
基于溶藻弧菌感染的合浦珠母贝转录组数据分析IAP蛋白家族的表达变化
2022-05-17 07:59蒋美君钟方杰喻达辉张斌严雪瑜
农业与技术 2022年9期
蒋美君钟方杰喻达辉张斌严雪瑜
(1.广西北部湾海洋生物多样性养护重点实验室/北部湾大学海洋学院,广西 钦州 535011;2.钦州市水产技术推广站,广西 钦州 535099)
合浦珠母贝(Pinctada fucata)又称马氏珠母贝,是中国南海海域主要培育的海水珍珠贝,在我国海水养殖业中占有重要地位[1]。近年来,随着珍珠养殖规模不断扩大,种质资源退化和病害问题日趋严重,海水珍珠产量和质量明显下降,严重威胁我国海水珍珠产业的持续健康发展[2]。合浦珠母贝仅具备非特异性免疫系统,极易被病原体和微生物侵染[3],提高合浦珠母贝自身免疫防御机制研究是如今重要的发展方向[4,5]。
溶藻弧菌(Vibrio alginolyticus)是一种嗜盐性革兰氏阴性短杆菌,主要生长在海洋或出海口,数量位于海水类弧菌之首,易引起鱼、虾、贝等海洋生物爆发流行性疾病[6,7]。已有研究发现,溶藻弧菌给石斑鱼(Epinephelus coioides)[8]、凡纳滨对虾(Litopenaeus vannamei)[9]、近江牡蛎(Ostrea rivularis Gould)[10]等养殖产业造成了严重的经济损失。Wang等[11]利用转录组测序技术分析合浦珠母贝响应溶藻弧菌感染的免疫机制,发现有636个差异表达基因响应了弧菌攻击,这些基因主要参与了丝裂原活化蛋白激酶信号通路(MAPK signaling pathway)、趋化因子信号通路(chemokine signaling pathway)、细胞凋亡(Apoptosis)和Wnt信号通路(Wnt signaling pathway)等免疫相关信号通路。本研究通过利用转录组测序技术开展合浦珠母贝在溶藻弧菌早期感染下的分子免疫调控机制研究,并筛选出凋亡抑制因子(inhibitor of apoptosis proteins,IAPs)蛋白家族中的几个关键差异表达基因进行基因表达变化分析,以期寻找和探索合浦珠母贝先天免疫调控机制的关键候选基因,为提高合浦珠母贝的非特异性免疫能力和培育具有优良性状的新品种提供理论基础[12]。
1 材料与方法
1.1 试验材料
试验用的1龄健康合浦珠母贝来源于广西北海的珍珠贝养殖场,在实验室条件下暂养7d后进行溶藻弧菌感染试验。在暂养和试验期间,每天定时投喂藻类。
1.2 试验方法
1.2.1 溶藻弧菌悬液制备
将冻存的溶藻弧菌用牛肉膏蛋白胨固体培养基进行划线分离活化2次,取活化的溶藻弧菌接种于牛肉膏蛋白胨液体培养基中,在28℃下200r·min-1振荡过夜培养,1600r·min-1离心10min后弃上清液,重新悬浮于PBS溶液,并配制成浓度为4.0 OD的具有活性的溶藻弧菌悬液,4℃保存备用。
1.2.2 溶藻弧菌感染试验及转录组测序
将合浦珠母贝用氯化镁溶液进行麻醉开壳后,于闭壳肌注射100μL制备好的弧菌悬液;采集注射前0h和注射后6h和24h的合浦珠母贝肝胰腺组织样品,每组各采集6个贝,所采集的组织样品置于液氮中冻存后置于-80℃冰箱中待用。
1.2.3 转录组测序及IAP蛋白家族基因表达分析
将合浦珠母贝肝胰腺组织样品在干冰保存的条件下送至中国上海美吉生物技术公司进行cDNA文库构建和mRNA测序;再对转录组测序获得的IAP蛋白家族中的基因按照p-adjust<0.001和|log2FC|≥1的标准进行差异表达分析,并利用KEGG(Kyoto Encyclopedia of Genes and Genomes)数据库对差异表达基因进行功能注释。
2 结果与分析
2.1 合浦珠母贝BIRC2基因在溶藻弧菌感染下的表达变化
转录组结果显示,在感染溶藻弧菌后6h时,合浦珠母贝BIRC2基因的表达量显著上升(P<0.05);而在24h时表达量下降,但仍显著高于感染前(0h)的表达量(P<0.05),结果如图1所示。基因功能分析结果显示,BIRC2基因富集到了KEGG中的NOD样受体信号通路(NOD-like receptor signaling pathway)、Toll和IMD信号通路(Toll and Imd signaling pathway)以及弓形虫病(Toxoplasmosis)等信号通路。
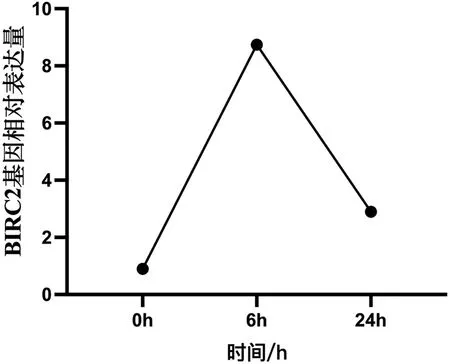
图1 合浦珠母贝BIRC2基因在感染溶藻弧菌不同时间中的表达变化
2.2 合浦珠母贝BIRC3基因在溶藻弧菌感染下的表达变化
如图2所示,在感染溶藻弧菌后6h时,合浦珠母贝BIRC3基因的表达量急剧上升,并显著高于感染前0h和感染后24h(P<0.05),而24h与0h的基因表达量差异不显著(P>0.05)。基因功能分析结果发现,BIRC3基因也富集到了NOD样受体信号通路(NOD-like receptor signaling pathway)、Toll和IMD信号通路(Toll and Imd signaling pathway)以及弓形虫病(Toxoplasmosis)等信号通路中。

图2 合浦珠母贝BIRC3基因在感染溶藻弧菌不同时间中的表达变化
2.3 合浦珠母贝BIRC4基因在溶藻弧菌感染下的表达变化
如图3所示,在感染溶藻弧菌后6h时,合浦珠母贝BIRC4基因的表达量较感染前(0h)有所增加,但与0h的表达量差异不显著(P>0.05);而在24h时,BIRC4基因的表达量明显下降,并显著低于6h(P<0.05),但与0h的表达量差异不显著(P>0.05)。基因功能分析结果发现,BIRC4基因被富集到了泛素介导的蛋白质水解(Ubiquitin mediated proteolysis)信号通路中。

图3 合浦珠母贝BIRC4基因在感染溶藻弧菌不同时间中的表达变化
2.4 合浦珠母贝BIRC7基因在溶藻弧菌感染下的表达变化
基因表达结果显示,在感染溶藻弧菌后6h时,合浦珠母贝BIRC7基因的表达量增加,并显著高于感染前0h和感染后24h(P<0.05);而24h与0h的基因表达量差异不显著(P>0.05),结果如图4所示。基因功能分析结果显示,BIRC7基因被富集到了弓形虫病信号通路中。
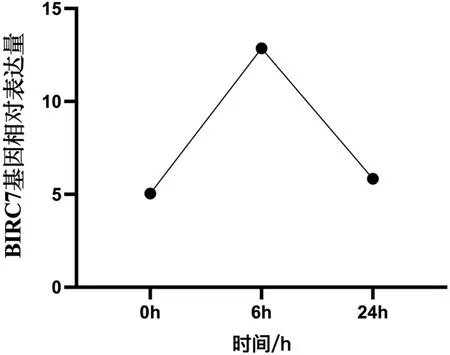
图4 合浦珠母贝BIRC7基因在感染溶藻弧菌不同时间中的表达变化
3 讨论
凋亡抑制因子(inhibitor of apoptosis proteins,IAPs)是一类具有抑制由病毒感染引起的细胞凋亡作用的内源性蛋白,大多数IAPs含有杆状病毒IAP重复区域(baculoviral IAP repeat containing,BIRC)[13]。1993年Crook等首先在杆状病毒CpGV和核多角体病毒OPMNPV中发现IAP家族[14],随后的研究发现人类细胞中存在7种IAPs,包括NAIP(BIRC1)、cIAP1(BIRC2)、cIAP2(BIRC3)、XIAP(BIRC4)、Survivin(BIRC5)、Apollon(BIRC6)和Livin(BIRC7)[15]。目前已在池蝶蚌(Hyriopsis schlegeli)[16]、太平洋牡蛎(Crassostrea gigas)[17]和中华绒螯蟹(Eriocheir sinensis)[18]组织中发现IAP分子能够抑制细胞凋亡。苗国英等[19]使用脂多糖(Lipopolysaccharide,LPS)模拟细菌感染对栉孔扇贝(Chlamys farreri)进行刺激,发现CfIAP-1和CfIAP-1基因表达量在0~12h没有明显变化,在24h迅速上升达到最高点,到48h开始下降,72h恢复到正常水平,整体呈现先升高后下降的趋势。Zhang等[20]利用鳗弧菌(Vibrio anguillarum)对太平洋牡蛎进行刺激,发现IAP基因表达量在0~12h逐渐升高,在12h时达到最高水平,然后从12~48h逐渐下降,也呈现出先升高后下降的趋势。
本研究共发现有4个IAP蛋白家族的基因在合浦珠母贝感染溶藻弧菌后表达变化显著,这4个基因的表达量均在感染期间(0~24h)呈现先升后降的趋势,结果与上述研究结果基本一致。其中,合浦珠母贝的BIRC2、BIRC3和BIRC7基因在感染6h时表达显著上调,并均与0h和24h差异显著(P<0.05);BIRC4基因的表达量也在感染6h时明显增加,并显著高于感染24h时的表达量(P<0.05),但与0h的表达量差异不显著(P>0.05)。通过基因功能分析发现,BIRC2、BIRC3基因富集到了KEGG中的NOD样受体信号通路、Toll和IMD信号通路以及弓形虫病等通路中;而BIRC4基因则富集到了泛素介导的蛋白质水解信号通路中,BIRC7基因富集到了弓形虫病信号通路中。
综上所述,本研究通过利用溶藻弧菌感染合浦珠母贝的转录组数据分析了IAP蛋白家族的基因表达变化,发现BIRC2、BIRC3、BIRC4和BIRC7基因的表达量发生显著变化,并发现这些基因被富集到了一些免疫相关的信号通路中,表明IAP参与了合浦珠母贝的免疫应答反应过程,提示其在贝类非特异性免疫调节中发挥着重要作用。
猜你喜欢
当代水产(2022年8期)2022-09-20
当代水产(2021年9期)2021-12-02
当代水产(2019年5期)2019-07-25
西南农业学报(2017年5期)2017-06-23
海洋渔业(2016年6期)2016-04-16
环境污染与防治(2016年9期)2016-03-13
微生物学杂志(2015年6期)2015-12-27
广东海洋大学学报(2015年3期)2015-12-22
地方文化研究(2015年4期)2015-11-07
大众考古(2015年7期)2015-06-26
