
瞬时受体电位离子通道与银屑病
2021-07-26 08:11钟尤马玉香强磊
药学研究 2021年6期
钟尤,马玉香,强磊
(中国药科大学,江苏 南京 211198)
银屑病在民间俗称“牛皮癣”,是一种发病率较高的慢性、顽固性皮肤病,具有遗传性、病情反复、临床表现多样等特点[1-2]。全世界患病率约为2%~4%,是成年人最常见的皮肤病之一[3]。银屑病的主要病理表现为表皮角质化过度及角化不全、棘层细胞肥厚、免疫细胞浸润及血管扩张等。该病的发病原因复杂,发病机制尚未完全阐明,至今尚未找到根治的方法。从目前的研究来看,银屑病是一种由环境因素刺激、多基因遗传控制、免疫机制介导的慢性炎症性皮肤病,其发生发展过程中涉及血管增生迁移、天然T细胞分化、炎性细胞浸润、角质形成细胞增殖等多个环节[4]。参与银屑病免疫反应的细胞有很多种,包含T细胞、中性粒细胞(neutrophil)、树突状细胞(dendritic cell,DC)以及巨噬细胞等[1]。银屑病皮损处有明显的炎症浸润,多种炎症因子相互作用,产生级联放大作用并形成炎症持续环路,使银屑病患者局部炎症反应异常激活[1]。银屑病患者的皮肤组织病变十分明显,其中最显著的病理特征为表皮增厚,在银屑病皮损中角质形成细胞(keratinocyte,KC)生长动力学呈现出一种特殊紊乱状态,表现为KC的过度增殖和异常分化[5]。
瞬时受体电位通道(transient receptor potential channel,TRP)是细胞膜上的一类阳离子通道,广泛参与感知各种细胞内外刺激及维持离子稳态等多种生命活动。按照TRP作用和组织分布的差异,可分为瞬时受体电位香草酸亚型(TRPV)、经典瞬时受体电位(TRPC)、瞬时受体电位锚蛋白(TRPA)、M型瞬时受体电位(TRPM)、多囊蛋白类瞬时受体电位(TRPP)、黏脂质类瞬时受体电位(TRPML)和无机械感受器电位C(TRPN)7个亚家族[6]。TRP通道在细胞快速感受外界刺激以及细胞增殖、分化和凋亡等方面起着至关重要的作用,其表达或者活性异常能够引起严重的人类疾病[7]。TRP通道能够接受外界环境中的各种刺激,例如温度、pH或渗透压的变化、压力、剪切应力、机械拉伸、氧化应激、磷脂和磷脂代谢物等。此外,它们能对组织损伤和炎症过程中产生的内源性物质和信使产生响应[8]。
瞬时受体电位离子通道表达在皮肤的许多细胞或结构上,如感觉神经、角质形成细胞、黑素细胞、免疫细胞、内皮细胞和成纤维细胞等[9]。TRP通道对维持正常皮肤功能(如皮肤屏障、温度感觉)起重要作用,并且病理条件下也发挥着重要作用。TRP通道参与多种皮肤病的发生发展,如皮肤炎症、皮肤癌和毛发异常等[10-12]。因此,它们还可以作为许多皮肤疾病新疗法的靶标,包括炎症性皮肤病、遗传性疾病(例如Darier氏病)、自身免疫病、纤维化疾病和伤口愈合延迟、色素沉着疾病、恶性黑色素瘤或非黑色素瘤皮肤癌[9]。近年来,TRP通道在银屑病发病机制中的作用引起了人们的重视。TRP通道在细胞稳态和生长控制中起着关键作用,并且参与了皮肤瘙痒感觉的传导、神经元与非神经组织免疫和炎症调节以及角质形成细胞增殖分化的调控等一系列有关银屑病发生发展的病理活动。因此,深入了解TRP通道与银屑病的关系,对研究银屑病发病机制,开发新的药物治疗靶点有着非常重要的意义。本文主要综述了目前研究中与银屑病生理病理过程关系较为密切的几个TRP家族与银屑病的关系(见图1)。
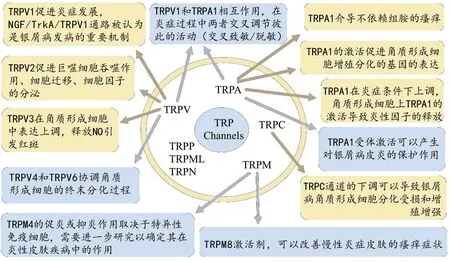
图1 TRP通道的分类及其参与银屑病发生发展的研究进展
1 TRPV亚家族与银屑病
TRPV1通道又称为辣椒素受体或香草酸受体亚型(VR)1,辣椒素或温度变化(>43 ℃)可以直接激活该通道。随着对TRPV1通道研究的深入,人们发现TRPV1也能被低PH和脂氧合酶产物如白三烯B4激活,并被ATP、缓激肽、前列腺素(PGE2,PGI2)、组胺、活性氧物种、蛋白酶和PAR2致敏[10,13]。研究表明TRPV1在非神经元皮肤细胞中具有促炎作用。在体外,已证明辣椒素(TRPV1激动剂)可以诱导角质形成细胞环氧合酶2(COX-2)的上调并增加炎性介质(如IL-8和PGE2)的释放[14-15]。除体外实验外,体内的研究也表明TRPV1在炎性皮肤疾病中的促炎作用。例如,辣椒素增强了豚鼠的过敏性接触性皮炎[16]。在无毛小鼠中,抑制TRPV1的活化可以降低紫外线诱导的MMP-2、MMP-3、MMP-9或MMP-13以及促炎细胞因子IL-1β、IL-2、IL-4、TNF-α和COX-2的mRNA和蛋白表达水平[17]。神经源性TRPV1还通过神经源性机制从感觉神经末梢释放降钙素基因相关肽(CGRP)和P物质(SP)诱发急性炎症(这种炎症被定义为神经源性炎症)[18]。研究显示TRPV1抑制剂在IMQ诱导的银屑病小鼠模型中,可在很大程度上预防瘙痒和皮肤炎症的发生,同时降低背根节细胞(DRG)神经元中降钙素基因相关肽(CGRP)的表达[19]。与WT小鼠相比,IMQ诱发的TRPV1-KO小鼠银屑病的临床评分、TEWL评分、皮肤增生程度、Munro微血管面积(MM)和血管生成均显著降低,表明皮肤炎症和屏障缺损减轻[20]。TRPV1与炎症过程中各类细胞因子的紧密关系,使其与银屑病发生发展产生了密切的联系。在银屑病斑块的角质形成细胞中,TRPV1的表达水平高于正常人[21]。NGF/TrkA/TRPV1通路被认为是银屑病发病的重要机制。银屑病患者的皮损在真皮和表皮都有更丰富的神经支配,与非皮损或非瘙痒性皮损相比,银屑病斑块中存在大量NGF免疫反应性角质形成细胞以及高表达TrkA的神经纤维[22]。
TRPV2在许多免疫细胞中表达,主要有巨噬细胞,肥大细胞,嗜中性粒细胞和淋巴细胞[23-24]。最近的研究得出结论,TRPV2对于肥大细胞和巨噬细胞功能至关重要。敲除巨噬细胞TRPV2显著地抑制了NF-κB通路,TRPV2对于鼠巨噬细胞的吞噬作用、迁移和细胞因子(TNF-α、IL-6)的产生有重要意义。同时,TRPV2介导了鼠和人培养的肥大细胞系中的脱粒和活化[25-27]。因此,开发靶向TRPV2的药物可能在巨噬细胞起关键作用的疾病例如银屑病中发挥有益作用。值得注意的是,人类皮肤的角质形成细胞也有TRPV2的表达[28],但角质形成细胞衍生的TRPV2的确切作用仍不清楚,需要进一步研究。
TRPV3在瘙痒性银屑病皮肤中的表达升高。在用TRPV3-激动剂(丁子香酚)刺激后,小鼠角质形成细胞释放出促炎细胞因子IL-1α,缓激肽以及组胺使TRPV3致敏[29]。此外,在小鼠角质形成细胞和背根节细胞(DRG)共培养系统中,加热后角质形成细胞衍生的TRPV3参与了ATP的释放,并且证明ATP降低了TRPV3对其激动剂的敏感性[30-31]。出除此之外,调节重要炎症过程的花生四烯酸可增强小鼠角质形成细胞的TRPV3通道功能。值得注意的是,TRPV3在皮肤稳态,NO代谢和伤口愈合中也起作用。在培养的角质形成细胞中TRPV3的激活可诱导NO的产生,这促进了角质形成细胞的迁移,并且在体内实验中,TRPV3激活改善了伤口的愈合[15]。基于NO调节血管紧张度和血管舒张的事实,可以推测角质形成细胞通过激活TRPV3释放NO介导血管舒张进而引发红斑[32]。这些研究提示TRPV3可能在银屑病主要症状——红斑中发挥作用。
TRPV4与连接蛋白(如β-catenin和e-cadherin)在功能上共表达,从而形成适当的表皮屏障[33]。TRPV4和TRPV6通过调节角蛋白和总苞素的表达来协调角质形成细胞的终末分化过程[34]。体内研究表明,小鼠Trpv6基因的缺失会导致皮肤钙水平下降,角质层厚度变薄,20%的动物会出现脱发和皮炎[35]。总之,TRPV4、TRPV6可能对皮肤屏障功能、角化细胞增殖和分化很重要,因此可能作为银屑病中针对角质形成细胞增殖分化异常的一个有前途的靶点。
2 TRPA1与银屑病
类似于TRPV1-4,TRPA1也可被温度变化激活,从而介导热敏。TRPA1主要在初级感觉神经元中表达,皮肤角质细胞中也有少量表达。TRPA1对瘙痒的感觉和知觉很重要。特别地,TRPA1介导不依赖组胺的瘙痒(这是银屑病等慢性瘙痒疾病所必需的),因此TRPA1是可以作为开发抗银屑病瘙痒药物的良好靶标[36-37]。事实上,TRPA1也在角质形成细胞增殖、分化和屏障功能中发挥作用,TRPA1激活可引起未分化的角质形成细胞中钙水平的升高[38]。在原代人角质形成细胞中,TRPA1的激活促进角质形成细胞增殖分化的基因(热休克蛋白、细胞周期蛋白、细胞周期蛋白依赖性激酶等)的表达[39]。TRPA1在炎症条件下似乎被上调,一些内源性促炎和伤害感受介质(前列腺素、缓激肽、蛋白酶、氧化应激)可以激活并敏化TRPA1[40-41]。TRPA1的激活通过感觉神经末梢或非神经元细胞释放SP和CGRP而导致炎症。体外实验显示,角质形成细胞上TRPA1的激活导致IL-1α,IL-1β和PGE2 的释放[39]。在体内,肉桂醛激活TRPA1会引起耳部水肿(SP依赖性)和白细胞浸润,而 TRPA1拮抗剂可以预防肉桂醛诱导的耳部水肿[42]。非常有趣的是,最近的研究表明,神经元与非神经元的TRPA1受体激活可以产生对银屑病皮炎的保护作用[43]。在IMQ诱导的银屑病模型中,TRPV1功能被抑制时,银屑病样皮炎症状减轻,而在TRPA1基因敲除小鼠和TRPA1拮抗剂治疗中银屑病样症状显著增强[44]。TRPV1和TRPA1通道可能在基础条件下在感觉神经元中形成异二聚体,并且在炎症过程中两者都能够交叉调节彼此的活动(交叉致敏/脱敏)[43]。这表明TRPA1可能通过调节神经元的兴奋性起作用,可能是通过调节TRPV1的活性发挥作用。同时,研究还证明,在CD4+T细胞向T helper表型1(Th1)激活过程中,TRPA1和TRPV1之间存在相互作用。在IMQ诱导的皮肤炎症中,TRPA1也在CD4+T细胞中共定位[44]。尽管TRPA1 的促炎作用已经被证实,但不断有新的证据表明其对于皮肤炎症的保护作用,对于TRPA1与银屑病的关系还有待进一步研究探索。
3 TRPC亚家族与银屑病
TRPC1、TRPC4、TRPC5、TRPC6和TRPC7是TRPC通道在表皮角质形成细胞中的重要代表[45]。由于钙是表皮角质形成细胞更新的主要调节剂,钙的内流可以触发角质形成细胞分化过程,因此多项研究表明TRPC通道在角质形成细胞分化和增殖中发挥着关键作用。TRPC1、TRPC4、TRPC6介导钙内流并诱导角质形成细胞的终末分化[46]。TRPC6激活剂(hyperforin)可以引起角质形成细胞TRPC6介导的钙内流,并且TRPC6激活随后诱导角质形成细胞的分化并抑制增殖[47]。而TRPC通道的下调可以导致银屑病角质形成细胞分化受损和增殖增强[48]。TRPC4也可以通过DAG刺激并增强钙进入细胞质。TRPCs功能受损可诱导角质形成细胞增殖并增加终末分化。越来越多的证据表明,银屑病斑块中TRPC通道的表达水平受到抑制[47]。未来对于TRPC 亚家族深入的研究,也许可以为银屑病药物开发打开新的思路。
4 TRPM亚家族与银屑病
TRPM通道在角质形成细胞和许多免疫细胞(T细胞、肥大细胞、粒细胞和单核细胞)都有表达,使其成为皮肤病治疗的潜在靶标[49]。然而,它们在疾病状态中的功能作用目前还不完全清楚。目前的研究发现,降温和薄荷醇能有效抑制人和小鼠的瘙痒行为,而TRPM8是主要的冷传感器和薄荷醇目标,使用选择性和有效的TRPM8激活剂,可以改善各种皮肤条件下的瘙痒症状[50]。众所周知银屑病等炎症性皮肤病可通过调节免疫细胞功能来改善。研究已经发现了TRPM2、TRPM4和TRPM7在调节免疫细胞中的功能性作用。例如,TRPM2在原代人CD4+T细胞中表达,并在T细胞刺激后上调[51]。TRPM2在T细胞中发挥作用[52],但其对 T细胞调节作用仍不清楚。此外,TRPM2与抗原刺激后肥大细胞脱粒,以及受ADP-核糖调节的粒细胞和单核细胞趋化因子产生有关[53-54]。TRPM4被报道可以介导免疫细胞中的膜去极化。因此,推测TRPM4激活可调节T细胞中的钙信号,从而影响细胞因子的产生和肥大细胞功能(减轻肥大细胞脱粒)[55-56]。另一方面,TRPM4与树突状细胞迁移有关[57]。因此,TRPM4的促炎或抑炎作用取决于特异性免疫细胞,需要进一步研究以确定其在炎性皮肤疾病中的作用。还要注意的是,TRPM7也可在免疫细胞(淋巴细胞,肥大细胞)中表达,并被认为参与淋巴细胞和肥大细胞的增殖。目前,科研人员正努力开发更加高效、高选择性 TRPM8激活剂用以改善瘙痒症状,但要搞清楚TRPM通道在免疫细胞发挥什么作用,能否有机会开发新的银屑病药物靶点,还有很长的路要走。
综上所述,TRP通道主要参与了皮肤瘙痒感觉的传导、神经元与非神经组织免疫和炎症的调节以及角质形成细胞增殖分化的调控等一系列有关银屑病发生发展的病理活动。因此,深入了解TRPs与银屑病的关系,对研究银屑病发病机制,开发新的药物治疗靶点有着非常重要的意义。希望未来有更多TRP通道具体机制的研究,以便更深入地讨论TRP通道与银屑病的关系,并为银屑病的治疗提供新的信息和策略。
猜你喜欢
医学概论(2022年3期)2022-04-24
清华金融评论(2022年4期)2022-04-13
皮肤病与性病(2021年3期)2021-07-30
国际放射医学核医学杂志(2021年10期)2021-02-28
房地产导刊(2020年7期)2020-08-24
儿童故事画报·智力大王(2020年1期)2020-04-28
中国外汇(2019年14期)2019-10-14
凤凰生活(2018年6期)2018-06-25
健康之家(2016年10期)2016-10-28
母子健康(2015年8期)2015-12-13
