
不同需冷量枣品种枝条休眠期间内源激素含量变化
2021-10-20 06:26张川疆杨植林敏娟王振磊冯一峰
塔里木大学学报 2021年3期
张川疆 ,杨植,林敏娟*,王振磊,冯一峰
(1塔里木大学植物科学学院,新疆 阿拉尔 843300)
(2南疆特色果树高效优质栽培与深加工技术国家地方联合工程实验室,新疆 阿拉尔 843300)
枣(Ziziphus jujubaMill.)属鼠李科(Rhamnaceae)枣属(ZiziphusMill.),是我国最古老的果树之一。其种质资源丰富,栽培历史悠久,是我国重要的经济林树种和特色果树[1]。新疆
从2000年开始发展枣产业,由于新疆独特的光热资源造就了枣产品卓越的品质,种植面积与产量跃居全国首位[2]。
植物休眠和解除是复杂的生物过程,受植株本身内在细胞变化与外界环境共同调节[3]。通常认为:休眠受激素控制,休眠的起始,终止和调控以及休眠阶段的改变受激素调节[4]。目前,在甜樱桃[5]、桃[6]等落叶木本植物休眠期间芽体激素含量变化上研究较多,但结果并不完全一致,而在枣树上的研究较少,难以为设施枣栽培提供理论依据。温带地区冬季来临之前日照变短,会产生脱落酸,促使植物芽体进入休眠[7];ZHEN C等[8]通过外施脱落酸的方式,延迟了葡萄冬芽的萌发。但脱落酸并非引起休眠的唯一因子,脱落酸与赤霉素、细胞分裂素间有拮抗作用,休眠现象或休眠解除与激素平衡有关。一般认为赤霉素是促进休眠解除的物质,低温促进赤霉素合成或去掉了阻止赤霉素合成的抑制物质,但一般认为赤霉素不是解除休眠的必需因素,而是芽生长的必需因素[9]。张文娟等[10]、吕长平等[11]在芍药自然休眠后期喷施赤霉素能有效打破休眠,促进萌发。细胞分裂素能克服存在于芽内阻止萌芽的抑制因子,外源喷施细胞分裂素可使芽内水由束缚态转变为自由态,通过和脱落酸之间的相互作用促进萌芽[12],于敏等[13]在种子休眠与萌发试验中也证实了这一观点。本试验通过对不同需冷量枣品种休眠期间内源激素含量变化进行了研究,旨在为人工调控休眠提供理论依据,对于枣促设施栽培具有重要的理论意义与实践价值。
1 材料与方法
1.1 试验材料
以塔里木大学园艺实验站枣种质资源圃的京枣39、伏脆蜜、胎里红为试验材料,其中京枣39需冷量为265 h,伏脆蜜需冷量为489 h,胎里红需冷量为620 h[14]。
1.2 试验方法
1.2.1 试验处理
自然越冬条件下:2020年10月15日到2021年1月14日自然越冬条件下每7 d采样1次,每次取枣头中部10~15 cm的枝条10根。用自来水清洗3次,去离子水清洗2次,充分去除枣树枝条表面的蜡质层,吸去枝条表面水分,用修枝剪剪成1~2 mm圆片,在液氮处理下在低温高速粉碎机上打成粉末,用于内源激素含量的测定。
人工低温处理条件下:2020年10月15日每品种采150根枝条,清洗枝条后用蜡将剪口封住。10根1组装入纸袋,放入4℃冰箱中,以自然休眠需冷量为中心前后共设置15个低温处理时数,京枣39在低温处理时数达92 h时,伏脆蜜在低温处理时数达240 h,胎里红在低温处理时数达528 h时,每隔24 h取出1袋,打成粉末,用于内源激素含量的测定。
1.2.2 内源激素含量的测定
称取样品1.0 g放入50 mL离心管,加入10 mL pH 2.2的盐酸-异丙醇提取液,冰浴振荡30 min,加入20 mL二氯甲烷,冰浴振荡30 min后,4℃4500 r/min离心35 min,取下层清液25 mL转入50 mL离心管中,通入氮气在低温冰浴氮吹仪下将清液吹干至沉淀,沉淀用2 mL 0.2%甲醇-甲酸溶液溶解后过膜转入色谱进样瓶中,用于高效液相色谱仪测定激素含量。
激素含量计算公式为:
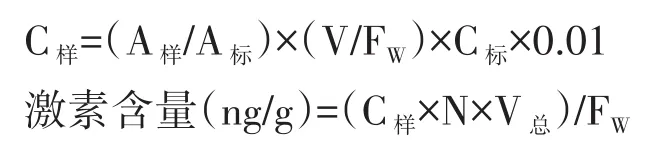
式中A样:样本峰面积;A标:标准品峰面积;V:提取液体积;FW:样本鲜重;C标:激素标准品浓度;N:稀释倍数;V总:沉淀溶解液体积。
1.3 数据统计与分析
试验数据用IBM SPSS Statistics 25、DPS 7.05软件进行处理。
2 结果与分析
2.1 不同需冷量枣品种枝条休眠期间赤霉素含量变化
2.1.1 自然越冬条件下不同需冷量枣品种枝条赤霉素含量变化
从图1中可以看出,自然越冬条件下3个枣品种休眠期间赤霉素含量在休眠诱导期变化幅度较大,且均在10月29日含量急剧下降,进入休眠后赤霉素含量变化不大,休眠解除后赤霉素含量变化幅度增大。

图1 自然越冬条件下不同需冷量枣品种枝条赤霉素含量变化
2.1.2 人工低温处理条件下不同需冷量枣品种枝条赤霉素含量变化
从图2、图3、图4中可以得知,人工处理条件下3个枣品种休眠期间赤霉素含量变化也各不相同。其中京枣39在休眠初期赤霉素含量逐渐上升,进入深度休眠后含量迅速下降,解除休眠前赤霉素含量迅速上升之后随着休眠的解除,赤霉素含量也随之降低;胎里红在休眠初期赤霉素含量变化不大,进入深度休眠后含量迅速上升,解除休眠后含量迅速下降;伏脆蜜在休眠初期赤霉素含量逐渐下降,进入深度休眠后含量呈梯度增加,当低温处理时数达408 h时,赤霉素含量迅速下降,随着休眠的解除,赤霉素含量也随之呈线性增加。
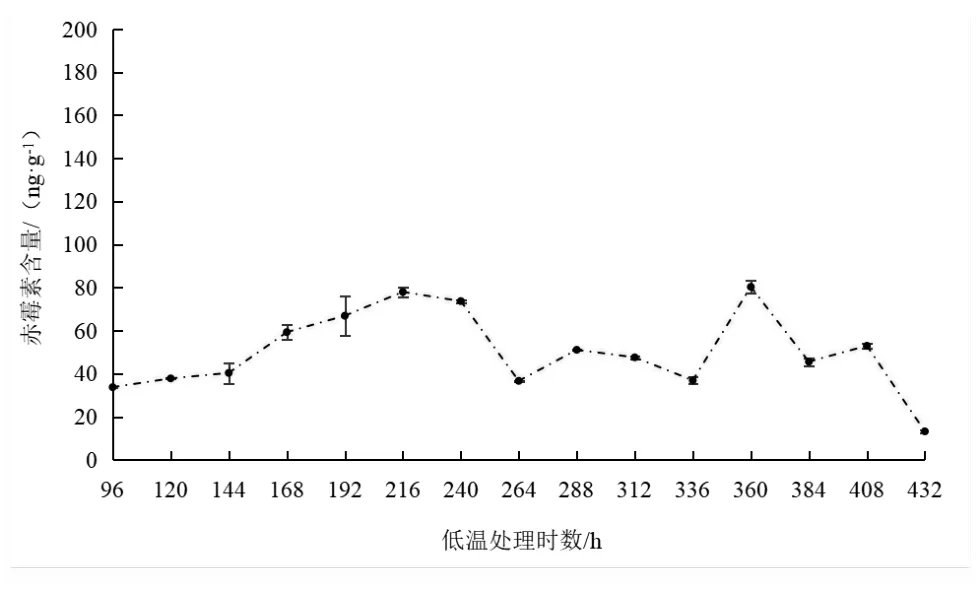
图2 人工低温处理条件下京枣39枝条赤霉素含量变化
2.2 不同需冷量枣品种枝条休眠期间生长素含量变化
2.2.1 自然越冬条件下不同需冷量枣品种枝条生长素含量变化
从图5中可以看出,自然越冬条件下胎里红和伏脆蜜2个枣品种休眠期间生长素含量变化差异性不大,总体呈现下降的趋势,变化幅度不大;但京枣39在休眠诱导期生长素含量呈下降趋势,进入休眠后生长素含量迅速上升,在解除休眠时达到峰值,解除休眠后含量迅速下降后开始回升。

图5 自然越冬条件下不同需冷量枣品种枝条生长素含量变化
2.2.2 人工低温处理条件下不同需冷量枣品种枝条生长素含量变化
从图6、图7、图8可知,人工处理条件下,3个枣品种休眠期间生长素含量变化均不相同。其中京枣39在整个休眠过程中生长素含量变化幅度较大;胎里红在整个休眠进程中生长素含量变化不明显,解除休眠时生长素含量出现了骤升骤降现象;伏脆蜜在整个休眠进程中生长素含量总体呈上升趋势。

图6 人工低温处理条件下京枣39枝条生长素含量变化

图7 人工低温处理条件下胎里红枝条生长素含量变化

图8 人工低温处理条件下伏脆蜜枝条生长素含量变化
2.3 不同需冷量枣品种枝条休眠期间脱落酸含量变化
2.3.1 自然越冬条件下不同需冷量枣品种枝条脱落酸含量变化
从图9中可以看出,休眠期间3个枣品种脱落酸含量呈现上升-下降-上升的趋势,且与自然休眠进程密切相关,在未进入休眠时脱落酸含量呈现上升趋势,随着休眠进程的持续进行,脱落酸含量开始逐步下降,当休眠解除后,脱落酸含量开始上升,在整个休眠过程中京枣39脱落酸含量显著高于其它2个枣品种,且峰值和谷值正对应着该品种进入和解除休眠的关键节点位置。其中京枣39脱落酸含量在11月19日达到峰值40.81 ng/g,较未进入休眠诱导期含量累计升高约8.48倍,在12月24日下降至谷值4.67 ng/g,较休眠初期含量累计下降了88.56%;胎里红脱落酸在11月19日达到峰值29.67 ng/g,较未进入休眠诱导期含量累计升高约7.05倍,在1月7日下降至谷值3.96 ng/g,较休眠初期含量累计下降了86.66%;伏脆蜜脱落酸含量在11月19日达到峰值28.93 ng/g,较未进入休眠诱导期含量累计升高约5.22倍,在12月31日下降至谷值5.91 ng/g,较休眠初期含量累计下降了79.57%,但总体脱落酸含量变化幅度不大。

图9 自然越冬条件下不同需冷量枣品种枝条脱落酸含量变化
2.3.2 人工低温处理条件下不同需冷量枣品种枝条脱落酸含量变化
由图10、图11、图12可知,人工处理条件下不同需冷量枣品种脱落酸含量变化呈上升-下降-上升的趋势,与自然休眠进程脱落酸含量变化趋势相同,但幅度和达到峰值、谷值的时间不同。其中京枣39低温处理时数达192 h时达到峰值24.99 ng/g,随着休眠的持续进行脱落酸含量持续下降,在低温处理时数达384 h下降至谷值4.66 ng/g,与该品种自然休眠需冷量一致;胎里红低温处理时数达624 h时达峰值21.89 ng/g,随后进入深度休眠,脱落酸含量开始下降,直至低温处理时数达816 h时下降至谷值15.61 ng/g,随着休眠的解除,脱落酸含量也随着开始上升;伏脆蜜低温处理时数达360 h时达到峰值43.13 ng/g,随后开始进入深度休眠,脱落酸含量也开始下降,直至低温处理时数达528 h时下降至谷值13.12 ng/g,之后脱落酸含量也开始上升。

图10 低温处理条件下京枣39枝条脱落酸含量变化

图11 低温处理条件下胎里红枝条脱落酸含量变化

图12 低温处理条件下伏脆蜜枝条脱落酸含量变化
3 讨论
通常认为果树休眠时由内源激素控制,休眠的起始、终止和调控以及休眠阶段的改变均受激素调节[15-16],不同需冷量枣品种休眠期间内源激素含量的变化较大。赤霉素与落叶果树芽休眠和萌发有着十分密切的关系。本试验发现枣树休眠诱导期植株体内有大量赤霉素合成,进入休眠后赤霉素含量迅速下降,深度休眠期赤霉素含量维持相对较低的水平,与赵晓菊等[17]在长春花、马海燕等[18]在葡萄上的研究结果一致,与葛会波等[19]、申小丽等[20]在草莓上的研究结果相反。
整个休眠期生长素含量的变化不大,休眠解除后其含量有迅速上升,生长素含量的变化与枣树休眠进程联系不紧密,与朱永亮等[21]在杨树上的研究结果一致。
脱落酸含量的变化与枣休眠进程关系密切。脱落酸是促进休眠和抑制植株花芽分化的重要物质[22]。潘根生等[23]在茶树、SUBBAIAH T K等[22]在苹果上的研究发现,植物进入休眠后脱落酸含量持续下降,解除休眠时脱落酸含量并未下降,相反,在春季开始生长时,脱落酸含量缓步上升。本研究也得到了一致的结论,脱落酸含量在休眠诱导期随着休眠进程的持续推进而上升,进入休眠后脱落酸含量持续下降,解除休眠后脱落酸含量开始缓慢升高。
4 结论
脱落酸含量的变化与枣品种休眠进程有着密切的关系。其含量在休眠诱导期随着休眠进程的持续推进而上升,进入深度休眠后脱落酸含量也相应减低,解除休眠后脱落酸含量开始缓慢升高。人工处理条件下,各品种脱落酸含量变化与自然休眠过程中产生的结论一致。因此可以把脱落酸含量变化看作枣树休眠进程中的一个关键性指标。
猜你喜欢
落叶果树(2022年3期)2022-06-07
山西农业科学(2021年11期)2021-12-17
军事文摘(2021年16期)2021-11-05
小天使·六年级语数英综合(2020年3期)2020-12-15
三农资讯半月报(2020年14期)2020-08-09
小天使·四年级语数英综合(2019年10期)2019-11-09
劳动保护(2018年8期)2018-09-12
中学生物学(2018年1期)2018-01-19
山西果树(2014年3期)2014-07-15
果农之友(2009年8期)2009-09-11
