
茄子和托鲁巴姆ZIP5 和ZIP11 基因对锌/镉毒害的响应分析
2022-05-20 09:41吴非刘飞张苹李栋孙亮亮徐进
山西农业大学学报(自然科学版) 2022年2期
吴非,刘飞*,张苹,李栋,孙亮亮,徐进
(1.山西农业大学 园艺学院,山西 太谷 030801;2.中国农业大学 资源与环境学院,北京 100091)
托鲁巴姆(Solanum torvumSw.)是茄子(So⁃lanum melongenaL.)的近缘野生种,是一种优良的茄子砧木和重要的镉(Cd)低累积作物之一[1]。有研究表明,茄子嫁接到托鲁巴姆砧木上后,其果实中Cd 含量下降了63%~74%[2]。Cd 由根系向茎叶的长距离运输,即Cd 木质部装载量,是托鲁巴姆地上部Cd 低累积的关键因素[3]。另外,荧光探针检测表明,在茄子根中Cd 主要分布在中柱,而在托鲁巴姆根中Cd 主要分布于皮层[4]。镉通过铁(Fe)、钙(Ca)和锌(Zn)转运/通道蛋白进入植物细胞[5],高浓度Cd 处理后的托鲁巴姆转录组分析结果表明,ZIP 家族转运蛋白(Zinc and iron regulated transporter proteins)基因的差异表达在托鲁巴姆Cd 的低累积发生过程中可能发挥了重要作用[6],但具体机制还有待进一步的研究。
ZIP 金属转运蛋白包括Zinc regulated transporter proteins(ZRT)和Iron regulated transporter proteins(IRT)[7],ZIP 家族转运蛋白在植物中广泛存在,对Zn2+、Fe2+、Cu2+及Cd2+等多种二价金属离子的吸收、分配及运输起重要作用[8-10]。ZIP 蛋白家族成员发挥功能往往具有不同的转运对象及专一性,其表达水平也受到环境金属离子浓度的调控[11]。拟南芥AtIRT1是在根表层细胞中高度表达的Fe转运蛋白[12];AtZIP1⁃5、AtZIP9⁃12和AtIRT3在拟南芥根和茎中受缺Zn 诱导表达[13],并且AtZIP2和AtZIP5也在缺铜(Cu)诱导下于根部表 达[14]。水 稻OsIRT1和OsIRT2主要在根部表达,对Fe2+进 行的吸收和运输[15];缺Zn 胁迫 下,OsZIP2仅在根部表达,OsZIP7仅在地上部表达,而OsZIP1、OsZIP3⁃5和OsZIP8在根和地上部中均表达[16-19]。研究ZIPs 转运蛋白在不同植物中的表达模式,揭示不同金属离子胁迫下ZIPs基因在各组织部位中的表达规律,可为提高作物养分吸收效率提供新的研究方向[20-21]。本研究利用qRTPCR 方法,分析了托鲁巴姆StZIP5/11基因和茄子SmZIP5/11基因在Zn 和Cd 毒害下的表达模式,测定拟南芥突变体(zip5和zip11)以及转托鲁巴姆StZIP5和StZIP11基因拟南芥分别在Zn、Cd 毒害下主根生长量,为初步探明StZIP5和StZIP11基因功能提供理论依据。
1 材料与方法
1.1 试验材料
托鲁巴姆和茄子种子先用无菌水浸泡30 min,无菌环境下置于50% 消毒水中浸泡消毒8 min,再用无菌水冲洗5~8 遍。将种子均匀的播种在1/2MS 培养基表面。4 ℃中春化3 d 后,放至人工气候室中进行萌发培养。培养条件为22 ℃、5000 lx、16 h/8 h 光暗。待子叶完全展开后,选取长势均匀一致的托鲁巴姆和茄子幼苗,转移到1/2 Hoagland 营养液中进行培养,培养条件不变。每3 d 更换1次营养液。对生长4 周的托鲁巴姆和茄子幼苗,在营养液中分别进行添加200 μmol·L-1Zn或10 μmol·L-1Cd,每处理设3 组重复。以正常1/2 Hoagland 营养液培养条件下生长的幼苗为对照,分别取处理后第1、3 天的叶、茎和根为样品。样品收集后立即液氮冷冻并放于-80 ℃保存。
拟南芥突变体zip5(SALK_009007C)和zip11(SALK_009007C)购自TAIR(https://www.arabidopsis.org/)。将托鲁巴姆StZIP5和StZIP11基因的CDS 序列分别与含有CaMV 35S 启动子的pCAMBIA1300 载体连接,构建StZIP5和StZIP11基因过表达载体,转化农杆菌菌株。利用拟南芥花 序 浸 染 法[22],将构建好的StZIP5和StZIP11基因过表达载体转化到拟南芥中,并获得转StZIP5和StZIP11基因的拟南芥StZIP5OX、StZIP11OX、StZIP5OX/zip5、StZIP11OX/zip11、StZIP5OX/zip11和StZIP11OX/zip5后代。将获得的种子在1/2MS(hyg+)固体培养基上进行筛选;利用qRT-PCR 对转基因T3代后代进行基因表达鉴定,选择阳性株系进行后续试验(表1)。

表1 鉴定转基因拟南芥引物Table 1 Primers for identification of transgenic A.thaliana
1.2 生物信息学分析
分析所用的托鲁巴姆StZIP5和StZIP11的CDS 序列由作者在前期采用RACE 方法克隆得到;茄子、拟南芥的相关序列信息分别获取于茄子EGDB(http://eggplant. kazusa. or. jp/)数 据 库[23]和拟南芥TAIR 数据库。采用Clustal Omega 进行多序列比较分析,并使用Jalview 进行作图;分子进化树利用MEGA-X 进行构建,自展值为1000,并使用FigTree 进行作图。
1.3 qRT-PCR
对收取的托鲁巴姆和茄子样品进行总RNA提取,使用Nanodrop 进行RNA 样品纯度检测以及浓度定量,以保证使用的RNA 样品合格和后续实验的进行。使用Roche LightCycler 480 荧光定量PCR 仪进行qRT-PCR。以Actin8基因作为内参,每个基因3次重复,使用2-ΔΔCt方法进行基因表达分析[24],显著性差异t检验P<0.05,引物序列见表2。

表2 引物序列Table 2 Primers of qRT-PCR
1.4 根长测定
使用Epson Perfection V500 Photo 扫描仪对处理4 d 的拟南芥幼苗主根进行扫描,并采用Image-J 对主根生长量进行测量。利用SPSS 软件对数据进行统计分析,显著性差异t 检验P<0.05。
2 结果与分析
2.1 序列同源性比较分析
对茄子SmZIPs 以及托鲁巴姆StZIP5 和StZIP11 的蛋白序列进行了多序列同源性比较分析(图1)。分析表明托鲁巴姆StZIP5 和StZIP11与茄子ZIP 家族成员之间的氨基酸序列具有较高的同源性,预示了托鲁巴姆StZIP5 和StZIP11 与茄子ZIP 家族成员之间具有类似的结构以及发挥相似或相同的功能。进一步分析发现,StZIP5 和SmZIP5 之间的序列一致性为97.97%,StZIP11 和SmZIP11 之间的序列相似度为96.19%(表3)。
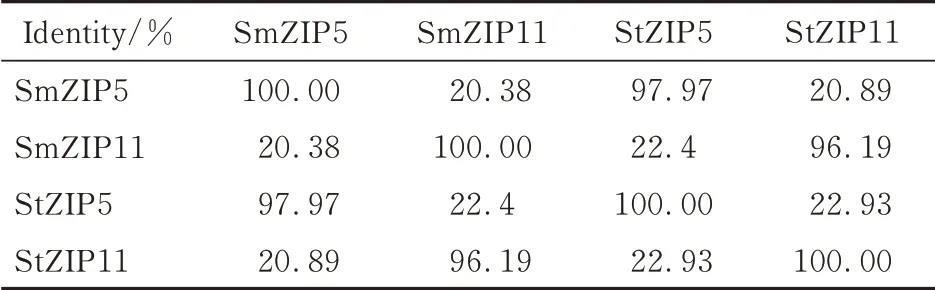
表3 StZIP5/11 和SmZIP5/11 序列的相似性百分比单位矩阵Table 3 Percent identity Matrix of sequences in StZIP5/11 and SmZIP5/11

图1 SmZIPs 的序列同源性分析Fig.1 Sequence identity analysis of SmZIPs
ZIP 蛋白家族之间的进化关系显示,不同物种之间的同名ZIP 成员具有很高的亲缘关系(图2),参与分析的ZIP 成员在进化上被分为了4个进化分支,ZIP2 和ZIP11 形成的进化分支长度最长,ZIP6 形成的进化分支个体成员最少。并且,StZIP5 和SmZIP5 之间的差异度为0.00,StZIP11和SmZIP11 之间的差异度为0.01,表明StZIP5 和SmZIP5 以及StZIP11 和SmZIP11 之间的仅具有极小的序列差异。

图2 拟南芥、茄子的ZIP 家族成员和托鲁巴姆StZIP5 和StZIP11的分子进化树Fig.2 Phylogenetic relationship of S.torvum StZIP5 and StZIP11 with other ZIP family members in A.thaliana and S.melon⁃gena
2.2 锌毒害对托鲁巴姆和茄子ZIP5 和ZIP11 基因表达的影响
对Zn 毒害下托鲁巴姆和茄子幼苗ZIP5和ZIP11基因在根、茎、叶中1、3 d 时表达水平进行了分析(图3)。StZIP5基因的表达水平均显著低于对照组;SmZIP5基因的表达水平在1 d 时的根、茎中同样显著低于对照,但在叶中1 d 时显著高于对照;在3 d 时,根中显著高于对照,茎中与对照组无显著差异,而叶中显著低于对照。StZIP11基因在根中1 d 的表达水平显著高于对照,3 d 时的表达水平较对照组无显著差异,SmZIP11基因在根中1 d的表达水平较对照组无显著差异,3 d 时的表达水平显著高于对照;StZIP11基因在茎中的表达水平较对照组均无显著差异,SmZIP11基因显著高于对照;StZIP11基因在叶中1 d 的表达水平显著高于对照,3 d 时的表达水平较对照组无显著差异,SmZIP11基因在1 d 和3 d 时均显著高于对照。

图3 锌毒害下托鲁巴姆和茄子ZIP5 和ZIP11 的基因表达Fig.3 Gene expression of ZIP5 and ZIP11 in S.melongena and S.torvum under high Zn stress
2.3 镉毒害对托鲁巴姆和茄子ZIP5 和ZIP11 基因表达的影响
对Cd 毒害下托鲁巴姆和茄子ZIP5和ZIP11基因的表达量进行了分析(图4)。托鲁巴姆和茄子幼苗Cd 毒害处理1、3 d 时,StZIP5基因在根中的表达水平较对照组无显著差异,而SmZIP5基因的表达水平在3 d 时显著高于对照;StZIP5基因的表达水平在茎中3 d 时显著低于对照,而SmZIP5基因的表达水平在1、3 d 时较对照组无显著差异;StZIP5和SmZIP5基因在叶中1 d 的表达水平均显著高于对照,但StZIP5基因3 d 时于对照组无显著差异,SmZIP5基因3 d 时显著低于对照。StZIP11和SmZIP11基因在根、茎、叶中1、3 d 时的表达水平均显著高于对照组。
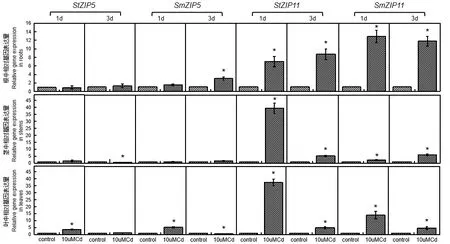
图4 镉毒害下托鲁巴姆和茄子ZIP5 和ZIP11 的基因表达Fig.4 Gene expression of ZIP5 and ZIP11 in S.melongena and S.torvum under high Cd stress
2.4 StZIP5 或StZIP11 在拟南芥野生型Col-0、突变体zip5 或zip11 突变体中的异源表达增加了Zn/Cd 毒害的敏感性
野生型拟南芥Col-0、拟南芥突变体zip5和zip11、转基因拟南芥StZIP5OX、StZIP11OX、StZIP5OX/zip5、StZIP11OX/zip11、StZIP5OX/zip11和StZIP11OX/zip5的主根生长量在45 μmol·L-1Cd 处理下均显著降低(图5)。但zip5和zip11的相对主根长量显著高于Col-0;StZIP5OX、StZIP11OX的相对主根长量显著低于Col-0。StZIP5OX/zip5、StZIP11OX/zip5的相对主根长量显著低于zip5;StZIP5OX/zip11、StZIP11OX/zip11的相对主根长量显著低于zip11。

图5 Cd 毒害下拟南芥的主根生长量和相对主根生长量Fig.5 Principal root growth and relative principal root growth of A.thaliana under high Cd stress
400 μmol·L-1Zn 处理下,拟南芥的主根生长量和相对主根生长量结果如图6所示。Col-0、zip5、zip11、StZIP5OX、StZIP11OX、StZIP5OX/zip5、StZIP11OX/zip11、StZIP5OX/zip11和StZIP11OX/zip5的主根生长量在400 μmol·L-1Zn 处理下同样被显著抑制。并且,zip5、zip11的相对主根长量显著高于Col-0;StZIP5OX、StZIP11OX的相对主根长量显著低于Col-0;StZIP5OX/zip5、StZIP11OX/zip5的相对主根长量显著低于zip5;StZIP5OX/zip11、StZIP11OX/zip11的 相 对 主 根长量显著低于zip11。
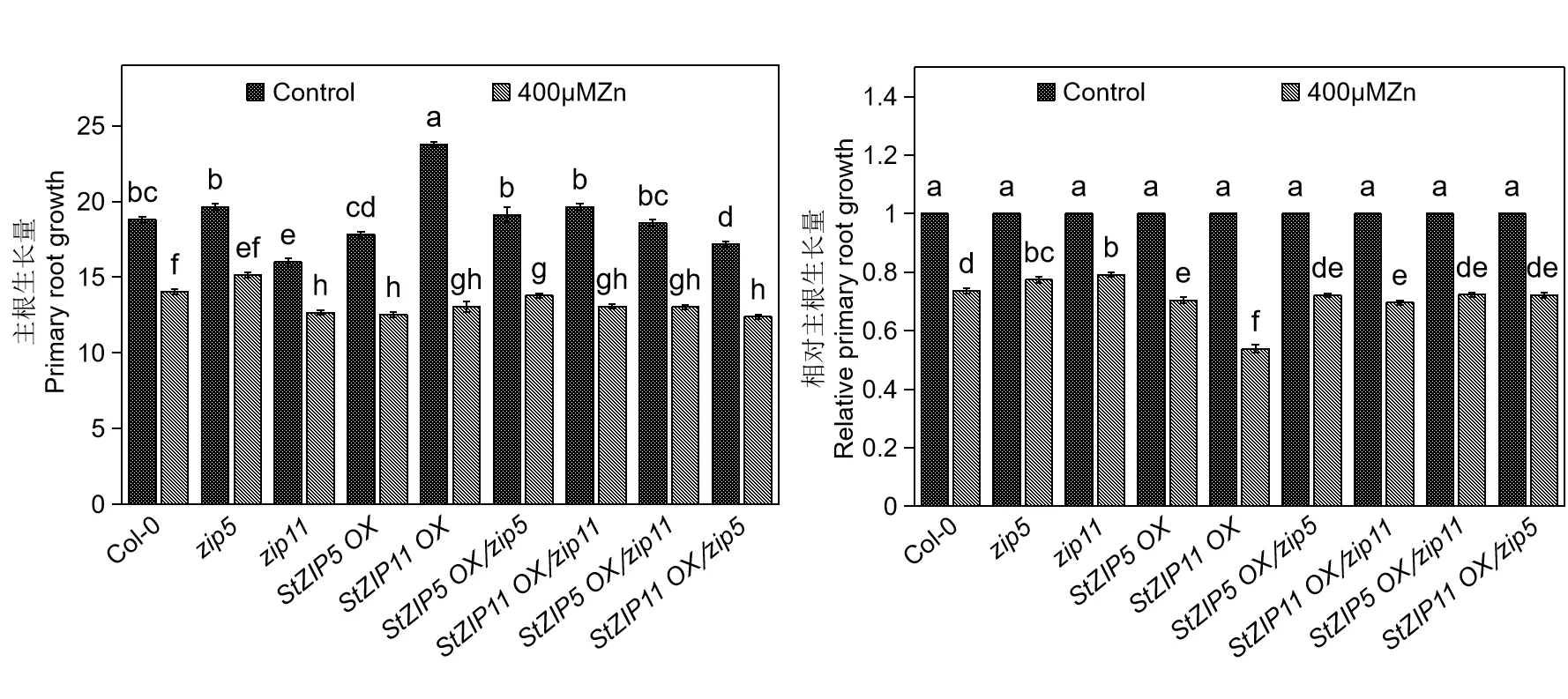
图6 Zn 毒害下拟南芥的主根生长量和相对主根生长量Fig.6 Principal root growth and relative principal root growth of A.thaliana under high Zn stress
3 讨论
本研究中,茄子SmZIPs以及托鲁巴姆StZIP5和StZIP11的蛋白序列同源性比较分析结果表明,茄子ZIP家族蛋白成员之间在功能结构区的氨基酸序列相对保守,与蒲琦[11]、傅明辉等[25]论述的对植物ZIP基因家族结构的观点结果一致。StZIP5和SmZIP5 之间的序列一致性为97.97%,StZIP11和SmZIP11 之间的序列一致性为96.19%。分子进化树显示,托鲁巴姆StZIP5、StZIP11 和茄子SmZIPs 与拟南芥同名ZIP 家族成员具有较高的亲缘关系。并且StZIP5 和SmZIP5 之间的差异度为0.00,StZIP11 和SmZIP11 之间的差异度为0.01,这些结果说明StZIP5 和SmZIP5 以及StZIP11 和SmZIP11 之间的仅具有极小的序列差异。
在200 μmol·L-1Zn 处理下,除StZIP11基因在叶中1 d 时的表达水平显著升高(3 d 时较对照组无显著差异)外,其余时间点StZIP5和StZIP11基因的表达水平较对照组显著降低或无显著差异。推测在Zn 毒害下,托鲁巴姆通过降低StZIP5和StZIP11基因的表达量来减少StZIP5 和StZIP11转运蛋白的含量,从而降低对Zn 的吸收以维持植物 体Zn的平衡。Lee 等[26]在Zn毒害下通过水稻OsZIP5基因表达结果也验证了这一结论。茄子在200 μmol·L-1Zn 处 理3 d 时,SmZIP5和SmZIP11基因在根中显著高表达,暗示茄子在Zn 毒害过程中吸收了较多的Zn。而在叶中,SmZIP5和SmZIP11基因在应对Zn 毒害时表达水平先升高后降低,推测在茄子叶中先通过增加转运分配,后通过降低表达量的方式以减少过量Zn 对叶的毒害作用。与Katarzyna 等[27]认为烟草NtZIP11基因在维持整个烟草锌的供应相符。
Cd 对植物的毒害作用主要是由于Cd 与Fe、Zn之间的竞争吸收,Cd通过Fe或Zn转运蛋白通道进入植物细胞,从而导致植物缺Fe或缺Zn[5]。10 μmol·L-1Cd 处理下,托鲁巴姆StZIP5和StZIP11基因和茄子SmZIP5和SmZIP11基因在根、茎和叶中的qRT-PCR 结果表明,ZIP5和ZIP11基因参与了托鲁巴姆和茄子对Cd 的吸收和转运过程。 托鲁巴姆StZIP11基因和茄子SmZIP11基因在1、3 d 时较对照组均显著高表达,但根中StZIP5基因在1、3 d 时较对照组均无显著,而茄子SmZIP5基因在3 d 时的根中显著高表达。这表明,相对托鲁巴姆,茄子在Cd 毒害过程中吸收了更多的Cd。在茎中StZIP5基因和SmZIP5基因的表达水平均较低,但托鲁巴姆StZIP5在3 d 时较对照组显著低表达。在叶中1 d 时StZIP5基因和SmZIP5基因的表达水平较对照组显著高表达。以上结果表明,茄子可能通过根部吸收大量Cd,并且通过茎向地上部运输,使Cd 在叶中富集。Mori等[3]通过对托鲁巴姆和茄子根部、地上部以及木质部汁液中Cd 的浓度测定结果也验证了这一结论。与茄子不同,托鲁巴姆茎和叶中StZIP5基因表达水平较对照组无显著差异或显著低表达;与Mori等[3]认为茎的截留作用是导致了托鲁巴姆地上部Cd 低累积相符。然而托鲁巴姆茎部对Cd 的具体截留机制还有待进一步揭示。
Cd 和Zn 毒害均能显著抑制植物根系生长[28-29]。在45 μmol·L-1Cd 和400 μmol·L-1Zn 处 理下,野生型拟南芥Col-0、拟南芥突变体zip5和zip11、转托鲁巴姆StZIP5和StZIP11基因拟南芥StZIP5OX、StZIP11OX、StZIP5OX/zip5、StZIP11 OX/zip11、StZIP5OX/zip11和StZIP11OX/zip5的主根生长均受到不同程度的抑制,与Castiglione等[28]得出的Zn 毒害能显著抑制植物根系生长一致。拟南芥AtZIP5或AtZIP11基因的功能丧失突变体(zip5和zip11)主根生长表现出了对Zn 和Cd胁迫显著的耐受性;托鲁巴姆StZIP5或StZIP11基因在拟南芥野生型Col-0 中的过量表达显著加重了Zn 和Cd 毒害下对拟南芥主根生长的抑制;托鲁巴姆StZIP5或StZIP11基因在拟南芥突变体zip5或zip11中的过量表达也显著加重Zn 和Cd 毒害对拟南芥主根生长的抑制。这些结果表明,托鲁巴姆StZIP5或StZIP11基因在拟南芥中的过量表达增强了Zn 和Cd 毒害对拟南芥主根生长的抑制作用。
4 结论
托鲁巴姆作为一种优良的茄子砧木和重要的Cd 低累积作物之一,其Cd 低累积机理的研究对茄子、番茄等低Cd 蔬菜品种的改良具有极其重要的意义。本研究结果表明StZIP5或StZIP11参与了托鲁巴姆低Cd 累积过程,有助于未来进一步的研究探明StZIP5或StZIP11在托鲁巴姆中调节Cd 累积与耐受的分子机理。
猜你喜欢
科学导报·学术(2020年19期)2020-07-09
阅读与作文(小学高年级版)(2019年9期)2019-11-06
农民致富之友(2019年14期)2019-05-24
小天使·一年级语数英综合(2017年4期)2017-04-18
江苏农业科学(2016年11期)2017-03-21
小天使·一年级语数英综合(2016年9期)2016-05-14
汉语世界(The World of Chinese)(2016年3期)2016-01-10
飞碟探索(2015年9期)2015-11-05
红领巾·探索(2015年9期)2015-09-10
恋爱婚姻家庭·养生版(2011年8期)2011-05-14
