
胡麻-小麦轮作更替土壤的细菌群落多样性分析
2021-09-16 03:35王立光叶春雷李进京李静雯罗俊杰
干旱地区农业研究 2021年5期
王立光,叶春雷,陈 军,李进京,李静雯,罗俊杰
(甘肃省农业科学院生物技术研究所,甘肃 兰州 730070)
作物长期单一种植往往引起土壤理化性质、土壤自毒作用和微生物群落变化,从而不利于植株生长与产量,造成连作障碍[1-3]。轮作是克服连作障碍的有效种植模式之一,在农业生产中广泛应用[4-7]。有研究表明,轮作不仅可以改善土壤酶活性,降低土壤自毒作用,还影响土壤微生物群落的多样性和丰度,从而影响作物生长和产量[4, 8-12]。也有研究证实,连作制度的强化可以在减少农药投入的情况下利于杂草和土壤养分治理,从而提高农业生态系统的盈利能力及环境改善效果[13]。
研究发现,胡麻(Linumusitatissimum)作为我国西北和华北黄土高原地区重要的油料作物,其连作会导致连作障碍发生[9]。胡麻连作导致土壤水提液自毒作用显著加强,导致种子萌发降低、植株生长受到抑制,产量降低,并对部分土壤酶活性造成不利影响,与土壤养分、酶活性呈现显著的负相关[14-17]。而胡麻轮作小麦可以减弱土壤水提液对胡麻种子萌发和幼苗生长的自毒作用,并有利于维持土壤酶活性,与土壤养分及酶活性呈显著正相关,消减胡麻连作障碍的发生,利于胡麻生产[9, 18]。目前,关于小麦轮作消减胡麻连作障碍的研究虽然在土壤理化性质、土壤酶活性及土壤自毒作用等方面已经取得部分成果,但胡麻-小麦轮作更替过程中土壤细菌群落多样性及群落结构变化的研究少有报道,因此,本研究通过高通量测序技术对胡麻-小麦轮作更替土壤内细菌群落进行测定,研究轮作更替过程中土壤细菌群落变化,以期从细菌群落变化角度解析胡麻-小麦轮作消减连作障碍机制,从而为胡麻连作障碍消减提供理论依据,为胡麻产业的良性发展提供技术支撑。
1 材料与方法
1.1 试验材料及样地概况
胡麻试验品种为陇亚10号,由甘肃省农业科学院作物所提供。种植地点为甘肃省榆中县试验地(104°06′35.27″ E, 35°50′36.06″ N),海拔1 600 m,属于黄土高原丘陵沟壑半干旱高寒区,年均降水量约450 mm,样地区域土壤类型为黄绵土。试验地为浇灌地,胡麻生育期充分浇灌2次。
1.2 田间试验处理
田间试验设置胡麻-小麦轮作更替样地,分别为第一年胡麻种植(TC1)、第二年小麦轮作(TR)和第三年胡麻种植(TC2),每个处理设置3个重复,共9个小区,随机区组排列,每样区面积30 m2(6 m×5 m)。小区间用宽60 cm、高50 cm土埂分割,以防小区间互相影响。
1.3 土壤样品采集
试验样地作物栽培管理与当地大田作物相同,未做特殊处理。收获期分别采集第一年种植胡麻、第二年小麦轮作和第三年胡麻种植地根际土壤,采用“S”型布点对土壤样品进行采集,保证6个采样点,各采样点采样土壤充分混合,剔除枯叶、腐叶等,每小区获得土样2份。每种处理的3个平行土样充分混合,最终每种处理获得土壤样本2个,用四分法取适量土样放入无菌自封袋,迅速置于干冰上保存,带回实验室进行后续分析。
1.4 土壤样品DNA提取及高通量测序
准确称取土样0.1 g,按照土壤微生物DNA提取试剂盒(MOBIO PowerSoil©DNA Isolation Kit:12855-50)说明步骤分别提取各土样的总DNA,Drop2000测定提取获得的DNA浓度和纯度,1.0%琼脂糖凝胶电泳检测DNA完整性。设计带有barcode的融合引物,针对细菌16S rRNA的V3~V4区进行PCR扩增,扩增特异性引物序列为338F:ACTCCTACGGGAGGCAGCA,806R:GGACTACHVGGGTWTCTAAT。扩增后利用2.0%琼脂糖凝胶对扩增产物进行电泳、并切胶回收PCR产物,利用QuantiFluorTM-ST蓝色荧光定量系统进行定量检测,按照样本测序量要求,对每个样本进行相应比例的混合,合格样品交由美吉生物医药科技公司进行Miseq文库构建及高通量测序,测序平台为PE300。
1.5 测序数据处理与分析
测序结束后,对原始数据进行拼接、质量过滤处理,从而得到优化序列,进行OTU(Operational Taxonomic Uniits)聚类分析、物种组成分析、Alpha多样性分析、Beta多样性分析、物种差异分析及功能预测分析。其中,序列拼接利用FLASH、序列过滤利用QIME,聚类利用UPARSE pipeline软件,功能预测采用Tax4Fun进行分析。
2 结果与分析
2.1 测序样本稀释曲线分析
对样本测序序列进行随机抽样,以抽到的序列数与其对应的OTU的数目绘制稀释性曲线(图1,见87页)。图1表明,随着序列数的增加,各土壤样本的稀释曲线斜率变小,趋于平坦,更多的测序序列读取只能产生少量新的OTU,贡献率较小,说明测序数据量合理,测序结果已可以真实反映各土壤样本细菌群落指标。
2.2 土壤细菌OTU分布Venn图
优化测序结果后共获得237750条有效序列,按97%相似度归并后通过OTU聚类分析得到1870条OTU。对各测序土样得到的OTU进行分类表明,3类样品中共有的OTU为1625条,TC1、TR和TC2分别具有特异OTU为20、11和36条,仅TC1和TR共有的OTU为61条,TC1与TC2共有而TR没有的OTU为50条,TC1不具有而TR和TC2共有的OTU为67条(图2)。
2.3 土壤细菌物种多样性分析
采用Chao指数、Ace指数、Shannon指数和Simpson指数对不同根际土壤细菌群落Alpha多样性进行分析,从而反映细菌群落的丰度和多样性。由表1可知各样本文库覆盖率均大于99.4%,说明测定结果能够真实反映各样本的细菌群落组成。不同处理的Shannon指数表现为TC1>TR>TC2, Simpson指数表现为TC2>TR>TC1,细菌群落丰度(Chao指数和Ace指数)则表现为TC1>TR>TC2,说明在轮作更替过程中细菌多样性和丰度都是降低的。
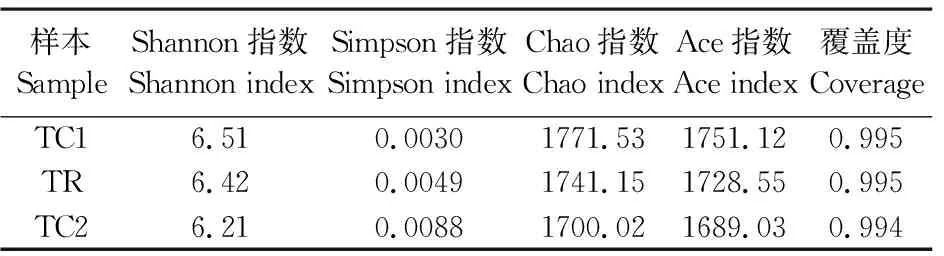
表1 土壤样本中细菌的Alpha多样性分析Table 1 Alpha bacteria diversity statistical in soil samples
2.4 土壤细菌物种丰富度分析
样本检测出的细菌主要分布在33个门,75个纲,143个目,263个科,433个属,811个种。以门分类水平对土壤细菌分析发现,TC1、TR和TC2土壤具有的细菌门数分别为33、33、32个,其中有32个细菌门为三者所共有,TC1和TR土壤内的Peregrinibacteria菌门在TC2土壤内未检测到(图3A)。如图3B所示,根际土壤细菌优势菌门(相对丰度>0.01)分析发现,3种土壤内的优势菌门种类相同、数目相同(均为11个),但各样本内优势细菌门的相对丰度存在差异。变形菌门(Proteobacteria)在TC1和TC2处理下的丰度占比最高,其相对丰度分别为31.65%和28.53%,而在TR处理下其相对丰度为24.52%,在轮作更替中表现为先降低再升高。酸杆菌门(Acidobacteria)在TR处理下丰度占比最高,其相对丰度为26.13%,其在TC1和TC2处理下的丰度占比分别为19.36%和11.18%,表现为先升高再降低。放线菌门(Actinobacteria)在TC1和TR土壤内的相对丰度接近,分别为12.14%和13.79%,但在TC2处理下相对丰度为22.82%,明显高于TC1和TR。绿湾菌门(Chloroflexi)在TC1、TR和TC2中的相对丰度分别为9.69%、12.68%和10.53%,与酸杆菌门相似,都表现为先升高再降低,但TC1和TC2处理下的相对丰度较接近。芽单胞菌门(Gemmatimonadetes)在TC1处理下相对丰度为8.04%,而轮作小麦后相对丰度降低至5.41%,再次胡麻种植后相对丰度上升至6.01%。后壁菌门(Firmicutes)在TC2根际土壤内相对丰度为12.63%,远远高于TC1(3.29%)和TR(1.32%)处理下的相对丰度,种植更替中其变化规律为先降低后急剧上升。TC1和TR根际土壤内拟杆菌门(Bacteroidetes)的相对丰度接近,分别为7.04%和7.11%,而在TC2处理下根际土壤的相对丰度仅有2.89%。硝化螺旋菌门(Nitrospirae)的相对丰度在更替种植土壤内表现为逐渐降低,其相对丰度依次为3.01%、1.81%、1.52%。浮霉菌门(Planctomycetes)和 疣微菌门(Verrucomicrobiae)的变化趋势与绿湾菌门一致,都是先升高再降低,且TC1和TC2内相对丰度几乎相同。Latescibacteria菌门在轮作更替根际土壤内的相对丰度为TR(1.23%)>TC1(0.92%)>TC2(0.38%),其变化规律与酸杆菌门类似。
2.5 土壤细菌群落结构变化分析
对测序结果的主坐标分析(Principal Co-ordinates Analysis,PCoA)发现,轮作更替根际土壤细菌群落丰度差异较明显。如图4A所示,土壤PC1贡献率为67.46%,PC2的贡献率为26.1%,TC1分布在PC1和PC2的负值区域,且分布相对集中,而TR分布于PC1的负值区域和PC2的正值区域,分布也相对集中,TC2聚集到PC1正值区域及PC2的正负值区域,分布较远。基于Weighted Unifrac距离矩阵进行聚类分析,如图4B所示,TC1和TR聚为一类,TC2单独聚为一类,说明TC1和TR根际土壤细菌群落差异相对较小,而轮作小麦后再次种植胡麻使根际土壤细菌群落差异变大。
2.6 土壤细菌群落Tax4Fun功能预测分析
Tax4Fun可以将基于Silva数据库中的16S分类谱系转化为KEGG(Kyoto Encyclopedia of Genes and Genomes)数据库内原核生物的分类谱系,从而实现对16S rRNA基因序列进行KEGG功能注释,基于此,可获得OTU在KEGG各功能水平的注释信息及各功能在不同样本内的丰度信息。为探究胡麻-小麦轮作更替过程中根际土壤细菌变化情况,本研究采用Tax4Fun软件进行细菌功能预测分析(表2)。测序获得功能基因与KEGG数据库比对分析发现,所有细菌的基因序列在第一等级上注释到6类功能,分别为新陈代谢、环境信息处理、遗传信息处理、细胞过程、有机系统和人类疾病,其中新陈代谢、遗传信息处理和环境信息处理作为主要组成位居前3类,其占比分别为61.30%~62.70%、11.33%~11.36%和18.45%~19.13%。二级功能预测分析显示,其包含碳水化合物代谢、氨基酸代谢、膜运输、细胞生长与死亡、折叠、分类与降解、能量代谢和信号转导等41个子功能组成。其中涉及新陈代谢的有11项,涉及环境信息处理功能的3项,涉及遗传信息处理功能的4项,涉及细胞运动、有机系统和人类疾病的子功能分别为5项、8项、10项。

表2 土壤样本细菌群落功能基因的KEGG丰度Table 2 Abundance chart of KEGG of functional genes for soil samples
3 讨论与结论
土壤微生物在土壤养分吸收转化和作物生长发育过程中扮演重要角色,而土壤细菌是土壤微生物群落中的重要群体,在群落中起着重要作用。我们前期已经对胡麻-小麦轮作中的土壤理化性质、土壤酶活性及土壤自毒作用等进行了研究,但关于轮作更替过程中土壤细菌群落变化的研究并未进行,也未见相关报道[14,16,17,19]。王芳等[20]对不同轮作方式下大豆根际细菌的研究表明,轮作作物种类、轮作顺序等对土壤细菌多样性都会产生影响。尹国丽等[21]的研究显示,轮作时间和轮作作物对土壤细菌多样性的影响是普遍的。这些结果表明轮作模式在消减连作障碍的过程中不仅与土壤酶活性、土壤自毒作用有关,还与土壤微生物群落存在关联。本研究结果显示,胡麻-小麦轮作种植更替过程中,土壤细菌的丰度和多样性都是逐渐降低的,但TC1和TR土壤内根际土壤细菌差异较小,且优势菌门保持相对稳定,而与TC2土壤内细菌差异较大。这种变化差异也与报道的胡麻-小麦轮作条件下植株生长和产量的变化差异相一致[9,15]。这也说明,胡麻-小麦轮作模式下土壤细菌群落变化很可能是影响消减连作障碍的又一因素。
在连作过程中,土壤内细菌群落差异随连作年限增加而改变,但也受连作作物的影响而存在差别。但众多研究结果表明,放线菌门、变形菌门、酸杆菌门和绿湾菌门等一般都包含在优势菌门内[22-24]。苜蓿、大豆和硒砂瓜连作研究表明,随着连作时间延长,细菌群落的优势菌门和主要菌门的数量没有改变,但菌门丰度发生较大改变,也就说明连作障碍发生伴随着菌门丰度变化,这也被认为是导致连作障碍的主要因素之一[22-23,25]。本研究显示在胡麻-小麦轮作更替过程各土壤中的优势菌门相同,并未由于小麦轮作改变优势菌门数量和种类。同时,小麦轮作使优势菌门与TC1内的优势菌门丰度保持相近,变化较小,而与TC2土壤内优势菌门丰度有较大差别,尤其是放线菌门、酸杆菌门和后壁菌门。主坐标分析显示,在更替种植土壤内,代表TC1和TR的细菌群落分布距离较近,而代表TC2的细菌群落分布距离较远,说明TC2群落结构存在较显著差异。进一步的聚类分析显示,相同更替年限的平行样品聚集在一个分支,且TC1和TR分支较近,而TC2单独处于一个分支,与TC1和TR间聚类水平降低,表明TR细菌菌门所占比例、菌群结构组成与TC1相似。这些结果说明小麦轮作不是通过改变优势菌门数量,而是通过维持细菌优势菌门和主要菌门的稳定性而消减下茬胡麻连作障碍的发生。这也暗示细菌群落的变化应该是导致胡麻连作障碍的一个重要因素。
种植模式对土壤细菌群落结构和丰度的影响,往往会导致土壤细菌群落功能组成的改变,而土壤细菌群落功能组成与土壤环境关系密不可分[26-28]。研究证实,胡麻残茬根、茎及叶水提液都具有明显的自毒效应,在胡麻种植过程中,这些自毒物质往往会通过不同形式进入土壤,造成毒害,使土壤水提液表现出明显的自毒作用[17,19,29]。本研究通过Tax4Fun对细菌群落的功能进行了预测,结果显示,在轮作更替过程中,一级功能层上细菌群落在遗传信息处理功能上几乎没有差异,但是在新陈代谢、环境处理和细胞过程功能方面存在差异,这可能与土壤内自毒物质的变化有关,因此后续有必要对各土壤内自毒物质种类及含量进行测定,从而解析细菌群落功能差异变化。本研究通过高通量测序分析了胡麻-小麦轮作条件下根际土壤细菌群落的丰度及其变化规律,并对细菌群落功能进行了初步预测,这些将为改良胡麻种植技术及解决种植中连作障碍的发生提供理论依据。
猜你喜欢
安徽农业科学(2022年20期)2022-11-11
科学技术与工程(2022年26期)2022-11-01
——基于CMS 模型的实证分析
市场周刊(2022年9期)2022-09-29
现代农业科技(2022年15期)2022-08-17
中国农学通报(2022年14期)2022-06-01
油气田环境保护(2022年2期)2022-05-09
中国土壤与肥料(2021年5期)2021-12-02
四川农业科技(2021年7期)2021-12-01
农业科技与信息(2021年19期)2021-11-30
中国果业信息(2020年10期)2020-12-15
