
小麦茎秆强度关联位点及优异等位变异分析
2022-01-06 03:07田胜明司红起马传喜
麦类作物学报 2021年11期
卢 杰 ,田胜明,王 胜,陈 璨,司红起,常 成,马传喜
(安徽农业大学农学院/农业部黄淮南部小麦生物学与遗传育种重点实验室,安徽合肥 230036)
随着小麦产量的提高,其倒伏的风险也增大[1],小麦倒伏导致机械收割成本增加以及籽粒品质和商品性变差[2]。小麦生产中,茎秆倒伏是最常见的倒伏类型,主要发生在基部节间,与株高和茎秆强度显著相关[3]。矮化育种使育成品种株高降低,抗倒性增强;但育种实践表明,过分强调矮杆将会造成地上部生物量下降,最终影响产量,且矮化后的植株若茎秆质量较差仍会出现倒伏现象[4]。因而,育种实践上小麦品种的抗倒伏策略应由原来的矮秆抗倒转变为壮秆抗倒,注重茎秆质量的改善与提高[1,3,5]。因此,研究小麦茎秆倒伏的遗传机理,将有助于抗倒伏品种的培育。
确定合适的易倒测定时期及倒伏评价指标是进行小麦抗倒伏研究的关键。研究发现,在乳熟期,小麦物质的生产和运输比较旺盛,茎秆中大量结构物质被分解和转移,是极易发生倒伏的时期[6]。抗倒伏指数[4]、茎秆断裂力[7]、茎秆弹性[8]、田间自然倒伏[9]、茎秆强度[10]等可以用来评价小麦抗倒伏性。其中,田间自然倒伏观察是小麦生产上最直接和常用的评价方法,但该方法易受环境因素制约,重复性差。采用抗倒伏指数和茎秆断裂力两个指标时,测量过程中需破坏茎秆。而茎秆强度评价指标具有不破坏小麦茎秆、重复性好等优势,且与小麦的抗倒伏性显著正相关[10],是较理想的抗倒伏性评价指标之一。
研究发现,除株高外,小麦茎秆基部的结构特征(节间长度、粗度和壁厚[10-12]以及维管束种类和数目[7,10,13])、木质素、纤维素和全糖含量[13-19]等均与小麦的抗倒伏性相关。国内外的研究学者对小麦抗倒伏遗传机理进行了研究,Ma等[15-18]发现,木质素含量与小麦抗倒伏性关系密切,并克隆了3个小麦木质素合成基因Ta-CCR1、Ta-COMT和Ta-CAD1。此外,一些与小麦茎秆抗倒伏性相关的QTL也有报道,其中,控制茎秆硬度的QTL位于3B和4D染色体上[20-21],控制基部第1节间长度的QTL位于2D和 4B染色体上[22],控制髓腔直径的QTL位于1A和2D染色体上[23],控制茎粗的QTL位于3B染色体上[23],控制茎壁厚的QTL位于2D和3BL染色体上[23-24],控制茎秆强度的QTL位于1B、2A、3A、3B、4A、4D、5A、5B和5D染色体上[23,25-26]。上述研究表明,小麦抗倒伏相关性状受多基因控制。可见,进一步挖掘与小麦茎秆强度相关的基因或QTL,开发可用于育种选择的分子标记或优异等位变异,将有助于小麦抗倒伏性的遗传改良。
本研究以126份小麦种质为研究材料,基于三个环境(2011-2012、2012-2013和2013-2014年度)下的乳熟期茎秆强度表型值,利用卢 杰等[27]前期筛选的242对SSR标记对其进行基因分型,应用混合线性模型(mixed linear model,MLM)检测与小麦乳熟期茎秆强度紧密关联的标记位点,发掘相关位点的优异等位变异,以期为小麦茎秆强度有关基因的克隆和抗倒伏分子育种提供参考。
1 材料与方法
1.1 试验材料
126份小麦种质材料为新审定或新选育的中间品系,其中,黄淮南片麦区77份,黄淮北片麦区18份,长江中下游麦区17份,西南麦区10份,国外种质4份。试验分别于2011-2012、2012-2013和2013-2014年度种植于合肥高新技术农业示范园(31.9′N,117.2′E)内,2行区,行距0.25 m,行长2 m,2次重复,常规管理。
1.2 茎秆强度的测定
参照肖世和等[10]的方法,于小麦花后28 d(乳熟期),使用茎秆强度仪(YYD-1A型)测定供试材料的茎秆强度,将茎秆强度仪显示的峰值作为供试材料的茎秆强度值,茎秆强度值是五个单茎捆扎在一起的实测值,进行3次重复。
1.3 分子标记的筛选及其扩增条件
利用242对多态性丰富且带型清晰的SSR引物[27]用于后续分析。采用SDS法提取小麦基因组DNA。PCR扩增体系为10 μL,包括模板DNA(50 ng·μL-1)2.0 μL,10×Buffer(含有 2.0 mmol·L-1Mg2+)1.0 μL,dNTPs(2.5 mmol·L-1) 0.8 μL,Taq DNA polymerase(5.0 μ·μL-1) 0.11 μL,上下游引物(10 μmol·L-1)各0.4 μL,ddH2O 5.29 μL。PCR扩增程序:94 ℃预变性5 min;95 ℃变性30 s,48~62 ℃退火30 s,72 ℃延伸30 s,共35个循环;72 ℃延伸10 min;4 ℃保存。利用高通量Tiling分析系统即毛细管电泳对PCR产物进行检测。
1.4 关联分析及优异等位变异发掘
参考Mackay等[28]的方法,对试验材料进行遗传结构分析。运用Structure软件[29]混合模型来确定群体结构,K值取1~10,单独运行6次,依据该运行结果选择适当的亚群数。
利用TASSLE软件[30]中的MLM,将群体结构分析所得的Q值和亲缘关系K作为协变量,进行性状与标记位点的关联分析,统计显著关联标记位点对表型变异的解释率。参考文自翔等[31]的无效等位变异估算方法,计算与茎秆强度显著关联标记位点等位变异的表型效应,进一步挖掘其优异等位变异及其载体材料。
2 结果与分析
2.1 供试材料茎秆强度的表型变异
从表1可以看出,2011-2012、2012-2013和2013-2014三个年度供试材料的茎秆强度变化范围分别为0.51~3.07、0.51~3.54和 0.57~ 2.56 N,变异系数分别为34.0、27.7和26.7,说明供试材料的表型变异丰富。方差分析结果(表2)显示,小麦茎秆强度受环境和基因型共同影响,且受环境的影响较大。进一步对三个年度的茎秆强度值进行相关分析,结果显示,三个年度间的茎秆强度值互相均呈极显著正相关,其中2011-2012年度与2012-2013年度间的相关系数为0.453,2011-2012年度与2013-2014年度间的相关系数为0.416,2012-2013年度与2013-2014年度间的相关系数为 0.428。综上分析表明,小麦茎秆强度易受环境因素影响,但在不同年度间,不同试验材料的茎秆强度表现出相对稳定的趋势。

表1 供试种质材料茎秆强度的表型变异Table 1 Phenotype variation of stem strength of the tested wheat germplasm materials

表2 供试126份种质材料茎秆强度的方差分析Table 2 ANOVA of stem strength in 126 wheat germplasm materials
2.2 群体结构分析结果
利用Structure软件对可能的K值模拟计算,利用K值与△K值作图(图1A),发现在K=2时△K值最大。由图1B可以看出,126份种质材料被分为2个亚群,其中第1亚群(POP1)主要来自西南麦区和长江中下游麦区,共44份材料;第2亚群(POP2)主要来自黄淮南片麦区和黄淮北片麦区,共82份材料,说明这126份种质材料的群体结构较为简单,可有效降低群体结构对关联分析的影响。K=2时生成的Q矩阵被用于茎秆强度性状与标记位点的关联分析。

A:假定K值范围从1到10计算超过6次的△K值;B:基于模型的126份小麦种质材料的群体结构。A:Calculated △K value over six runs with putative K ranging from 1 to 10; B:Model-based population structure for 126 wheat germplasm.图1 126份小麦种质材料的群体结构分析Fig.1 Population structure analysis of the 126 wheat germplasm materials
2.3 茎秆强度相关分子标记的鉴定结果
利用TASSLE软件中的MLM对126份自然群体的基因型及不同年度的茎秆强度进行关联分析,结果(表3)表明,在2个或2个以上年度重复检测到的显著关联位点共37个,分布于1A(2个)、1B(1个)、2A(1个)、2B(5个)、2D(2个)、3B(1个)、3D(3个)、4A(3个)、4B(1个)、4D(2个)、5A(4个)、5B(3个)、5D(2个)、6A(1个)、6B(3个)、7D(3个)等16条染色体上。2011-2012、2012-2013和2013-2014三个年度与茎秆强度显著关联的标记位点分别有28、29和25个,表型变异解释率均大于10%的标记位点分别有6、13和9个。有8个标记位点在三个年度均能被检测到,分别为wmc83、gwm539、wmc48、barc358、barc59、barc322、barc134和barc352,属于稳定 位点。

表3 与茎秆强度显著关联的标记位点及其表型变异解释率Table 3 Phenotype contribution of marker loci significantly associated with stem strength %
2.4 优异等位变异分析结果
选取分布在2B、2D、5A、5B和6B染色体上表型解释率在8%以上的5个稳定位点(wmc83、gwm539、barc358、barc59 和barc134 )进行等位变异分析,结果(图2)表明,共发掘11个在三个年度茎秆强度均增加的优异等位变异,分别为wmc83-A110、wmc83-A147、wmc83-A151、gwm539-A120、barc358-A179/161、barc358-A185/161、barc358-A185/179、barc358-A190/161、barc59-A182、barc59-A191和barc134-A194,其中barc134-A194等位变异的分布频率最高(39.7%),wmc83-A110的增效效应最大,平均为0.80 N。此外,wmc83-Miss、wmc83-A155、gwm539-A125、gwm539-A152、barc358-A161、barc358-A179、barc358-A190/185、barc59-A167、barc134-A186、barc134-A191和barc134-A192等位变异在三个年度均降低茎秆强度,其中wmc83-A155等位变异的分布频率最高 (34.1%),barc358-A161的减效效应最大,平均为-0.66 N。
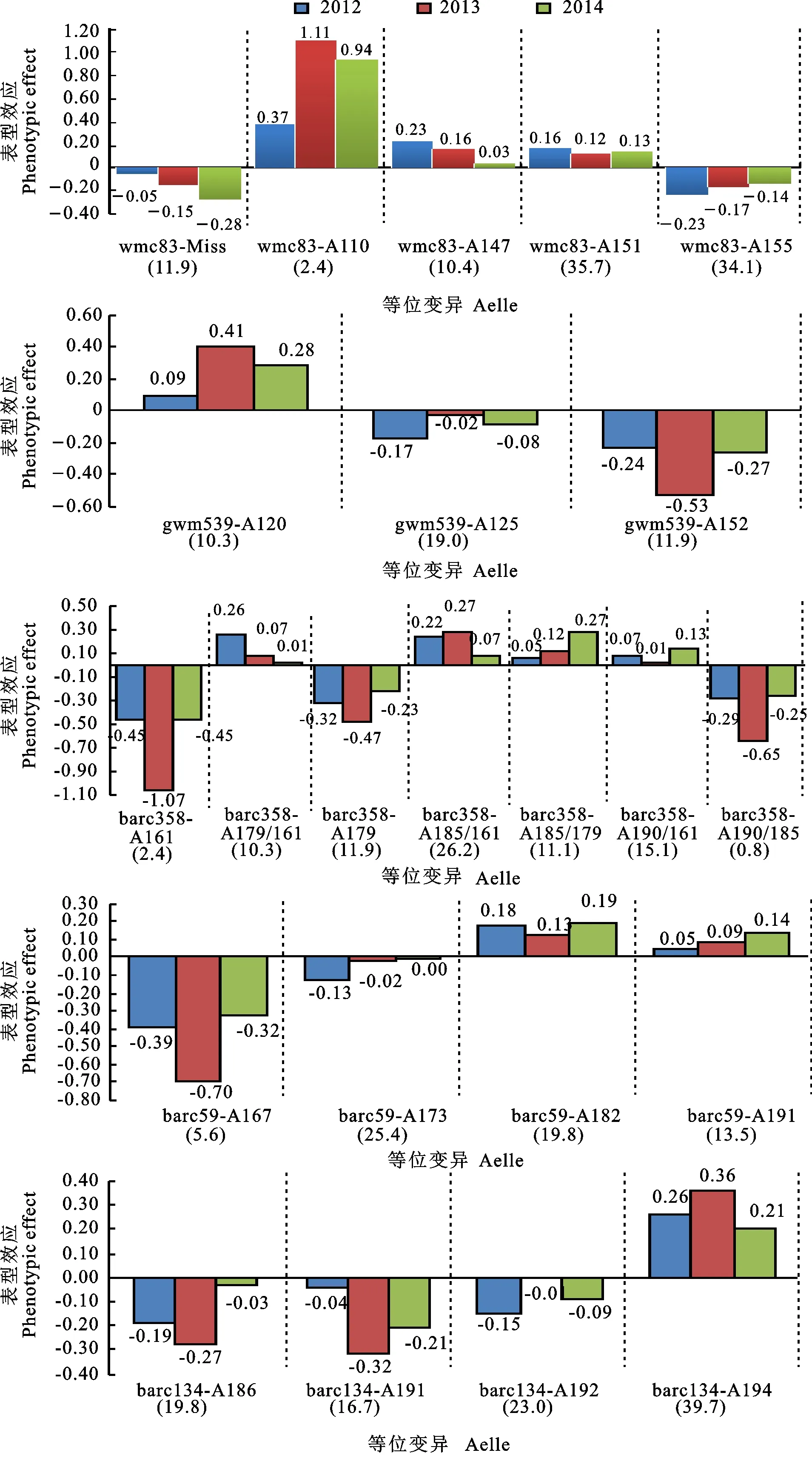
括号内数据为携带该位点品种的比例,单位为%。Values in brackets represent the percentage of varieties carrying the associated locus,and the unit is %.图2 5个与茎秆强度显著关联位点等位变异的表型效应Fig.2 Phenotypic effect of allelic variation of the five loci which significantly associated with stem strength
统计分析上述11种优异等位变异在供试材料中的分布,结果(表4)显示,共116份种质材料携带上述优异等位变异,其中携带5、4、3、2和1种优异等位变异的材料分别有2、12、21、42和39份,三个年度茎秆强度平均值分别为2.65、2.04、1.80、1.70和1.49 N;未携带上述优异等位变异的材料有10份,平均茎秆强度为1.21 N。在122份国内材料中(表5和表6),黄淮南片麦区、黄淮北片麦区、长江中下游麦区和西南麦区的种质材料中携带2(40.3%)、1(27.8%)、1 (35.3%)和4(50.0%)种优异等位变异的分布频率最高;仅西南麦区发现有聚合5种优异等位变异的材料(渝0926和黔079984-14)。此外,携带4种及以上优异等位变异的材料中,西南麦区材料中分布频率最高(70.0%),明显高于其他三个麦区。总的来说,来自西南麦区的重要载体材料有内麦10号、内麦8号、黔079984-14、内麦11号和渝0926,来自黄淮南片及北片麦区的重要载体材料有中麦1095、豫农69、安农0711和漯6099(表6),这些载体材料可作为小麦茎秆强度遗传改良改良的种质资源。
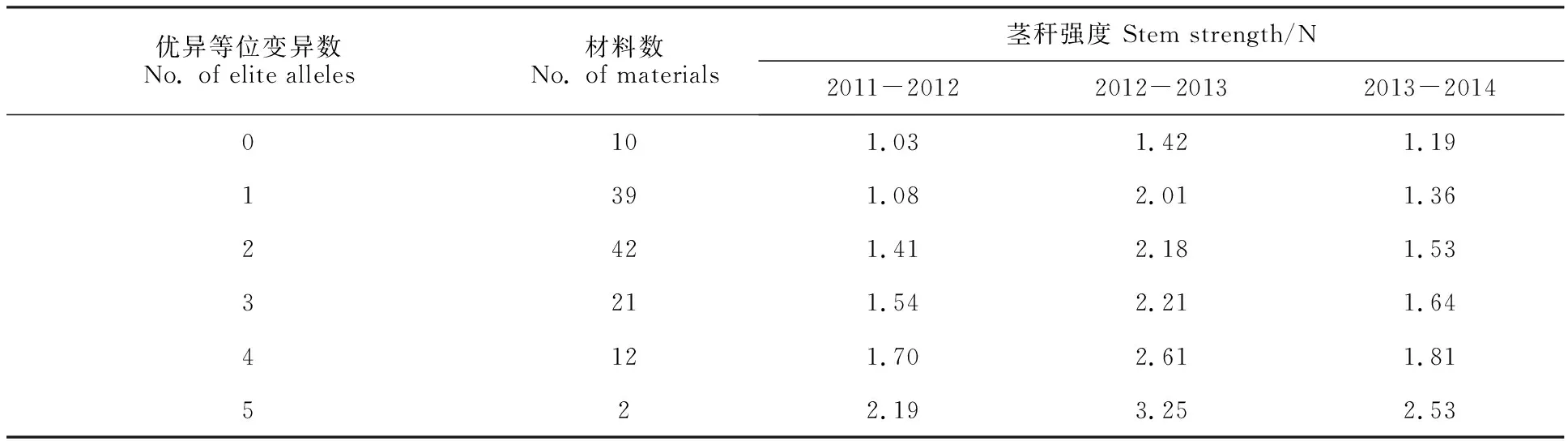
表4 载体材料优异等位变异分布及其茎秆强度Table 4 Distribution of elite alleles and stem strength of carrier materials

表5 不同麦区种质材料中携带优异等位变异数量的分布频率Table 5 Distribution frequency of elite alleles in experimental materials from different wheat regions
3 讨 论
本研究共检测到37个与小麦茎秆强度显著相关的位点,其中在多环境下稳定检测到的位点有8个,表明小麦茎秆强度是多基因控制的数量性状,易受环境因素影响,与前人的研究结果一致[23,25-26]。本研究在5A和5B染色体上检测到两个稳定位点,分别为barc358和barc59,Keller等[26]在5A和5B条染色体上也检测到与茎秆强度相关的QTL,但由于所用的遗传标记不同,两个研究中鉴定到的关联位点是否相同,有待进一步分析验证。
多年多点的表型数据是提高关联定位准确性的重要策略,多个环境下均能检测到的位点具有较高的可靠性[32],具有一定的分析利用价值。本研究中有5个位点(wmc83、gwm539、barc358、barc59和barc134)在多个环境条件下表现稳定,表型解释率高,共包含11种优异等位变异,且载体材料的茎秆强度随着携带优异等位类型数目的增多而增大,表现出一定的聚合效应。因此,在小麦抗倒伏遗传改良中,可通过聚合茎秆强度优异等位变异而提高茎秆质量。
本研究发现,不同麦区的试验材料中优异等位变异分布差别较大。来自西南麦区的材料聚合较多的优异等位变异,茎秆强度较高,推测与本麦区独特的大穗大粒品种类型有关,大部分该类品种茎秆粗壮,具有较好的茎秆质量和强度;其中重要的载体材料有内麦10号、内麦8号、黔079984-14、内麦11号和渝0926,三个年度其平均茎秆强度分别为2.68、2.64、2.63、2.43和2.09 N。本研究中增效效应最大的优异等位变异wmc83-A110分布在内麦10号和黔079984-14材料中,这些载体材料可作为本区域小麦茎秆强度改良的重要资源。Zhang等[1,3]研究发现,60年代以来黄淮麦区主推小麦品种的茎秆强度呈显著增长趋势,表明在小麦株高处于适宜范围时,本区域小麦抗倒伏育种目标已从降低株高转至增加茎秆强度。今后黄淮麦区小麦抗倒伏育种中仍应以改良茎秆强度为主要育种目标,聚合更多茎秆强度优异等位变异,达到提高茎秆强度的目的。相比西南麦区,黄淮南片和北片麦区试验材料中聚合4种优异等位变异的分布频率偏低,但本研究亦鉴定出中麦1095、豫农69、安农0711和漯6099共4份重要的载体材料,三个年度的平均茎秆强度分别为2.67、2.31、2.12和2.10 N;此外,上述试验材料除聚合较多优异等位变异外,还具有较好的经济和农艺性状,可作为本区域小麦茎秆强度改良的种质资源。
4 结 论
本研究基于混合线性模型(MLM),对多环境下126份小麦种质材料的茎秆强度进行关联分析,得到5个稳定标记位点,发掘出11种优异等位变异,且载体材料茎秆强度随优异等位变异聚合数目的增多而增大。内麦10号、内麦8号、黔079984-14、内麦11号、渝0926、中麦1095、豫农69、安农0711和漯6099共9份材料聚合了4种及以上优异等位变异,且茎秆强度较高,可作为相应麦区小麦茎秆强度遗传改良的种质资源。
猜你喜欢
分子催化(2022年1期)2022-11-02
今日农业(2022年14期)2022-09-15
今日农业(2022年13期)2022-09-15
江西农业大学学报(2022年3期)2022-07-07
中国种业(2022年1期)2022-01-27
中国种业(2022年1期)2022-01-27
中国生态农业学报(中英文)(2021年9期)2021-09-10
烟草科技(2021年6期)2021-06-24
生物学教学(2018年4期)2018-11-29
电脑知识与技术(2018年19期)2018-11-01
