
蝙蝠,听觉和回声定位研究的模型动物*
2022-05-23 07:43付子英陈其才
生物化学与生物物理进展 2022年5期
付子英 唐 佳 陈其才
(华中师范大学生命科学学院,武汉 430079)

普氏蹄蝠(Hipposideros pratti),用于听觉和回声定位研究的蝙蝠之一。
孔子曰:“工欲善其事,必先利其器”。阿基米德(Archimedes)也曾说:“只要给我一个支点,我就可以撬起一个地球”。纵观人类社会,几乎每前进一步都与利用工具息息相关。在生命科学研究中,人们通常将那些适用于研究的生命体即实验对象,称为模式(型)生物(model organism),包括模式植物(model plant)、模式动物(model animal) 和模式微生物(model microorganism),从而成为研究中最重要的“工具”之一。在生物演化中,许多生命活动的基本方式和原理,如遗传原理、生理过程及机制等,在众多生物物种中是保守和共享的。因此,在模式生物上获得的研究结果,可用于揭示在众多生物物种中具有普遍规律的生命现象,这正是模式生物研究策略能够成功的基础。人们对生活于黑暗环境中的蝙蝠如何感知外界环境有着浓厚的兴趣。从1793 年意大利生理学家斯帕兰札尼(Spallanzani)发现盲眼蝙蝠仍能自由飞行和生存,到1938 年皮尔斯(Pierce) 和格里芬(Griffin)成功地记录到蝙蝠发出的超声,100多年的研究最终揭开了这一谜团——蝙蝠是通过主动发出声音信号,并听其回声来感知周围环境。Griffin[1]于1944 年提出将自然界中任何利用类似声纳原理探测周围环境的过程称之为回声定位。Grinnell[2]记录到蝙蝠发出的超声也被认为是蝙蝠回声定位研究中一项具有里程碑意义的发现。自此以来,科学家以蝙蝠为模式动物,从听觉[3-8]、回声定位[9-11]和生态适应与演化[8,12-17]等方面开展了研究,取得了令人瞩目的成果。本文主要从神经生物学的角度,对蝙蝠听觉和回声定位方面的研究及认识做简要介绍。
1 蝙蝠的生物学性状与分类
蝙蝠是神奇而又特别的动物。经过长期的演化,其中回声定位蝙蝠(以下所述及的蝙蝠皆与此意相同)听觉系统的结构和功能均达到了一种完美的高度[12-13],成为了黑暗环境中的“精灵”和捕食高手。
蝙蝠在生物学上属于哺乳纲(Mammalia),翼手目(Chiroptera),是哺乳动物中仅次于啮齿类动物的第二大类群,也是唯一能真正飞行的哺乳动物。现发现有大约1400 种[18],约占哺乳动物的25%[10],几乎分布于除南、北极以外的所有地区。中国是世界上蝙蝠物种多样性最丰富的国家之一,已知的种类超过了140 种[19]。根据蝙蝠的形态等特征,进一步将其分为小(或阳) 蝙蝠亚目(Yangochiroptera) 和 大(或 阴) 蝙 蝠 亚 目(Yinpterochiroptera),阴蝙蝠亚目蝙蝠主要通过视觉导航,而阳蝙蝠亚目蝙蝠视觉严重退化,听觉系统特化以适应通过回声定位获取周围环境信息的需要,因此被称为回声定位蝙蝠[3,9,11,20]。就体型大小 而 言 , 大 黄 蜂 蝙 蝠 (Crasceonycteris thonglongyai) 最小(约1.5 g),狐蝠(Pteropus spp.)最大(约1000 g,翼宽约1.5 m),大多数回声定位蝙蝠体重基本在100 g以内。回声定位蝙蝠种类繁多,回声定位叫声也多种多样,现已开始在生物多样性国家(megadiverse country)建立蝙蝠回声定位叫声公共数据库(echolocation call library for bats)[21-22]。为简便,根据它们发出的回声定位声信号的频率构成模式和特征,大体上归为3种类型[3,12-13,23-24](图1): 恒 频- 调 频(constant frequency-frequency modulation,CFFM) 蝙蝠,如胡须蝠(Pteronotus parnellii rubiginosus) 等,其发声信号开始为一段时程极短的FM成分(其作用鲜见有报道),继之为一段较长时程的CF成分再接一段短时程的下扫FM 成分(图1 左);调频(frequency modulated,FM) 蝙 蝠,如 大 棕 蝠(Eptesicus fuscus)等,其发声信号为下扫FM 声(图1 中);咔嚓声(Click)或咔嚓样声(Clicklike)蝙蝠,也称为捡食蝙蝠(gleaning bat),如大蝙蝠亚目的果蝠(Rousettus),通常发出时程极短(40~50 μs)的宽频带(可达80 kHz)声音信号,它们也具有回声定位能力[25],但所受到的关注远少于CF-FM 和FM 蝙蝠(图1 右)。通过从遗传学角度比较3 类蝙蝠耳蜗转录组(cochlear transcriptomics),研究发现在基因表达方面,CFFM 蝙蝠与Click 蝙蝠间的差异要大于FM 蝙蝠与Click 蝙蝠间的差异,FM 蝙蝠与Click 蝙蝠之间差异较小,这些结果为3类回声定位蝙蝠间的差异提供了遗传学方面的证据[26]。很多回声定位蝙蝠的发声信号具有性别差异,如雄性普氏蹄蝠(Hipposideros pratti)(一种CF-FM 蝙蝠)回声定位信号主频(第二谐波的CF)高于雌性[27]。
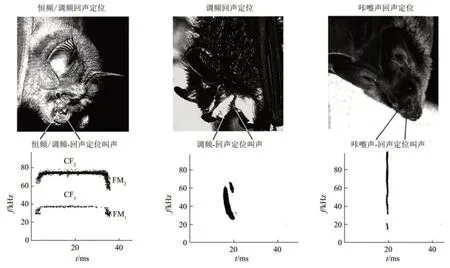
Fig.1 Echolocation systems in bats.CF/FM:horseshoe bats,hipposiderids,and the neotropic moustached bat emit multiharmonic echolocation signals consisting of a long pure tone terminated by a brief FM component.CF/FM bats often forage within or close to dense vegetation FM.Most insectivorous bats foraging on the wing emit brief,downward-FM echolocation signals when approaching and catching a prey(FM).They often emit longer and only shallowly modulated signal when they search for prey(not shown).Gleaning bats and some flower visiting bats emit very brief signals over a broad frequency band(Click-like).These echolocation sounds are far less intense than those of FM and CF/FM bats[12]图1 根据回声定位蝙蝠所发出的声音信号模式将其分为3大类型:恒频-调频(CF-FM)蝙蝠,通常发出由多个谐波构成的回声定位信号,在长的纯音尾端接一段短的下扫FM成分;FM蝙蝠,通常发出下扫较浅调制的FM信号;咔嚓样声(Click-like)蝙蝠又称为捡食蝙蝠(gleaning bat),通常发出时间非常短的宽频带声音信号[12]
2 蝙蝠的听觉和回声定位研究
回声定位蝙蝠的体型大小及其听觉功能的高度特化,使得它们具备了适合用于听觉和回声定位研究的诸多优势[5,9,28],被相关领域的研究者认为是良好的听觉和回声定位研究的模式动物。
2.1 回声定位蝙蝠听觉系统的特化
蝙蝠听觉系统的基本结构、传导通路及功能与一般哺乳动物类似。但由于该系统是蝙蝠回声定位系统的组成部分之一,为适应回声定位的需要,听觉系统结构和功能与非回声定位动物相比,产生了明显的特化[5],使该系统被“放大”,从方法学上为实验研究提供了便利。
2.1.1 外周听觉系统的特化
蝙蝠的外周听觉系统包括外耳、中耳和内耳。外耳形状多样,显现出对接收回声定位信号的高度适应。为了避免自身发出的高强度声信号对耳蜗造成伤害,附着于镫骨上的镫骨肌特化成为特殊的声幅度衰减结构,在蝙蝠发出高强度回声定位信号的一瞬间,可预收缩并向外拉动镫骨以降低对卵圆窗的撞击,进而削弱强声信号对耳蜗的冲击,一旦发声完毕,镫骨肌松弛恢复正常[29]。内耳耳蜗不像一般哺乳动物那样牢固地嵌入颞骨内或与之融合固化,而是悬挂于内耳腔中,周围有脂肪组织缓冲并衰减强声引起的震动。最新研究揭示,相较于实验室小鼠和非回声定位蝙蝠,回声定位蝙蝠耳蜗毛细胞高表达ISL1基因,这可能是蝙蝠能够耐受高强度声暴露的原因[30]。蝙蝠在捕食终末相发出的声脉冲重复率高达200 Hz,镫骨肌能以相同频率产生收缩运动[5]。内耳所占体重比非常大,内耳耳蜗沿蜗轴旋绕2.5~3.5 圈,且底圈增大以适应感受高频率的回声定位信号[5],而灵长类动物耳蜗仅旋绕约1.75 圈[23],采用脑核磁共振成像和组织学比较研究也显示,使用CF-FM和FM声脉冲进行回声定位的蝙蝠要比啮齿类大鼠和小鼠有更大的耳蜗和更多的转数(turns),这些差别可能与它们的回声定位行为相关[31]。内耳的结构也极其精细,在CF-FM 蝙蝠中,对回声定位信号第二谐波的CF(CF2)又称为主频(dominant frequency,DF)及其临近频率产生过度表征,并形成了一个被称为“听觉凹(auditory or acoustic fovea) ”的特化区[3,12,32]。该区域的神经元具有极强的声音强度耐受性和锐化的频率调谐的能力,能精确感受和调谐多普勒频率漂移,并锁定昆虫翅振产生的周期性频率变化[33]。相比起来,FM蝙蝠这方面的特化并不明显。近期研究还提示,蝙蝠耳蜗听神经螺旋神经节的解剖学差异可能与蝙蝠类型的演化有关[20]。
2.1.2 中枢听觉系统的特化
蝙蝠从延脑(medulla oblongata) 到听皮质(auditory cortex,AC),由多个(级)听觉核团构成上行的中枢听觉通路。
a. 耳蜗核(cochlear nucleus,CN):在一些CF-FM蝙蝠,表现出CN核团增大和细胞分化,尤其是神经元分化出对来自“听觉凹”频率范围的信号有精确的分析能力[4,34-35]。
b.上橄榄复合体(核)(superior olive complex or nucleus, SOC) 和 斜 方 体 核(nucleus of trapezoid body,NTB):在结构与功能方面与一般哺乳动物相比,SOC 要比NTB 分化明显,可分为不同亚核,其中内侧上橄榄核(medial SO)被认为主要用于分析回声定位信号到达双耳的时间差[36]。
c. 外侧丘系核(nucleus of lateral lemniscus,NLL):蝙蝠的NLL也不乏出现某些特化[37]。有些亚核出现体积增大,甚至出现不同的功能区[38],一些神经元也表现出对回声定位信号谐波的联合[39],以及对FM扫频方向[40]敏感等。
d.下丘(inferior colliculus,IC):蝙蝠中脑的IC 是听觉中枢上、下行投射的重要中继站和整合中枢[6,41]。与其他哺乳动物相比,蝙蝠IC 更为发达,在大脑中占据了很大比例,且外凸扩展覆盖了部分上丘(superior colliculus,SC)。IC 的频率表征排布成等(同)频层状,频率变化沿背腹轴由低到高[5]。频率层的厚度与蝙蝠回声定位信号频率相关,如FM蝙蝠大棕蝠常用回声定位信号频率为20~30 kHz,在IC 就 占 据 了 较 大 比 例[42];而CF-FM蝙蝠胡须蝠“听觉凹”60~63 kHz的窄段频率等频层高度扩展[12,43]。IC 神经元在回声定位方面也具有了高度的适应性和选择性。相关研究发现,在蝙蝠中脑内存在延迟调谐[44]、多普勒频率漂移补偿[45],以及对自然声信号具有选择性[46]等特性的神经元,显示出其特化与回声定位行为上的匹配,这种特化或许由适应其生境需要所驱动的演化所致。
e. 内 侧 膝 状 体(medial geniculate body,MGB):蝙蝠丘脑MGB 可分为不同的亚核,主要接受同侧IC的上行投射和同侧听皮质(AC)的下行投射。为适应回声定位,MGB 与听觉相关的神经元产生了某些方面的特化,其中70%神经元对CF1与CF2联合敏感,30%神经元对CF1与CF3联合敏感,而对FM1与FM2、FM1与FM3联合敏感神经元的比例基本相当,另外,还存在对声时程敏感[47-49]以及对延迟调谐联合敏感性(delay-tuned combination sensitivity)[50]的神经元。
f. 听皮质(AC): 通常指初级听皮质(primary AC,AI),是听觉信息加工的高级中枢,在整个蝙蝠脑皮质中占据了相当大的比例,但对其所做的研究相对少于皮质下的一些听觉结构。在FM 蝙蝠大棕蝠的AC 研究发现,频率表征沿前后轴依次按前高后低的顺序排布,相同频率的神经元由内到外排布成同频带,而每个同频带内的神经元又形成若干贯穿于整个皮质厚度的柱状组构,同一柱内的神经元具有相似的声反应特性。为适应回声定位,在CF-FM 蝙蝠-胡须蝠的AC 形成了分区的功能模块[32,51-52],如多普勒恒频漂移区(Dopplershift CF area,DSCF)、CF/CF 多普勒漂移幅度区(Doppler magnitude),以及FM/FM 回声延迟或靶物距离区(echo-delay or target range)等,各模块内的神经元有各自的反应特性[5,11]。
2.2 蝙蝠的听觉与回声定位
由于回声定位蝙蝠的听觉系统与回声定位系统密不可分。在过去80 多年的时间里,以回声定位蝙蝠为模型,在神经生物学层面,对其听觉及回声定位做了大量的研究,对其所取得的成果和认识,在先前的一些报道和专著中均做过相应的评述[5,11,53-54],主要聚焦在以下几方面。
2.2.1 动态的反应潜伏期
神经元反应的潜伏期随刺激条件尤其是刺激强度的改变而呈现动态改变的特性,也是所有可兴奋细胞对刺激反应共有的基本特性之一。然而,对于蝙蝠不仅仅是基本特性,也是回声定位行为调控的一部分[11]。不同种类动物的AC神经元反应特性不同,猴、猫的AC神经元通常对声刺激呈现出持续型或紧张型反应(sustained or tonic response)[55];而蝙蝠的AC神经元反应往往呈现出非常短暂的相位型反应,这种反应模式的神经元也大量存在于蝙蝠AC下的听觉核团内[56]。那些短暂反应的潜伏期还显示出一种重要的功能特性,即有较长潜伏期的神经元有较尖锐的回声-延迟调谐,而有较短潜伏期的神经元则有较宽的回声-延迟调谐[57]。因此,在每次回声定位叫声后,蝙蝠AC神经元群便启动表达关于靶物粗略的位置信息,然后随时间推移,超速地锐化听像,与采用多重分辨成像加工算法锐化视觉图像[57]。类似这种短潜伏期神经元还能跟随蝙蝠捕食终末相的高声脉冲重复率,成为听觉系统快速采集靶物信息的一种重要手段,与其捕食行为相匹配[58]。Sallesa等[59]研究显示,蝙蝠发出的高频率回声定位声脉冲,能以类似于“高速摄影”的方式进行“声学快照(acoustic snapshot)”,以采集并累积信息来判断听觉目标的移动。
2.2.2 频率调谐和“听觉凹”
频率调谐是人和动物听觉系统的一项基本的听觉功能,但回声定位蝙蝠,尤其是CF-FM 蝙蝠,对频率调谐的能力和精准程度远胜于人和其他动物。如CF-FM 蝙蝠胡须蝠和马蹄蝠,显示出异常锐化的频率调谐,从耳蜗到听皮质的整个上行听觉通路,回声定位信号主频CF 成分被过度表征,形成了一个类似于视网膜“中央凹”的结构,称其为“听觉凹”[11-12,32]。该区域的神经元具有尖锐的频率调谐曲线,可满足蝙蝠探测靶物回声多普勒频率漂移(Doppler shifts)的需要,听觉系统或可从中计算出昆虫移动的速度,以及探测出由昆虫翅振引起的快速多普勒调制[5]。近期的研究还显示,普氏大蹄蝠(Hipposideros pratti)IC神经元对回声定位行为相关的第二谐波CF(即主频)成分具有尖锐的频率调谐和很高的选择性[60],而且尖锐的频率调谐可能参与蝙蝠在噪声条件下的回声定位信号加工[61]。
2.2.3 声时程调谐
声时程是回声定位蝙蝠发声信号主动改变的参数之一[11]。声时程调谐或选择性神经元(durationtuned or selective neurons)最先在蛙的听觉中脑内被发现[62-63]。后来在蝙蝠、小鼠以及大鼠等动物的中脑和听皮质中均发现存在有时程调谐神经元[63-65],可见时程调谐是一种跨脊椎动物物种的神经生物学特性[66]。这种功能特性对于在听觉加工过程中从感觉输入中提取时相信息至关重要[64,67-68]。蝙蝠听中枢的时程调谐神经元呈现出低通、高通和带通等不同的类型,可能是为了匹配对回声定位信号时相信息加工的需要,从而在对叫声时相快速动态变化中的时相特征加工方面发挥重要作用[11,69]。这些时程调谐神经元不仅提供了一种编码声音时相特征的重要神经机制,而且也与频率调谐和神经组构有关。通过记录清醒大棕蝠IC单个神经元的反应,测定听时程调谐神经元的波谱-时相调谐特性及沿IC背-腹的分布位置,结果显示波谱时相敏感性和空间组构在IC 有两种模式:具有锐化的频率调谐和宽的时程调谐特性的神经元位于背侧IC,而那些具有宽的波谱调谐和窄的时相调谐的神经元则位于腹侧IC[70]。另有研究还发现,在清醒大棕蝠的听觉中脑不仅存在时程调谐神经元,而且还存在能解码刺激时程的神经元[71]。
2.2.4 回声-延迟(靶物距离)调谐神经元
研究发现,在好几种回声定位蝙蝠的听觉中枢均含有回声-延迟调谐神经元,这些神经元被认为参与蝙蝠对靶物距离的判断。例如,当用特定时间间隔的声信号来模拟“脉冲-回声对”刺激时,这些神经元显示出易化反应(facilitated responses),在最佳时间间隔即最佳延迟时反应达到最大[5,11-12,57,72]。研 究 发 现,大 棕 蝠 中 脑 上 丘(midbrain superior colliculus)[73]、丘脑和听皮质[50]均发现有回声-延迟调谐神经元,这些神经元不仅能对回声-延迟进行调谐,而且也能对回声的方位角(azimuth)和俯仰(elevation)方位进行调谐,从而在靶物位置的三维(3D)表征方面发挥重要作用[11]。一直以来认为蝙蝠需要将回声延迟时间转换成为距离以评估猎物的距离,它们是通过学习来掌握这种关系,还是生来就已经编码好存于脑内,且在成年后还能调整?针对该问题,近期的研究发现,新生和成年蝙蝠均不能对其进行调整,提示声速在蝙蝠脑内被先天编码,而且,还提示蝙蝠按照时间来编码外部世界,并不将时间转换成为距离,这些结果解释了先天的演化和灵活的感官知觉[74]。
2.2.5 神经元对回声定位行为相关声信号的反应和加工模式
蝙蝠的听觉系统能够高效地识别和处理行为相关声信号[75]。行为学研究显示,在回声定位过程中,CF-FM蝙蝠回声定位信号CF成分主要用来分析速度相关信息,而FM成分主要用来分析距离以及靶物细节信息[32,76]。近年来,采用与大蹄蝠和普氏蹄蝠回声定位行为相关的CF-FM 声刺激,记录下丘单个神经元的声反应特性,发现下丘神经元对这种行为相关的CF-FM 声信号表现出single-on(SO)和double-on(DO)两种反应模式,SO神经元对仅在CF-FM声刺激CF成分的开始产生1次on反应,而DO神经元在CF-FM声刺激CF成分和FM成分的开始分别产生1 次on 反应[77-78]。进一步研究还发现,与SO 神经元相比,DO 神经元具有较短的反应潜伏期、恢复周期以及频率选择性,提示两种类型的神经元可能在蝙蝠回声定位的不同时相发挥作用[60,79-80],可见SO 和DO 反应为回声定位行为相关的声反应。采用在体细胞内记录的方法研究行为相关声反应的形成机制,发现SO神经元的锋电位后超极化(post-spike hyperpolarization,PSH)的持续时间显著高于DO 神经元[78],且SO神经元的PSH 不受声刺激时程和强度的影响,而DO 神经元的PSH 随着CF 时程和强度的增加而缩短[81]。这种较长时间的PSH 可能抑制了FM 成分诱发的反应,使神经元表现为SO反应。为比较行为相关声反应的物种差异,采用CF、FM 和CF-FM 声刺激,记录了CF-FM (Hipposideros pratti)和FM(Pipistrellus abramus)蝙蝠的下丘诱发电位反应。所获得结果显示,CF 声诱发CF-FM 蝙蝠产生on-off 反应,而FM 蝙蝠对CF 和FM声刺激均只能产生on反应,提示2种类型的蝙蝠可能存在种属特异性的神经回路。然而,当用CF-FM声刺激时,2种蝙蝠下丘均能对CF和FM成分产生诱发电位反应,推测回声定位蝙蝠可能存在共性的神经回路[82]。
行为相关声信号的加工具有噪声耐受性[83]。人或动物可通过声信息交流获取目标信息,以适应生活、生存以及繁殖的需要。然而,自然环境中不可避免地存在一定程度的噪声,包括生物噪声及非生物噪声,可能会对声信息的识别和加工存在影响。研究显示,斑胸草雀、雪貂等动物听觉高位核团(如听皮层)神经元对行为相关声信号的反应不受背景噪声的影响[84-86],这可能是行为相关声信号的加工具有噪声耐受性的机制。在普氏蹄蝠上的研究显示,在背景噪声条件下,普氏蹄蝠会提高回声定位信号的强度,但维持回声定位信号的主频不变。在体电生理记录显示,调谐在回声定位信号第二谐波的下丘神经元(H2 神经元)在相同的背景噪声下听觉阈值增加,且增加量与发声强度增加量匹配,而下丘神经元的最佳频率以及频率选择性不受背景噪声的影响,结果提示普氏蹄蝠下丘神经元在背景噪声条件下的声反应特性与其回声定位行为相适应[61,87]。
2.2.6 皮质功能组构模块化
对胡须蝠的听皮质研究显示,根据神经元的反应特性可划分为不同的功能模块,分别处理不同的回声信息[11](图2)。在DSCF,含有特化的对昆虫翅振(wing flutter or beat)引起的多普勒频率漂移敏感的快速多普勒调制神经元;在CF/CF区则含有选择性地对多普勒漂移幅度即靶物速度敏感的神经元;而FM/FM 区含有选择性地对回声延迟或靶物距离反应的神经元[5,51-52,88]。在蝙蝠AC 上研究所发现的功能特化区或模块化区,实质上也是对感觉皮质局部区域特化观念提供了一个重要佐证[11]。有趣的是,研究还发现这些皮质区除了对蝙蝠的声纳叫声产生反应外,也对它们的通讯叫声起反应[89],提示对这些脑区基本作用的认识不能完全局限于传统。然而,这种令人称奇的功能组构并未在FM蝙蝠的听皮质上发现[5],为何种类间的差异如此之大?这个问题或许要借助一种新的能将一个给定皮质区所有神经元的反应有效成像的光学记录技术,以及学科间的联合来解决[90]。尽管如此,但对苍白蝠(Antrozous pallidus)(一种FM 蝙蝠)听皮质的研究,仍不乏有新的发现,观察到在反应上具有选择性的两个分离区,功能上也明显不同,分别负责回声定位和对猎物产生的噪声进行定位,通过比较噪声选择区和回声定位区即FM扫频选择区神经元之间的空间感受野,结果提示这两种不同的声定位行为的机制可能不同[91]。在另一种FM蝙蝠——游离尾蝙蝠(Tadarida brasiliensis)听皮质上研究也发现,它们对所发出FM回声定位信号的扫频方向选择性呈现出一种层状组构[92]。这些研究提示,FM蝙蝠听中枢某些功能相关的组构模式不是完全没有踪迹可寻,相信随着研究的不断深入,那些潜在的或隐匿的奥秘最终会见端倪。
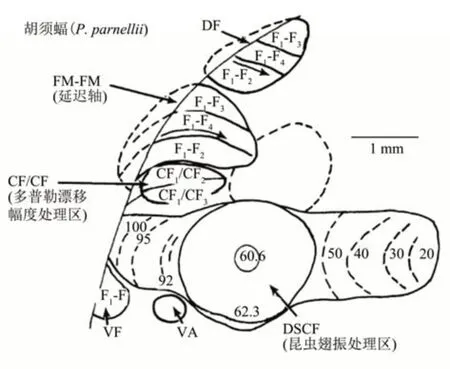
Fig.2 Electrically functional organisation of the auditory cortex of P.Parnellii with an auditory fovea图2 CF-FM蝙蝠胡须蝠(P.parnellii)听皮质的电生理上不同的功能分区或模块化
2.2.7 中枢对声信号加工的神经调控
以回声定位蝙蝠为模式动物,在中枢声信号加工的调控及其对可塑性的影响方面做了诸多研究,包括高位中枢对低位中枢的离皮质调控[93-94]、不同中枢间的相互作用与调控[95-97]、双侧同中枢间的相互作用[98],以及同一中枢内细胞间的相互作用[99-100]。20 世纪末,在大棕蝠[93]和胡须蝠[94]上发起了一系列有关离皮质调控的研究,证实了蝙蝠脑内存在对听觉输入精细的离皮质调控机制。研究发现,这种调控作用是多方面的,如离皮质调控系统能影响蝙蝠下丘听敏感性[93,101],改变下丘神经元的各类强度-发放率函数[102]等;而另一些研究发现,电刺激大棕蝠听皮质可影响同侧IC 神经元对声信号强度和频率的敏感性[103-104],并调制神经元的声反应和频率调谐[105],以及调制听觉系统对声频率信息处理和生物声纳时间域信息的处理[94,106],且其调制作用不仅可以影响即时的声信号加工,还可引起听觉系统出现可塑性的改变[107-108]。离皮质调控还能影响听觉外周-耳蜗内毛细胞对声刺激的敏感性[109-110]。用电生理方法从功能上对离皮质调控通路研究,结果显示有不同的路径。如电刺激内侧膝状体,观察到皮质对下丘的反馈调控可通过下丘-内侧膝状体-皮质-下丘环路来完成[111];当用电刺激听皮质,同时在下丘中央核和外侧核记录其反应,并采用可逆性阻断的方法,发现亦存在一条皮质-下丘外侧核(ICx)-下丘中央核(ICc)的下行反馈通路[103]。它们对皮质下声信号加工,以及在成年后为适应环境而产生可塑性改变等,均提供了一种结构和功能的保障。
2.2.8 目标选择性神经元与尺度不变性
蝙蝠在巡航、捕食和归巢过程中,需要从回声中选择出那些有意义的目标或物体,包括捕食对象和巡航与归巢的参照物等。研究发现,蝙蝠在追踪某个空间目标时,它们能通过调节其口形聚焦声纳场和调控发声信号的声纳束以锁定目标[112-115]。提示蝙蝠可以成为探讨听觉目标神经表征的一个极好的模型[11]。由此可见,在人类知道用雷达聚焦和锁定目标之前,蝙蝠使用这种技能已经先于人类千万年。有研究证实,在蝙蝠的听皮质内存在目标或物体选择性神经元,这些神经元对听觉目标以一种尺度不变性或大小恒常性方式产生反应[11,116-118]。根据回声对植物进行分类的研究主要集中在FM蝙蝠[119-120],因为FM信号比CF信号能提供更精确的时间信息和更丰富的频率信息,有利于在捕食过程中对植物进行区分和质地探测[23,121]。由于蝙蝠的飞行、植物生长造成的大小改变等,均可使发出的声信号至靶物的入射角度出现差异,但它们却不会造成蝙蝠对靶物的属性和类型判断的失真,这是由于蝙蝠从不同入射角度所反射的一系列回波中提取一些共同的特征,这些特征中含有靶物属性和类别信息[119]。Müller 和Kuc[122]在实验中研究了不同角度的发声在不同类型的植物中产生的回声效果差异,结果显示声波入射角度对回声的影响因植物类型的不同而不同。比如,无花果的叶子在自然状态下表现出一定的方向性特征,而紫杉叶片却没有方向性特征,这些差异中可能就含有属性和类型信息。再例如,某种植物在某个入射角度的某个参数值比另一种植物大,那么在其他入射角度的该参数值也仍会比较大,但程度可能会有所不同[122]。这个结果或许可以解释回声定位蝙蝠是如何保持在不同发声角度下对靶物属性和类型做出判断的[119]。回声中含有的靶物属性和类型信息不随靶物体积大小而改变的特性称为尺度或大小不变性。这与视觉视物类似,对同一类型的物体,不会因其大小不同而改变对物体属性和类型的正确判断。von Helversen[123]的实验证明,在完全黑暗条件下,长舌叶鼻蝠(Glossophaga soricina)能够学会区分相同半径和深度的空心半球与抛物面,并能扩展在训练阶段所提取的回声声学特征,且这些特征与靶物大小无关。他认为,尺度不变性分类的线索,可能与这两种靶物所反射的回声对发声信号入射角度改变的依赖性不同有关,如空心半球可保持恒定,而抛物面可随之改变。并/或与回声的“音质”和频谱模式有关,而与绝对音高无关,犹如歌唱家所演唱的歌曲是否美妙动听和打动人,与其绝对声高无关,而与音域的宽窄、音质的好坏和理解歌曲所赋予的情感有关[123]。有人将呈现给苍白矛吻蝠(Phyllostomus discolor)回声的频谱时相特性转换成人工元音的形式后呈现给人听,实验结果表明蝙蝠的尺度不变性或大小恒常性分类能力与人类对人工元音的分类能力非常相似[117,124],该结果为哺乳动物在听觉的尺度不变性方面遵循共同机制的假说提供了实验证据。人们利用上述研究和发现来为人类服务,在一项人类借助回声定位来探测封闭空间的研究中,所取得成果令人鼓舞[125];时变回声信息不仅让蝙蝠对靶物几何形状识别的效果显著,而且对人类利用回声定位亦具有重要的启示作用[126]。
2.2.9 3D空间定位和导航
无论回声定位蝙蝠导航是瞬间的,还是借助短期记忆或长期记忆,均离不开使用回声定位来“查看”周围环境和标志物,产生的感觉输入激活不同类型的导航细胞实现成功导航。近些年来,Ulanovsky及其团队以蝙蝠为模型,开展了3D定位和导航研究,取得了一系列令人瞩目的成果[127]。他们在蝙蝠上不仅发现了与大鼠[128]等动物相似的位置细胞(place cell)和网格细胞(grid cell),也发现了某些新类型的导航细胞,以及那些与大鼠相同类型的导航细胞在3D 空间下的反应和细胞域差异[129]。蝙蝠大脑不仅能对头部方位进行3D 编码[11,130]和表征[131],而且脑内头方位系统中的混合维度神经元还能进行最佳的动态编码[132],表明蝙蝠完全能根据它们声纳发声回声携带的信息计算出外界物体在3D 空间的位置。 Yartsev 和Ulanovsky[133]训练埃及果蝠在房间中自由飞行,用无线神经-遥测技术记录埃及果蝠海马CA1 区的单细胞放电,同时用高速摄像机记录蝙蝠的位置信息,结果显示,海马CA1 区位置细胞的位置域是各向同性的,且在较大房间时位置域也较大,并能涵盖这两种大小的空间。采用实时监测自由活动动物头部朝向的3个欧拉角:水平方位角(azimuth)、俯仰角(pitch)以及翻滚角(roll),同时用无线神经-遥测技术记录埃及果蝠背侧前海马回(dorsal presubiculum)单个细胞的放电,结果显示,背侧前海马回区存在azimuth、pitch 和roll 调谐神经元(分别称之为azimuth细胞、pitch细胞和roll细胞),且azimuth 调谐细胞的特性与大鼠头部朝向细胞类似。同时,亦存在双联合调谐细胞和三联合调谐细胞,分别对3个或其中2个方位角进行调谐。蝙蝠属于社会性和群居动物,有研究发现,在蝙蝠海马CA1区有部分细胞可以表征其他蝙蝠的位置信息,还有部分可以表征移动的非生命物体的位置信息,这些细胞可能通过社交传感,专门负责获取环境中其他蝙蝠踪迹及其位置的信息,因此将它们称为社交位置细胞[134]。蝙蝠飞行期间对3D 空间的动态表征不仅存在于大脑皮质,也存在于皮质下的听觉中脑[131]。不同部位及不同类型的导航细胞如何整合并指导行为,细节仍不清楚,但目前认为,蝙蝠在3D 空间和飞行状态下,脑内几种主要类型的导航细胞能在航程中各自行使相应的功能,引导蝙蝠直达目的地[129](图3)。Ulanovsky 等对蝙蝠大脑3D 空间定位和导航研究的成果,受到广泛关注,被Abbott 在Nature 上撰文做了专题报道[129],并被列为2019 年《自然》(Nature)十大科学专题报道之一[127]。

Fig.3 Schematic graph shows three-dimension navigation and navigation cells’functions of bats图3 蝙蝠3D导航及不同导航细胞的作用模式图
3 展望
该领域的研究者将继续以蝙蝠为模型,从多学科、多层面来开展蝙蝠回声定位研究,以下几方面的问题有可能会受到更多关注:a.生物声纳系统的各基本组成部分在脑内彼此连通的界面部位在何处?是1个还是多个?彼此间的关系等目前仍不十分清楚。b.快速将感觉信息转换成为运动动作以回避障碍物和捕获猎物,转换的中枢部位如皮质和皮质下如何分工?c.通过回声定位,快速对复杂的听境分析选择出有用的目标或靶物,该过程和能力有多少是属于先天遗传的,又有多少是属于后天习得的?d.目前关于尺度不变性的研究虽然已受到不少关注,但由于其机制十分复杂,致使研究依然局限于对相对简单靶物的分析,对于蝙蝠如何对环境中诸多复杂靶物保持尺度不变性,仍有待进一步研究。e.在蝙蝠的3D 导航研究方面,蝙蝠各类导航细胞获取的信息如何进行整合?不同的功能神经元群和脑区之间怎样协作?空间导航认知与地磁导航之间是何种关系,怎样进行协调和整合?总之,人们通过数十年来对蝙蝠听觉系统和回声定位的研究,大量研究设计的原理得到证实。所取得的成果不仅具有基础理论认识方面的价值,同样也对多领域具有潜在的实际应用方面的价值,包括与人体健康相关的临床应用价值,与机器智能化仿生研究及开发相关的价值等,例如,在医学超声诊断和航空目标探测方面,同样存在目标的识别、区分和锁定等,若研制出的临床超声探测诊断仪器的空间分辨率能达到微米级,则有可能分辨出单个机体细胞的病变,其意义不言而喻,在农业机器人领域,对植物的探测分类可用以合理控制和除去杂草等。
猜你喜欢
电子产品世界(2021年8期)2021-01-16
小资CHIC!ELEGANCE(2021年46期)2021-01-11
睿士(2020年11期)2020-11-16
VOGUE服饰与美容(2020年9期)2020-09-02
VOGUE服饰与美容(2020年2期)2020-02-16
中国计算机报(2019年49期)2019-02-07
中国新闻周刊(2017年36期)2017-10-21
小溪流(画刊)(2016年12期)2017-02-04
创新时代(2016年8期)2016-10-21
微型小说选刊(2015年5期)2015-06-05
