819。花梗长度与叶长、叶宽呈极正相关,与叶长宽比呈正相关。叶长与叶宽、长宽比均呈正相关。
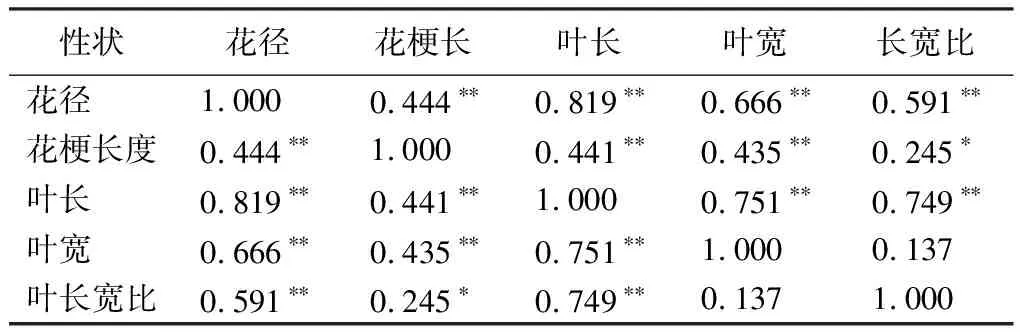
表11 杂交组合F1代数量性状间的相关性
3 结论与讨论
花色是杜鹃花育种中最重要观赏性状。本研究中花色划分为白色、红粉、红色、紫红、紫粉、紫色和复色共7类,按照色系归为白色系、红色系、紫色系和复色系共4个色系。3个杂交组合子代花色均出现了丰富的变异,当双亲花色差异越大时,子代花色变异也更为丰富,而且与父本花色类似的子代占比最高,说明父本花色对子代的花色影响较大。同时结果也表明,不同杂交组合的子代花色分离出现明显差异。杂交组合‘紫鹤’ב红苹果’中亲本花色均属紫色系,子代均呈现深浅不同的紫色或紫红色。杂交组合‘白鹤’ב粉红泡泡’中母本是白色系,父本是红色系,子代中白色花色占比为22.6%,远低于粉红色子代的比例,与刘晓青等[16]推测白色是杜鹃花的弱性基因观点一致。该组合子代还出现了亲本没有的紫色系花色,可能紫色的基因来源于父本‘白鹤’。因为父本‘白鹤’是锦绣杜鹃品种之一,据Riek et al.[20]分析,锦绣杜鹃的亲本之一为毛白杜鹃(Rh.mucronatum),虽然花色为白色花,但携带紫色基因。‘紫波’ב麒麟’杂交子代花色变异最为丰富,子代除了亲本紫色、红色系花色外还出现了少量的白色系和复色系,而刘晓青等[16]认为亲本中有白色系和复色系,子代才会产生白色系和复色系,与本研究结果不一致。花色呈现与花朵中的花色苷种类有关,根据HPLC分析,红色花含有2~4个主要的花色苷类型,而紫色花有2~6个类型[20],所以紫色花后代花色分离更为丰富。杜鹃花花色呈现不但与花色苷种类有关,也与由花色苷形成的色素量有关。杜鹃花花色遗传是由3对主效等位基因控制的[20],具有质量遗传特点,但是子代花色深浅差异,说明色素含量是有差异的,又具有数量遗传的特征,因此推测浅色花或白色花是由于色素含量降低造成的。
表型多样性是受遗传多样性和环境多样性的共同影响的,既具有稳定性,又具有变异性[21]。本研究中只有‘紫波’ב麒麟’的子代出现少量复色系花色,且‘紫波’ב麒麟’组合的亲本形态差异较大,推测复色现象是因为该组合2个亲本的亲缘关系较远而产生的。赵蕴阳[22]研究发现远缘杂交可能会引起转座子激活,从而导致表观遗传不稳定性,进而引起基因表达和表型变异。刘青林等[23]、李军等[24]、Morita et al.[25]认为转座子激活引起基因不稳定表达会形成花色嵌合体。虽然复色性状增加了观赏植物的多样性,但是其后代性状也易发生改变[26-27]。本研究中出现的复色性状能否稳定遗传,且遗传能力如何还不清楚,尚需要进一步研究。
花冠裂片内斑点是影响杜鹃花性状的重要因子之一。本研究中杂交组合‘紫鹤’ב红苹果’亲本花冠裂片内均含斑点,子代花冠裂片内均含斑点;杂交组合‘白鹤’ב粉红泡泡’亲本花冠裂片内均含斑点,子代出现少数不含斑点植株;杂交组合‘紫波’ב麒麟’父本花冠裂片内不含斑点,母本花冠裂片内含斑点,子代出现不含斑点植株。花斑的出现情况表明,花冠裂片内斑点具有显性遗传的特征,这与刘晓青等[16]研究结果一致。此外,百合[28]、三色堇(Violatricolor)[29]等花被上的花斑也具有显性遗传特点,所以推测斑点在杜鹃花遗传中为显性遗传。在花冠类型上,3个组合杂交试验的父本均为套筒,母本为单瓣或重瓣,子代的花冠类型出现了完全不同的分离比例,说明花冠类型的遗传相对复杂,还需要增加正反交试验和世代分离,分析其遗传规律。
本研究通过对杜鹃花的花部和叶部的数量性状进行相关性统计,发现多数花部性状和叶部性状呈正相关关系。其中,花径和叶长的相关系数最高,为0.819,说明杜鹃花的营养生长和生殖生长具有较强的联系性;叶长与叶宽、叶长宽比呈极显著相关,相关性系数分别是0.751和0.749,说明叶长一定程度上决定了叶宽;花径和花梗长度之间呈极显著相关,说明花梗长度一定程度决定了花径。所以在选择育种过程中,可以利用性状之间的相关性对决定性状的基因进行宏观筛选,通过较容易测量的叶长和花梗长度等性状推测其他不宜测量的基因,并筛选出目标性状,以提高育种效率[30-31],还可以利用变异系数高的性状作为指示性状对变异系数低且相关性强的性状进行间接选择,以提高育种效果[32]。此外,针对成熟期才表现出来的性状也可以利用这种相关性进行预测[32]。如:杜鹃花实生苗的幼龄期较长,有鳞杜鹃从播种到开花需要3~5年,无鳞杜鹃则需要5~10年,甚至有些种类的杜鹃花需要十几年[33]。可以通过分析叶片形态的遗传变异规律和杂种优势表现,利用叶部性状和花部性状的相关性起到早期鉴定和预先选择的作用[34-35],达到筛选目标性状,缩短杜鹃育种周期的效果。
猜你喜欢
中国生育健康杂志(2022年4期)2022-11-25
金沙江文艺(2022年3期)2022-03-25
第二课堂(小学版)(2019年4期)2019-05-13
安徽农学通报(2017年9期)2017-05-19
中国医药导报(2017年6期)2017-04-06
农业与技术(2016年22期)2017-03-07
亲子(2016年6期)2016-05-14
江苏农业科学(2015年5期)2015-10-20
科技资讯(2014年26期)2014-12-03
现代农业科技(2009年1期)2009-04-09
