
ZK93426 促进小鼠精子获能相关蛋白酪氨酸磷酸化的机制研究*
2022-09-18 15:15王晓琛
中国男科学杂志 2022年4期
马 婕 王晓琛 刘 强
上海交通大学医学院 组织胚胎学与遗传发育学系(上海 200025);上海市生殖医学重点实验室(上海 2000025)
哺乳动物新鲜射出的精子必须在女性生殖道完成获能和顶体反应才具备受精的能力,精子获能的主要标志为1)细胞膜胆固醇流失,细胞膜通透性加强[1];2)胞内pH 升高[2];3)蛋白酪氨酸磷酸化水平升高[3];4)胞内钙离子浓度增加[4];5)精子发生超激活运动[5];6)细胞质膜超极化[6]等。 尽管关于获能相关的分子机制研究已经取得很大进展,但具体机制还亟待深入研究。
GABAA受体是中枢神经系统受抑制性神经递质γ-氨基丁酸(γ-aminobutyric acid,GABA)调节的离子型受体,哺乳动物中有19 种编码GABAA受体亚基的基因,包括α1-6,β1-3,γ1-3,ρ1-3,δ,ε,θ 和π[7]。 GABAA功能性受体通常包含5 个亚基,5 个亚基围绕使中央形成一个Cl-和HCO-3选择性的离子通道[8]。 众多的亚基导致体内可能存在数百个GABAA受体亚型,神经系统中最丰富的亚型是α1β2γ2[9]。 GABAA受体上有一位点可结合苯二氮卓类药物,该位点被称为苯二氮卓类受体(benzodiazepine receptor,BZR)。 GABAA受体中的αx+/γy-(x=1,2,3,5,y=1-3)亚基交界面是苯二氮卓类药物的高亲和位点[10]。 在生殖系统中,已有研究发现GABA 可以在体外促进哺乳动物获能相关蛋白酪氨酸磷酸化和顶体反应的发生[11-14]。 在男性精浆[15],女性输卵管、卵巢等组织[16]中均发现了GABA 的存在,在人[17],小鼠[12],大鼠[18]等精子中,也发现存在GABAA亚基。 但GABAA受体在精子中的表达模式和发挥功能的具体机制还有待研究。
ZK93426 是一类β-咔啉衍生物,是高效的,选择性和竞争性的苯二氮卓类受体拮抗剂[19],具有微弱的反向激动效应。 之前的研究中,我们发现苯二氮卓受体拮抗剂ZK93426 可促进小鼠和人精子获能相关蛋白酪氨酸磷酸化升高,并且依赖于获能培养液中的和BSA 的存在,使用蛋白激酶A(protein kinase A,PKA)抑制剂H89 可抑制ZK93426 的促进作用,表明ZK93426作用于PKA 上游发挥其促进功能[20]。 本文中我们研究了ZK93426 对精子生理指标的影响,以期揭示其促进蛋白酪氨酸磷酸化的机制。
材料与方法
一、试剂与仪器
(一)主要试剂
ZK93426 购于Tocris;二甲基亚砜(DMSO)、精子培养缓冲液(modified Krebs-Ringer medium,HM)、精子获能缓冲液(modified Krebs-Ringer bicarbonate medium,HMB)相关配制试剂购自Sigma-Aldrich;Fluo4-AM,Pluronic F-127 (0.04% w/v),DiSC3(5)(3,3'-二丙基噻二碳青碘化物)购自Invitrogen;BCECF-AM[2,7'-双-(2-羧乙基)-5-(和-6)-羧基荧光素,乙酰甲氧基甲酯],MQAE[N-(乙氧基羰基甲基)-6-甲氧基溴化喹啉]购自碧云天生物技术有限公司;Valinomycin,CCCP(羰基氰化物3-氯苯腙)购于MedChem Express;考马斯亮蓝G-250购自国药集团化学制剂有限公司。
(二)主要仪器
恒温培养箱(Heal Force);计算机辅助精液分析系统(Computer-Aided Sperm Analysis, CASA)(Hamilton Thorne);Gemini XPS 荧光读板仪(Molecular Devices);CytoFlex S 流式细胞检测仪(BeckMan)。
二、实验方法
(一)小鼠精子的获得
小鼠精子培养缓冲液HM 及获能缓冲液HMB 按照Pietrobon 等人所描述的方法配制[21]。
根据上海交通大学医学院IACUC(Institutional Animal Care and Use Committee)的指导准则对实验小鼠C57BL/6 进行安乐死。 取小鼠附睾尾部于37℃预热的HM 培养液中,并将其剪碎,37℃,5% CO2的条件下,使附睾尾部精子上游。 15min 后,弃组织碎片,并用HM清洗精子2 次。 使用HM 重悬精子,并调整浓度为2×107/mL。
(二)精子胞内钙离子浓度[Ca2+]i测定
[Ca2+]i的变化通过装载离子探针Fluo4-AM,并通过流式细胞仪进行测定。 精子悬液在Pluronic F-127(0.04% w/v)的存在下,与终浓度为2μM 的Fluo4-AM探针于37℃孵育20min,多余的染料通过离心洗去,重悬于HM 培养液中,并于37℃孵育30min 以去酯化,Fluo-4 AM 进入细胞后可以被细胞内的酯酶剪切形成Fluo4,从而被滞留在细胞内。 500×g 离心5min,去上清,将精子重悬于HMB 中,调整浓度为0.5-1×107/mL,加入50μM ZK93426 或DMSO 或10μM A23187 于37℃,5% CO2条件下孵育30min,于第28min 时,加入2ng/μL PI 染色2min。 随后立即使用流式细胞仪检测[Ca2+]i变化。 A23187 为钙离子载体,可促进钙离子大量内流,因此作为阳性对照。
(三)精子胞内氯离子浓度[Cl-]i测定
精子悬液与5 mM MQAE 于37℃孵育45min,离心洗去多余染料,将精子重悬于HMB 中,调整浓度为0.5 ~1×107/mL。 荧光读板机测定355/460nm 处的荧光值,待荧光值稳定后加入DMSO 或50μM ZK93426 通过ΔF/F0 显示[Cl-]i水平的变化。 ΔF 为加入药物前后的荧光值差,F0 为加入药物前的平均基础荧光值。
(四)精子pHi测定
精子与终浓度为3μM 的BCECF-AM 于37℃共孵育30min,离心洗去多余染料,重悬于HM 缓冲液中并于37℃去酯化30min,500×g 离心5min 后将精子重悬于HMB 中,随后上机测定。 用490nm 和440nm 波长激发BCECF 所得到的发射荧光之比与pH 有很好的线性关系。 待荧光值稳定后加入DMSO 或ZK93426。 最终通过ΔR/R0 显示胞内pH 的变化,ΔR 为加入药物前后的荧光比值差,R0 为加入药物前的平均基础荧光比值。
(五)精子细胞膜电位Em 测定
通过荧光染料DiSC3(5)测定精子细胞膜电位的改变。 当细胞膜超极化时,染料进入细胞,与细胞内蛋白质结合后淬灭其荧光,并在其光谱中表现出轻微的偏移,其荧光信号降低。 将精子细胞重悬于HMB 培养液中,加入50μM ZK93426 并于体外获能30,60,90min,并分 别 在 第 22, 52, 82min 时( 提 前 8min) 加 入DiSC3(5)(1μM)孵育5min 后,加入终浓度为1μM 的CCCP 孵育3min,使用荧光读板机监测620/670nm 下荧光值变化,通常1 ~2min 荧光值达到稳定。 通过添加1μM Valinomycin 和不同浓度的KCl 来校正Em。 假设[K+]i为120mM,根据能斯特方程,初始KCl 浓度为4.77mM,对应Em 值为-86mV,额外加入KCl 使其终浓度为8、12、20 和36mM,分别对应于-72,-61,-47 和-32mV的质膜电位。 非获能组精子细胞重悬于HM 中,其余操作相同。
(六)统计分析
使用GraphPad Prism 8 进行统计学分析及绘图。曲线图使用双因素方差分析(two-way ANOVA)进行统计学分析,多组之间的比较使用Bonferroni 校正。 柱状图使用单因素方差分析(one-way ANOVA)进行统计分析。 对仅有两组的数据使用T检验进行统计分析。 所有数据以Mean±S. E. M 表示。 当p<0.05 时认为具有统计学差异,*p<0.05,**p<0.01,***p<0.001。 所有实验至少重复3 次。
结 果
一、ZK93426 促进小鼠获能过程中精子[Ca2+]i升高
通过荧光探针Fluo4-AM 检测小鼠精子[Ca2+]i变化。结果表明,与DMSO 对照相比,小鼠精子与ZK93426于获能培养液中共孵育30min 后,精子[Ca2+]i显著升高(图1)。

图1 ZK93426 处理导致小鼠获能过程中精子[Ca2+]i升高
A-C:PI 荧光与Fluo4 荧光流式散点图及直方图:根据PI荧光值区分死(第一、二象限)、活(第三、四象限)细胞群体,在活细胞群体中根据Fluo-4 荧光强度区分低(直方图左侧)或高(直方图右侧)Fluo-4 荧光值细胞所占比例。 分别表示DMSO(A)、ZK93426(B)、A23187(C)处理30min 精子[Ca2+]i改变。 D. ZK93426 处理组与DMSO 溶剂对照组和阳性对照组A23187 的高Fluo-4 荧光群体百分比对比统计图。n=3,***p<0.001。
二、ZK93426 处理导致小鼠精子获能过程中[Cl-]i降低
通过氯离子探针MQAE 检测了小鼠精子[Cl-]i的改变。 MQAE 的检测灵敏度高,是一种目前应用最广泛的新型氯离子探针,当[Cl-]i升高时,MQAE 的荧光强度随着氯离子浓度的增加而成比例地减少。 我们使用荧光读板机实时检测小鼠精子获能过程中[Cl-]i的改变,可见加入ZK93426 后,精子[Cl-]i显著降低(图2)。

图2 ZK93426 处理导致小鼠精子[Cl-]i降低
A.MQAE 测定精子胞内氯离子浓度改变,可见ZK93426处理后,精子细胞胞内荧光升高,表示[Cl-]i降低。 B.ZK93426处理组与溶剂对照组(DMSO)的MQAE 荧光值统计比较。图2A 中箭头指示加入药物的时间点。n=6,***p<0.001。
三、ZK93426 处理加速小鼠精子获能过程中胞内碱化
通过pH 探针BCECF-AM 实时监测小鼠精子pHi的变化。结果显示,加入ZK93426 可在5min 内快速促进pHi升高(图3)。
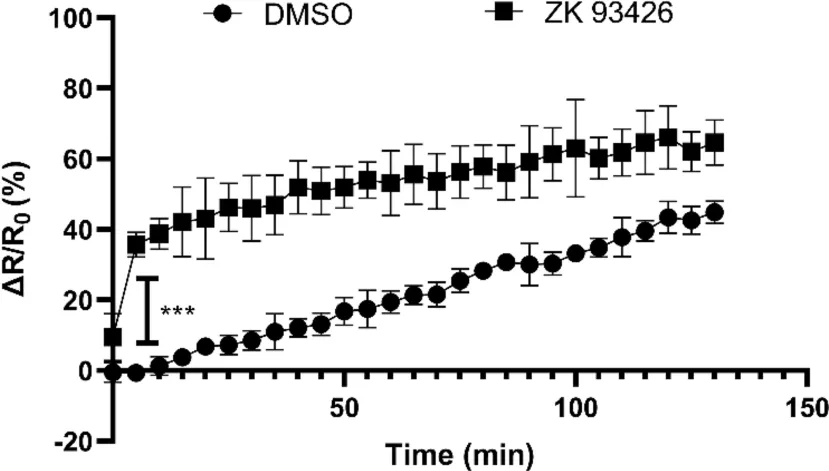
图3 ZK93426 处理导致小鼠精子获能过程中pHi升高
使用BCECF-AM 测定小鼠精子pHi变化,可见ZK93426促进小鼠精子获能过程中胞内碱化。n=4,***p<0.001。
四、ZK93426 抑制小鼠精子获能过程中细胞膜超极化
通过荧光探针DiSC3(5)检测了小鼠精子细胞膜电位改变。 在小鼠精子悬液中加入ZK93426 或DMSO 于37℃获能30,60,90min 后,检测精子细胞膜电位。结果显示,小鼠精子获能过程中细胞膜逐渐超极化(图4A、B 和D),但ZK93426 可抑制获能过程中细胞膜的超极化(图4C 和D)。
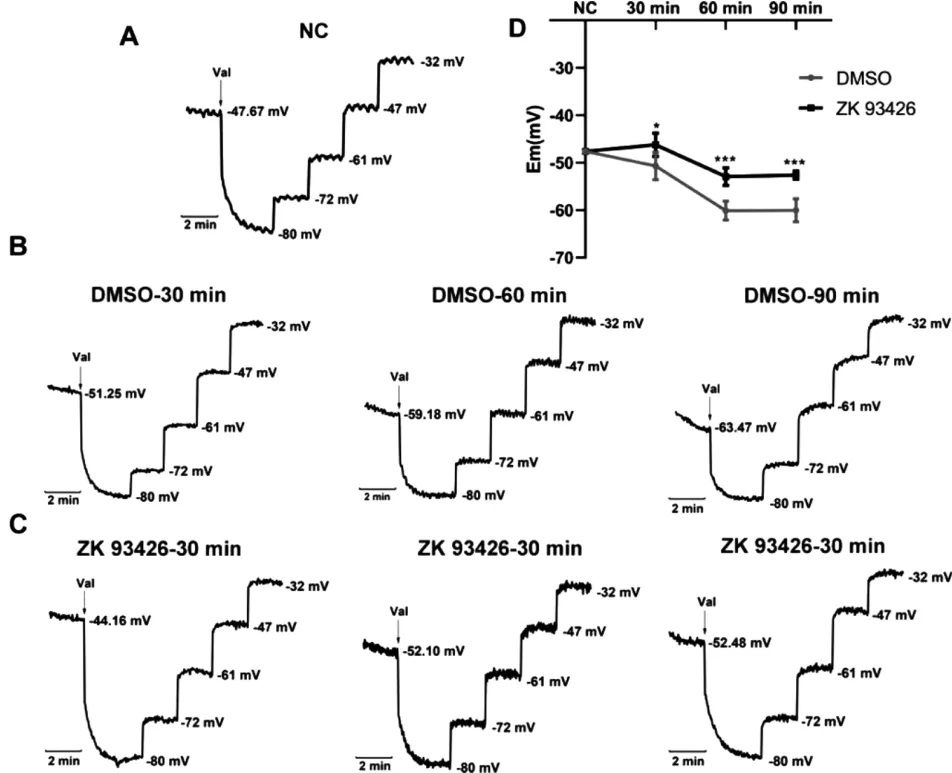
图4 ZK93426 抑制小鼠精子获能细胞膜超极化
A.非获能培养液中,小鼠精子细胞膜电位。 B-C:获能培养液中,小鼠精子与ZK93426(C)或DMSO(B)孵育30,60,90min 后,小鼠精子的膜电位值。 D. 细胞膜电位统计图,n≥3,*p<0.05,***p<0.001。
讨 论
精子睾丸后成熟依赖于男性和女性生殖道的微环境,附睾尾部是一个低Na+,Cl-,HCO-3的弱酸性环境,这种环境对于维持精子静止状态至关重要,当精子进入女性生殖道后,精子暴露于高Cl-,HCO-3,Ca2+的环境中,这些离子流入精子细胞膜后会激活cAMP/PKA 级联化反应,并且诱导了胞内碱化,蛋白酪氨酸磷酸化的发生及细胞膜的超极化,并最终使精子完成获能并获得受精能力[2]。
本文延续之前的研究,对ZK93426 促进的蛋白酪氨酸磷酸化的机制进行了进一步的探究。 HCO-3和Ca2+是调控精子获能相关蛋白酪氨酸磷酸化的两个重要离子,Ca2+可以增加ATP 对sAC:可溶性腺苷酸环化酶(Soluble adenylate cyclase,sAC)的亲和性,并促进sAC 对HCO-3的敏感性[22]。 HCO-3可以激活sAC 以产生大量cAMP 来激活PKA,从而激活下游一系列级联反应最终导致蛋白酪氨酸磷酸化水平升高[23]。 其中HCO-3在获能期间的内流又会导致胞内pH 的升高,胞内的碱化又会激活pH 依赖性钙离子通道CatSper 及钾离子通道Slo3 的开放,进而分别调控胞内钙离子浓度及细胞膜电位的变化[23]。 我们的结果表明ZK93426 快速促进小鼠精子pHi升高,并导致[Ca2+]i水平显著升高。 考虑到之前的研究中我们发现ZK93426 促进的蛋白酪氨酸磷酸化升高是依赖于HCO-3且ZK93426 作用于PKA 上游发挥功能,我们推测ZK93426 极有可能是通过促进HCO-3内流从而快速激活cAMP/PKA 通路来促进蛋白酪氨酸磷酸化的升高。 此外,ZK93426 促进的胞内碱化也可能激活了CatSper 通道,从而造成[Ca2+]i升高。
我们的结果还表明ZK93426 处理后[Cl-]i降低。Cl-在调控蛋白酪氨酸磷酸化中也发挥了重要功能。 当培养基中不添加Cl-时,获能相关蛋白酪氨酸磷酸化无法发生,加入cAMP 类似物可以恢复获能相关蛋白酪氨酸磷酸化,表明Cl-作用在cAMP/PKA 上游发挥功能[24]。 此外,有研究发现,孕酮处理人精子也会导致瞬时胞内Cl-的外流,并且加入GABAA受体抑制剂后,Cl-外流受到抑制[25]。 Cl-也是激活大鼠GABAA受体必不可少的离子[26]。 这提示我们Cl-外流在ZK93426 促进的蛋白酪氨酸磷酸化中可能也发挥了重要功能。
由于ZK93426 处理导致了[Cl-]i降低,细胞膜电位也受到了影响。 我们推测ZK93426 促进Cl-外排导致的细胞去极化也可能激活了某些电压钙离子通道(voltage-gated Ca2+channel,Cav)从而导致[Ca2+]i增加。已知弱去极化可打开Cav3通道,而强去极化可打开Cav1和Cav2通道[27]。 尽管早期研究中无法检测到附睾尾成熟精子的Cav 电流,但是后续的研究认为是由于精子的成熟状态和技术限制造成的无法检测,并且证明了Cav2.3参与调控精子顶体反应和钙离子内流[28]。
综上所述,本文的结果表明ZK93426 可能通过调控[Ca2+]i,[Cl-]i,pHi及Em 来共同调节小鼠精子获能相关蛋白酪氨酸磷酸化的升高。 该结果可为精子获能的机制提供一定的理论基础。
猜你喜欢
心肺血管病杂志(2020年5期)2021-01-14
中成药(2018年12期)2018-12-29
天然产物研究与开发(2018年1期)2018-02-02
中成药(2018年1期)2018-02-02
中成药(2017年9期)2017-12-19
中成药(2017年5期)2017-06-13
中国塑料(2016年7期)2016-04-16
华南农业大学学报(2015年5期)2015-12-04
中国医疗美容(2015年4期)2015-04-27
科学中国人(2015年13期)2015-02-28
