
番茄褪绿病毒对Q型烟粉虱重要生物学参数及保护酶和解毒酶活力的影响
2021-10-19 13:37刘国霞李娇娇张安盛
环境昆虫学报 2021年5期
刘国霞,李娇娇,张安盛*
(1.山东省农业科学院植物保护研究所,济南 250100;2.山东省农业科学院生物技术研究中心,济南 250100)
近年来番茄褪绿病毒病(Tomatochlorosisvirus,ToCV)在番茄Lycopersiconesculentum产区普遍发生,成为继番茄黄化曲叶病毒病之外又一严重病害。发病植株表现叶脉间变黄,叶脉颜色加深,叶片增厚变脆,果实小、颜色偏白,产量和品质受到严重影响(刘永光等,2014)。番茄褪绿病毒在美国首次报道(Wisleretal.,1998),随后在欧洲(Accottoetal.,2001;Dovasetal.,2002)、地中海地区(Segevetal.,2004)以及我国台湾地区(Tsaietal.,2004)相继发现。我国最早于2012年在北京检测到ToCV(Zhaoetal.,2013),随后在山东(Zhaoetal.,2014)、江苏(Karwithaetal.,2014)等地迅速扩散,而且带毒率呈快速增长趋势(郑慧新等,2016),给番茄生产带来巨大威胁。
烟粉虱Bemisiatabaci属半翅目Hemiptera、粉虱科Aleyrodidae,是番茄褪绿病毒重要传毒介体,研究表明Q型烟粉虱对ToCV的传播扩散具有重要作用(Shietal.,2018)。病毒对媒介昆虫的生物学和生理指标可能产生有利、中性或有害的影响(Stoutetal., 2006),如携带番茄黄化曲叶病毒(Tomatoyellowleafcurlvirus,TYLCV)的B型烟粉虱存活率、产卵量、成虫寿命和体长明显降低或缩短,而携带该病毒的Q型烟粉虱相应指标未受影响,说明TYLCV对B型烟粉虱具有不利影响,而对Q型烟粉虱影响偏中性(Panetal., 2013)。研究发现在携带ToCV的番茄上取食的Q型烟粉虱产卵量增加,发育历期缩短,存活率和寿命未受影响,而且带毒Q型烟粉虱更喜好取食健康番茄,有助于病毒扩增(Shietal.,2018)。而Li等(2018)研究结果显示Q型烟粉虱在携带ToCV的番茄上取食,其产卵量、雌虫寿命和产卵期显著下降,而发育历期增加(Lietal., 2018)。内共生菌组成对Q型烟粉虱传播ToCV也有重要影响(Belloetal., 2019)。
解毒酶和保护酶是昆虫体内重要的两类酶系,对于抗逆和生理适应具有重要作用,是昆虫生理适应性的重要指标(Lozinskayaetal., 2004)。烟粉虱取食感染TYLCV的番茄导致体内三大保护酶系过氧化物酶(POD)、超氧化物歧化酶(SOD)和过氧化氢酶(CAT)活性持续升高,解毒酶羧酸酯酶(CarE)和谷胱甘肽S转移酶(GST)活性先提高后下降,保护酶、解毒酶活性的变化对于烟粉虱在带毒番茄上的适应性有重要作用(曹增等,2015)。本文对携带ToCV的Q型烟粉虱在非病毒寄主植物棉花上的发育历期、产卵量、成虫寿命和体长进行研究,并测定了带毒和无毒烟粉虱主要保护酶和解毒酶生物活性,以期明确ToCV对烟粉虱种群扩张的影响,为了解ToCV传播扩散的流行病学机制奠定基础。
1 材料与方法
1.1 供试虫源与植物
供试虫源:无毒烟粉虱采自山东省农科院植保所实验农场棉花植株,接到日光温室中健康番茄(奔六)苗上,在温度26℃±1℃,相对湿度60%±5%,光周期L∶D=16 h∶8 h的条件下饲养。烟粉虱经以mtCOI为基础的CAPS方法鉴定为Q型(刘国霞等,2014)。
供试毒源:病毒植株采自寿光市稻田镇西稻田村日光温室,将病毒植株移栽到山东省农科院植保所日光温室盆栽种植,接入Q型烟粉虱成虫,定期添加番茄带毒植株和健康番茄苗,同时移出衰老死亡番茄植株,保证番茄植株持续带毒。经PCR检测烟粉虱带毒率超过90%。
供试植物:取5~7叶期棉花(鲁棉研37)用于生物学实验。
1.2 烟粉虱携带ToCV病毒检测
采集被ToCV感染的番茄植株上烟粉虱成虫及初羽化成虫各10头,参照李娇娇等(2018)单头虫体采用Trizol方法提取RNA,利用RT-PCR检测烟粉虱携带ToCV情况。烟粉虱在棉花上产卵24 h后,收集虫体利用同样方法提取RNA,检测烟粉虱带毒情况。
1.3 ToCV对烟粉虱生物学指标的影响
1.3.1发育历期和后代体长的测定
用内径2 cm、高0.5 cm的有机玻璃管和120目防虫网自制微虫笼,取10对带毒和无毒烟粉虱分别置于微虫笼中,固定在棉花(6~7叶期)叶片背面,放于人工气候箱中(温度26℃,相对湿度60%,光周期L∶D=14 h∶10 h),24 h后除去烟粉虱和微虫笼,在叶片上做好标记,解剖镜(Nikon, C-DSS230)下统计产卵数量。每天检查由卵发育到1龄、2龄、3龄、4龄和成虫的数量(罗晨等,2006)。初羽化成虫在解剖镜下区分雌雄,用测微尺测量虫体体长。实验设10个重复。
1.3.2产卵量和成虫寿命
取带毒和无毒烟粉虱初羽化成虫1对,分别接于1%琼脂保湿的棉花叶片的玻璃管中,倒置放于人工气候箱中(温度26℃,相对湿度60%,光周期L∶D=14 h∶10 h),每3 d记录产卵量,成虫转移至新叶片。每天观察成虫存活情况,若雄虫死亡则补充,直至雌虫死亡。重复30次。
1.4 ToCV对烟粉虱重要保护酶和解毒酶活力的影响
1.4.1烟粉虱体内保护酶活力测定
分别取100头带毒和无毒烟粉虱雌虫,加500 μL生理盐水冰浴匀浆,8 000 rpm离心20 min取上清液备用。采用考马斯亮蓝G250法测定酶源样品的蛋白含量。
采用过氧化物酶(POD)测试盒、超氧化物歧化酶(SOD)测试盒和过氧化氢酶(CAT)测试盒(均为南京建成生物工程研究所生产)按照试剂盒操作要求测定带毒烟粉虱和无毒烟粉虱体内POD、SOD和CAT活力。活性单位均采用试剂盒规定的单位每毫克蛋白(U/mgprot)。每个处理重复3次。
1.4.2烟粉虱体内解毒酶活力测定
分别取100头带毒和无毒烟粉虱雌虫,按照羧酸酯酶(CarE)测试剂盒要求制备样品,用于测定谷胱甘肽S转移酶(GST)测试盒和乙酰胆碱酯酶(AChE)的样品制备方法同1.3.1。按试剂盒(均为南京建成生物工程研究所生产)说明书测定带毒烟粉虱和无毒烟粉虱体内CarE、GST和AChE活力。活性单位均采用试剂盒规定的单位/每克蛋白(U/mgprot)。每个处理重复3次。
1.5 数据分析
采用SPSS 17.0统计软件中独立样本t检验进行处理间实验数据的差异性分析。重复数据计算其平均值和标准差(SD)。
2 结果与分析
2.1 烟粉虱携带ToCV病毒情况
采集生物学实验中带毒烟粉虱同一片叶子上的烟粉虱成虫,检测带毒率发现,除1头雌成虫未扩增到特异条带外,其余7头雌虫和1头雄虫均能扩增到ToCV病毒特异条带(图1-A),带毒率为88.89%,说明带毒烟粉虱可用于随后的生物学实验和酶活性分析。带毒烟粉虱在棉花上取食产卵24 h后,取24头烟粉虱检测带毒情况,发现有62.5%的烟粉虱携带ToCV(图1-B)。说明烟粉虱在棉花上取食24 h期间,大部分依然带毒。
在ToCV感染的番茄上,初羽化烟粉虱7头雌虫和3头雄虫均扩增到ToCV病毒特异条带(图1-A),1头雄虫扩增带较弱,说明初羽化成虫携带病毒率为100%。说明在ToCV感染的番茄上初羽化烟粉虱可用于测定产率量和雌虫寿命试验。
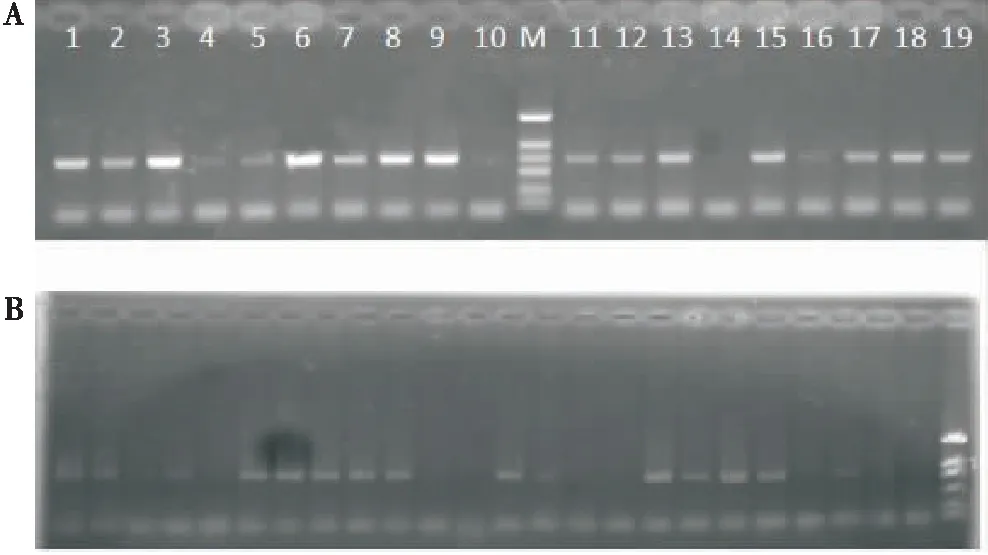
图1 烟粉虱携带ToCV病毒RT-PCR检测
在健康番茄上取食的烟粉虱未检测到ToCV。
2.2 ToCV对烟粉虱生物学指标的影响
带毒和无毒烟粉虱在棉花上各龄期的发育历期见表1。由表1可知,卵和4龄若虫发育期最长,为6.35 d左右,其次为1龄,为3.5~3.9 d,卵到成虫整个发育历期为21 d左右。带毒烟粉虱在1龄若虫历期较无毒烟粉虱长,但差异不显著,其它龄期历期及整个发育历期与无毒烟粉虱相比未表现出差异。
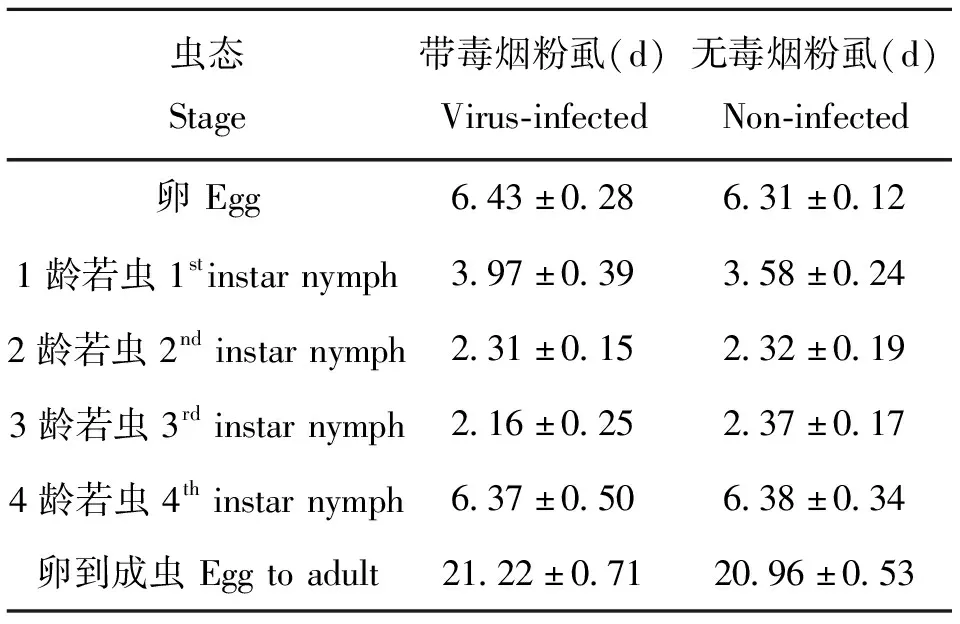
表1 带毒和无毒烟粉虱的发育历期
利用解剖镜测量了带毒和无毒烟粉虱在棉花上产卵羽化的后代体长,雌虫约0.8 mm,雄虫约0.74 mm。带毒烟粉虱雌虫长度比无毒雌虫短(P<0.05),雄虫体长无差异。单雌产卵量约80粒,寿命约18 d,带毒和无毒烟粉虱在棉花上的单雌产卵量和寿命无明显差异。
2.3 ToCV对烟粉虱重要解毒酶和保护酶活性的影响
采用考马斯亮蓝蛋白定量方法制定标准曲线,对提取的带毒和无毒样品蛋白含量进行测定,样品浓度在0.25~0.30 mg/mL。
携带ToCV的烟粉虱和无毒烟粉虱体内保护酶和解毒酶含量见表3。带毒烟粉虱体内超氧化物歧化酶(SOD)活性高于无毒烟粉虱,但差异不显著(P>0.05)。带毒烟粉虱过氧化氢酶(CAT)活性是无毒烟粉虱的3.36倍,差异极显著(P<0.01)。带毒烟粉虱过氧化物酶(POD)活性降低,差异不显著。

表2 带毒和无毒烟粉虱成虫体长、产卵量和雌虫寿命
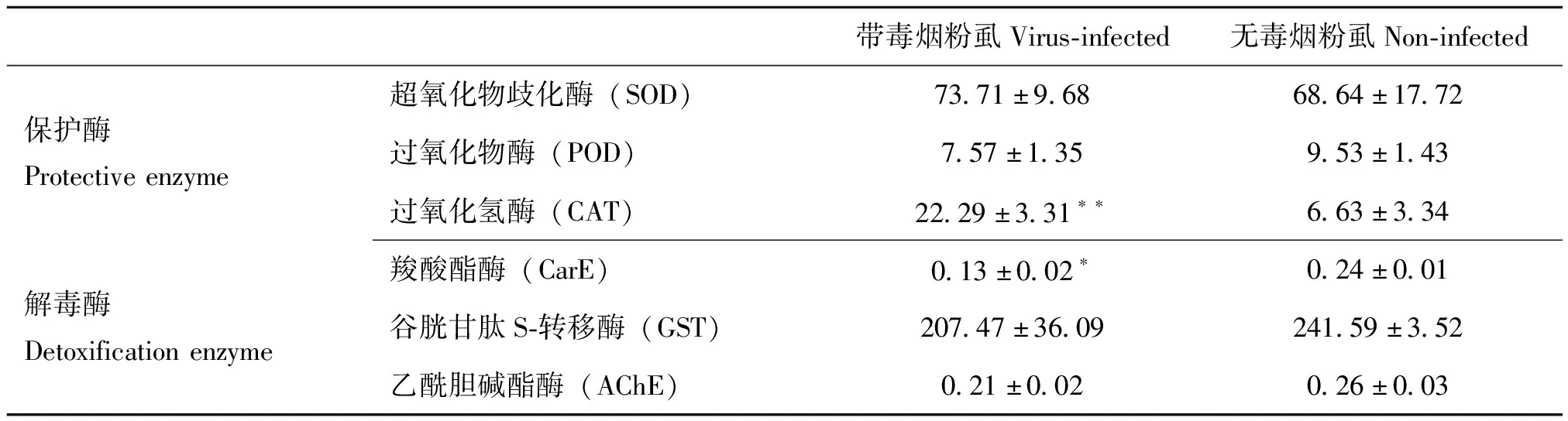
表3 带毒和无毒烟粉虱体内保护酶和解毒酶活力
解毒酶中,带毒烟粉虱羧酸酯酶(CarE)活力降低,为无毒烟粉虱的54%,差异显著(P<0.05)。带毒烟粉虱谷胱甘肽S-转移酶(GST)和乙酰胆碱酯酶(AChE)活力带毒烟粉虱略低于无毒烟粉虱,差异不显著。
3 结论与讨论
ToCV是韧皮部限制性病毒,自然条件下只能依靠媒介昆虫粉虱传播(王富等,2016),B型烟粉虱和纹翅粉虱Bemisiastriata传毒效率较高(Wintermantel and Wisler,2006)。近年来研究发现Q型烟粉虱也可有效传播ToCV(李娇娇等,2018),番茄ToCV发病率随Q型烟粉虱种群数量增加和取食时间延长而升高(代惠洁等,2016)。因此推测ToCV的广泛传播与Q型烟粉虱的发生危害有密切关系。病毒对寄主的影响分为直接影响和间接影响,Li等(2018)研究了ToCV对Q型烟粉虱的间接影响,发现Q型烟粉虱在感染ToCV的番茄上产卵量、产卵天数和雌虫体长显著降低,认为ToCV侵染对Q型烟粉虱有不利影响,可能跟ToCV侵染促使番茄释放更多的萜类化合物有关(Fereresetal., 2016)。本试验研究了ToCV对Q型烟粉虱的直接影响,发现在ToCV的非寄主植物棉花上,带毒烟粉虱在发育历期、产卵量、雌虫寿命方面和无毒烟粉虱无显著差异,仅后代雌虫体长明显短于无毒烟粉虱,说明ToCV对Q型烟粉虱生长发育和种群增长的直接影响较小。携带TYLCV的Q型烟粉虱在非寄主植物棉花上的各项生物学指标影响不大(Panetal., 2013),跟本研究ToCV对Q型烟粉虱生物学的影响结果一致。棉花和番茄都是烟粉虱的适宜寄主(张永军等,2003),在棉花上带毒烟粉虱随着时间延长体内ToCV病毒逐渐消失,研究发现雌虫最长持毒时间为7 d(Weietal., 2019),因此在本实验中烟粉虱生物学参数基本没有受到病毒的影响,雌虫体长的减小可能对于Q型烟粉虱种群增长有一定影响。
保护酶是清除自由基的重要物质,SOD、CAT和POD是昆虫体内主要的保护酶系,其中SOD将部分自由基催化成H2O2,CAT和POD可催化分解H2O2成为H2O和O2,从而使细胞内自由基维持在低水平,保护机体免受伤害(Krishn and Kodrik,2006)。研究证实保护酶活性跟烟粉虱寄主适应性相关(周奋启等,2011)。携带ToCV的Q型烟粉虱SOD和POD活性与无毒烟粉虱无显著差异,CAT活性是对照的3.36倍,差异达到极显著水平,说明取食ToCV侵染的番茄使机体产生了大量的H2O2,Q型烟粉虱通过提高CAT活性而提高分解H2O2的能力,从而保护机体免受自由基毒害。本研究结果与曹增等(2015)、陈晨等(2016)研究结果一致,B型烟粉虱在TYLCV感病番茄上取食3种保护酶(SOD、CAT和POD)活性持续升高,而Q型烟粉虱取食72 h后SOD和POD活性升高,取食30 d后CAT和POD活性比对照显著升高(曹增等,2015),白背飞虱Sogatellafurcifera(Horváth)取食南方水稻黑条矮缩病毒(Southernriceblack-streakeddwarfvirus,SRBSDV)后这3种保护酶活性显著提高(陈晨等,2016)。
CarE是烟粉虱体内对杀虫剂进行解毒代谢的重要解毒酶,田间抗性品系CarE活性明显高于敏感品系(何玉仙等,2007)。AChE是昆虫中枢神经系统中主要神经递质-乙酰胆碱的分解酶,同时也是有机磷和氨基甲酸酯类杀虫剂的靶标酶(冷欣夫等,1996),GST可催化内源性谷光甘肽与化学杀虫剂中具有毒理作用的亲电基团结合而排出体外(周奋启等,2011),对于烟粉虱抗性也有重要作用(杨秀清等,2001)。本研究中,带毒烟粉虱体内CarE活性明显低于无毒烟粉虱,GST和AChE活性也有所降低,但差异不显著,这跟烟粉虱取食感染TYLCV的番茄和感染TYLCCNV的烟草NicotianatabacumL.后解毒酶活性降低的研究结果一致(Luanetal., 2013;曹增等,2015)。烟粉虱取食TYLCV侵染的番茄后解毒酶活性被诱导,但30 d后CarE和GST活性降低(曹增等,2015),推断烟粉虱可能通过降低解毒酶活性减少能量消耗而增加在寄主植物上的适合度(Luanetal., 2013)。烟粉虱CarE和GST活性随寄主嗜好性而改变,在嗜好寄主上酶活性降低,在非嗜好寄主上升高,CarE和GST活性在一定程度上可作为烟粉虱对寄主适应性的参考指标(周福才等,2010),因此本研究中带毒Q型烟粉虱CarE活性降低说明Q型烟粉虱对ToCV侵染的番茄适应性较好。
猜你喜欢
上海蔬菜(2022年6期)2022-12-31
广西林业科学(2022年1期)2022-03-03
湖北农业科学(2021年22期)2021-12-13
湖北农业科学(2020年17期)2020-11-09
农家科技下旬刊(2020年5期)2020-06-19
植物保护(2018年5期)2018-12-05
农家顾问(2016年5期)2016-05-14
延安大学学报(自然科学版)(2015年4期)2015-06-15
江苏农业科学(2014年3期)2014-07-16
现代农业科技(2009年19期)2009-03-20
