
鼠疫耶尔森菌小肽基因sORF34缺失株毒力及短期免疫保护效果评价
2022-10-19 06:18曹诗洋谭亚芳周亚洲陈红艳任一帆杨瑞馥杜宗敏
中国人兽共患病学报 2022年9期
郭 晓,曹诗洋,谭亚芳,周亚洲,陈红艳,任一帆,王 艳,杨瑞馥,杜宗敏
鼠疫是由鼠疫耶尔森菌(Yersiniapestis,简称鼠疫菌)感染引起的自然疫源性人兽共患病,主要宿主是啮齿动物,可通过鼠蚤传染给人(腺鼠疫),并可通过气溶胶途径在人与人之间传播(肺鼠疫)。鼠疫传染性强、病死率高[1],也是最具威胁的生物战剂和生物恐怖制剂之一[2]。疫苗接种是预防鼠疫最为经济有效的方法,但目前尚未有毒副作用小、保护效果好的疫苗问世。
目前鼠疫疫苗主要有灭活疫苗、重组亚单位疫苗、活载体疫苗、减毒活疫苗等。其中,鼠疫EV76减毒活疫苗在中国、蒙古和前苏联部分地区被用在高危人群中,它是基于色素沉着位点(pigmentation locus,pgm)缺失的减毒株,能够诱导机体的体液免疫和细胞免疫应答[3-4],对腺鼠疫和肺鼠疫能够起到预防作用。但是鼠疫EV76减毒活疫苗可引起不同程度的副作用。此外,有其他pgm-减毒株感染造成实验人员死亡的个案报道[5]。因此,本研究致力于进一步降低已有减毒株的残存毒力,并使其具有良好的保护率。
一般而言,细菌基因组编码的长度小于50个氨基酸的产物称为小肽(small ORF encoded peptide,SEP),由于在基因组注释的过程中被漏掉而长期被忽视。近年研究发现,小肽在真核生物和原核生物中都发挥着重要的功能[6-7]。本研究组在研究鼠疫菌小肽组学的过程中,鉴定出一个新型小肽,编号为SEP34,其基因位于鼠疫菌91001染色体YP1841基因下游,即2 543 689~2 543 865 bp处,暂命名为sORF34,在敲除编码基因后鼠疫菌毒力明显下降[8]。
本研究针对sORF34基因,分别以田鼠型鼠疫菌201株和EV76疫苗株构建了sORF34缺失株,并评价其经皮下感染小鼠的毒力变化。采用皮下免疫方式评价sORF34缺失株诱导机体免疫应答情况和对鼠疫不同感染途径攻毒的短期保护效果,探讨鼠疫EV76ΔsORF34株作为减毒活疫苗候选株的可行性。
1 材料与方法
1.1 材 料
1.1.1 菌种、质粒及实验动物 鼠疫菌201株(本室保存),鼠疫菌EV76疫苗株(本室保存),质粒pKD46、pKD4、pCP20(本室保存),SPF级6~8周龄雌性BALB/c小鼠(北京维通力华实验动物技术有限公司提供,许可证号:SCXK(京)2016-0006)。
1.1.2 试剂 氨苄西林(上海生工生物工程有限公司)、卡那霉素(MP,美国)、BHI培养基(BD公司,美国)、LB培养基(实验室自制)、溶血赫氏琼脂培养基(青岛海博生物技术有限公司)、DNA Maker 2000 plus(北京博迈德基因技术有限公司)、QIAquick PCR Purification Kit(Qiagen公司,德国)、QIAquick Gel Extraction Kit(Qiagen公司,德国)、Taq DNA polymerase(北京博迈德基因技术有限公司)、L-阿拉伯糖(上海生工生物工程有限公司)、天然F1(Fraction 1)蛋白(本室制备并保存)、LcrV(Low calcium response V)重组蛋白(本室制备并保存)、HRP-羊抗小鼠IgG(H+L)抗体(Thermofisher公司,美国)、HRP-羊抗小鼠IgG1抗体(Thermofisher公司,美国)、HRP-羊抗小鼠IgG2a抗体(Thermofisher公司,美国)、HRP-羊抗小鼠IgG2b抗体(Thermofisher公司,美国)ELISA辅助试剂盒(北京达科为生物技术股份有限公司),引物均合成自上海生工生物技术有限公司。
1.1.3 仪器 HZQ-F160全温震荡培养箱(太仓市实验设备厂)、T100 PCR仪(Bio-Rad公司,美国)、UV-5100B分光光度计(上海元析仪器有限公司)、Gene Pluser Xcell 电转仪(Bio-Rad公司,美国)、核酸电泳仪(北京六一仪器厂)、GelDoc Go自动成像仪(Bio-Rad公司,美国)、iMark酶标仪(Bio-Rad公司,美国)。
1.2 方 法
1.2.1 小肽基因缺失株的构建 使用λ-Red一步突变法构建小肽基因缺失株[9]。首先设计并合成同源区突变盒引物sORF34-P1/P2(表1),以pKD4质粒为模板,对卡那抗性基因进行扩增获得突变盒(预变性94 ℃/5 min;变性94 ℃/40 s,退火54 ℃/40 s,延伸72 ℃/2 min 30 s,30个循环;再延伸72 ℃/5 min),回收PCR产物。之后将pKD46质粒分别电转入鼠疫201株和鼠疫EV76疫苗株,L-阿拉伯糖诱导重组蛋白表达,制备感受态细胞。将1.5 μg 卡那抗性突变盒扩增产物电转入感受态细胞,加入1 mL LB培养基复苏2 h,集菌后全部涂布于双抗LB平板(氨苄西林100 μg/mL,卡那霉素50 μg/mL)培养2 d,挑取单菌落到双抗板上划线增菌,sORF34-F/R(表1)引物进行PCR鉴定(预变性95 ℃/5 min;变性95 ℃/40 s,退火58 ℃/40 s,延伸72 ℃/2 min 30 s,30个循环;再延伸72 ℃/5 min),并测序验证克隆正确。在37 ℃恒温培养箱中培养1 d,100 μL菌液稀释1 000倍后,涂LB平板,挑取单菌落使用pKD46-JD-F/R(表1)引物进行PCR鉴定(预变性95 ℃/5 min;变性95 ℃/40 s,退火56 ℃/40 s,延伸72 ℃/1 min,30个循环;再延伸72 ℃/5 min),筛选pKD46质粒消除克隆。获得消除pKD46菌株后,再电转入pCP20辅助质粒,导入质粒的单克隆于37 ℃培养1 d,100 μL菌液稀释1 000倍后涂布LB平板,挑取单克隆,sORF34-F/R引物进行PCR确定抗性基因消除,测序筛选正确克隆。获得最终的sORF34基因敲除株,命名为201ΔsORF34、EV76ΔsORF34。
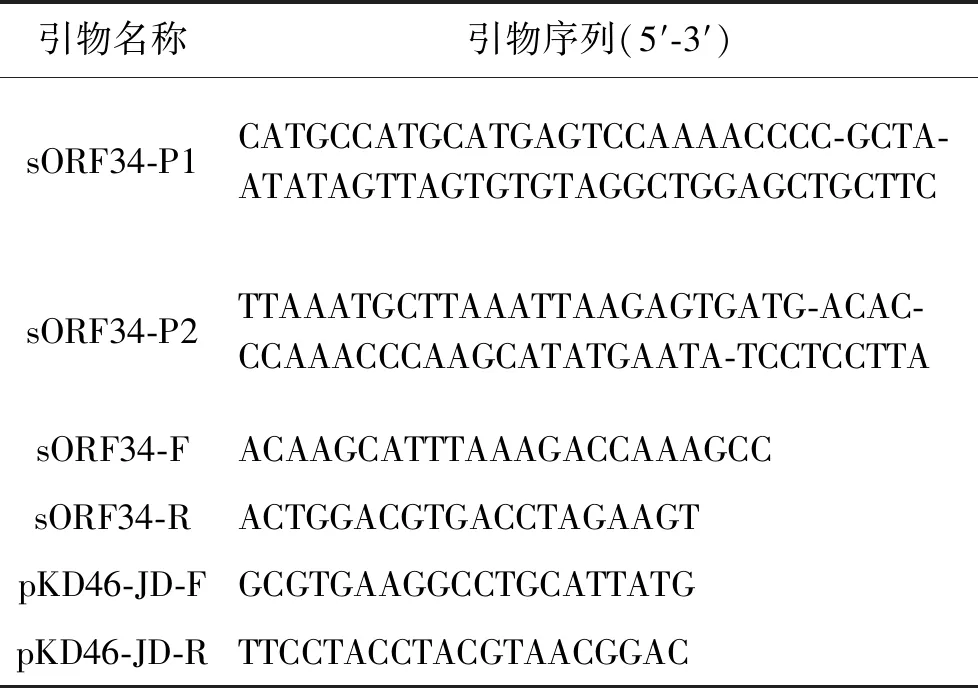
表1 实验所用引物Tab.1 Primers used in this study
1.2.2 细菌培养 鼠疫菌201株、鼠疫菌EV76疫苗株、基因缺失株201ΔsORF34和EV76ΔsORF34均转接三代培养。甘油种接种于5 mL的LB培养基中,26 ℃、200 r/min过夜培养。上述菌液1∶20接种于5 mL LB 培养基,培养至OD620≈1.0,而后1∶20接种于5 mL LB培养基,培养至对数中期(OD620≈1.0),获得最终培养菌液。滴鼻攻毒前将最终培养菌液移至37 ℃孵育1 h。
1.2.3 鼠疫201ΔsORF34株皮下攻毒途径感染小鼠LD50测定 使用无菌磷酸缓冲盐溶液(PBS)将鼠疫201ΔsORF34菌液调整为 OD620≈1.0理论浓度为2×107CFU/100 μL后,10倍稀释至2×102~2×106CFU/100 μL。选取SPF级6到8周龄雌性BALB/c小鼠50只,随机分成5组,每组10只,取100 μL各梯度菌液进行皮下感染,滴板计数确定实际感染量。感染后每天观察小鼠2次,记录死亡情况,连续观察14 d,计算不同感染剂量小组的小鼠死亡情况,使用Reed-Muench法[10]计算鼠疫201ΔsORF34皮下感染途径半数致死量(LD50)。相同方法,调整鼠疫菌201株菌液浓度为1~1×105CFU/100 μL,每组小鼠5只进行试验并计算出鼠疫201株皮下感染途径LD50。
1.2.4 皮下感染途径鼠疫EV76ΔsORF34株生存曲线观察 6~8周龄雌性BALB/c小鼠20只,随机分成两组,每组10只。使用无菌PBS将鼠疫EV76疫苗株和EV76ΔsORF34株菌液配制成理论感染剂量2×108CFU/mL,取100 μL以皮下途径(s.c.)感染小鼠,滴板计数确定实际感染量。每天观察记录2次小鼠发病及死亡情况,连续观察2周,绘制生存曲线。
1.2.5 动物免疫与攻毒 6到8周龄雌性BALB/c小鼠60只,随机分组进行腹股沟皮下注射免疫接种。鼠疫201ΔsORF34株(20只)、EV76ΔsORF34株(20只)、EV76株(10只)免疫剂量分别为2×103CFU/100 μL、2×105CFU/100 μL、2×105CFU/100 μL,对照组(10只)注射100 μL PBS溶液,21 d后上述菌株分别以2×103CFU/100 μL、2×105CFU/100 μL、2×105CFU/100 μL剂量加强免疫,对照组注射100 μL PBS溶液。首次免疫后42 d,存活小鼠使用鼠疫201株进行皮下攻毒和滴鼻攻毒,攻毒理论剂量分别为1×103CFU/只(300 LD50)和1×105CFU/只(40 LD50)。攻毒后每天观察记录2次小鼠发病及死亡情况,连续观察2周,计算生存率。
1.2.6 ELISA检测血清抗体水平 免疫小鼠于初免后40 d眼眶静脉采血,分离血清备用。1 μg/mL纯化F1抗原蛋白、2 μg/mL重组鼠疫低钙响应因子V(LcrV)抗原每孔100 μL加入ELISA板中,4 ℃过夜包被ELISA板,封闭液37 ℃封闭2 h,血清分别从1∶200到1∶409 600、1∶20到1∶2 560连续2倍稀释,100 μL不同稀释度血清加入检测孔,37 ℃孵育30 min,PBST洗液洗板5次;检测孔分别加入HRP-羊抗小鼠IgG抗体、HRP-羊抗小鼠IgG1抗体、HRP-羊抗小鼠IgG2a抗体、HRP-羊抗小鼠IgG2b抗体,37 ℃孵育20 min,PBST洗液洗板5次;检测孔加入TMB 37 ℃避光孵育10 min;加入终止液,酶标仪检测450 nm处OD值。大于2.1倍阴性对照组OD值判为阳性。
1.2.7 统计分析 用GraphPad Prism 8.0.1软件进行数据分析,在皮下途径感染小鼠生存曲线和保护效果评价实验中,组间比较采用Log-Rank(Mantel-Cox)检验;在保护性抗体滴度比较实验中,采用方差分析(ANOVA)进行多组间比较,使用Tukey检验进行事后两两比较;检验水准α=0.05。
2 结 果
2.1 小肽基因sORF34缺失株的鉴定 利用1.2.1中用于鉴定sORF34基因的引物对sORF34-F/R对缺失株和野生株进行菌落PCR扩增,扩增产物通过1.5%琼脂糖凝胶电泳进行检测。敲除株和野生株理论PCR产物大小分别为248 bp和316 bp。图1和图2为对所构建的阳性敲除菌株的PCR鉴定结果,条带大小、产物测序结果均符合预期,表明小肽基因缺失株201ΔsORF34、EV76ΔsORF34构建成功。

图1 201ΔsORF34基因缺失菌株PCR鉴定结果Fig.1 Electrophoresis of PCR verification of Y.pestis 201 strain sORF34 gene deletion mutant

图2 EV76ΔsORF34基因缺失菌株PCR鉴定结果Fig.2 Electrophoresis of PCR verification of Y.pestis EV76 strain sORF34 gene deletion mutant
2.2 皮下途径感染201株与201ΔsORF34的LD50比较 鼠疫201株实际感染剂量分别为0.46~4 600 CFU/只,201ΔsORF34实际感染剂量分别为3.24×102~3.24×106CFU/只,通过皮下途径感染BALB/c小鼠,201ΔsORF34株14 d观察期结束后,根据不同感染剂量组小鼠死亡情况绘制出生存曲线(图3),并使用Reed-Muench法[10]求出两株菌皮下途径感染BALB/c小鼠的LD50。计算结果,201株的LD50为3.1 CFU,201ΔsORF34的LD50为 4.75×104CFU,约是其亲本株1.5×104倍。由此可见sORF34基因缺失引起了鼠疫201株毒力的降低。

图3 不同剂量鼠疫菌201株(A)与201ΔsORF34(B)皮下感染小鼠生存曲线Fig.3 Survival curves of mice s.c.challenged with Y. pestis 201(A) or 201ΔsORF34(B) at different doses
2.3 皮下途径感染EV76和 EV76ΔsORF34后的生存曲线比较 将EV76疫苗株及EV76ΔsORF34株通过皮下途径免疫接种BALB/c小鼠,滴板计数结果显示EV76疫苗株实际感染剂量为2.5×107CFU/只,EV76ΔsORF34株实际感染剂量为3.1×107CFU/只。接种后观察14 d,绘制生存曲线(图4)。EV76感染组生存率为70%,EV76ΔsORF34 感染组生存率为100%。Log-Rank(Mantel-Cox)检验,两组生存率差异有统计学意义(P=0.035),且实际攻毒剂量EV76ΔsORF34较EV76株更高,可见EV76ΔsORF34较其亲本株毒力进一步下降。

①:P<0.05图4 EV76疫苗株与EV76ΔsORF34株皮下感染途径小鼠生存曲线Fig.4 Survival curves of mice s.c.challenged with high doses of Y. pestis EV76 or EV76ΔsORF34
2.4 减毒株皮下免疫后对皮下、滴鼻攻毒的保护效果评价 鼠疫201ΔsORF34株、EV76ΔsORF34株、EV76株采取皮下途径进行两剂免疫,间隔21 d,PBS溶液注射组为对照。在首剂免疫后42 d,201ΔsORF34株免疫组存活18只,EV76ΔsORF34株免疫组存活20只,EV76株免疫组存活10只,PBS组存活10只。存活小鼠随机均分为两组,用鼠疫201株皮下和滴鼻途径对各免疫接种组和PBS组进行攻毒,观察14 d,绘制生存曲线(图5)。皮下途径攻毒中201ΔsORF34株免疫组、EV76 ΔsORF34株免疫组与EV76疫苗株免疫组动物均100%存活;滴鼻途径攻毒中201ΔsORF34、EV76ΔsORF34免疫组与EV76疫苗株免疫组生存率分别为55.6%、40%、40%,能够起到部分保护作用,Log-Rank(Mantel-Cox)检验,3组保护率差异无统计学意义(P=0.699)。

A:300 LD50 Y.pestis 201皮下攻毒;B:40 LD50 Y.pestis 201滴鼻攻毒;①:P>0.05图5 3种鼠疫菌减毒株免疫小鼠后行皮下、滴鼻途径感染的生存曲线Fig.5 Survival curves of mice immunized with EV76,EV76ΔsORF34 or 201ΔsORF34 after exposure to Y. pestis 201 by subcutaneously or intranasal route
2.5 皮下免疫后小鼠血清保护性抗体滴度比较 鼠疫菌的荚膜抗原F1和毒力抗原LcrV是鼠疫菌的2种重要保护性抗原[11],因此我们使用ELISA方法检测了相应的抗体滴度。小鼠初免后40 d,经眼眶静脉采血并分离血清。其中鼠疫201ΔsORF34株、EV76株、EV76ΔsORF34株免疫组可以刺激机体产生较高滴度的抗F1-IgG(图6A)。EV76免疫组和201ΔsORF34免疫组较 EV76ΔsORF34 免疫组抗F1-IgG滴度更高。EV76免疫组和201ΔsORF34免疫组未检测出抗LcrV-IgG,EV76ΔsORF34免疫组可检测出较低滴度的抗LcrV-lgG(图6B)。对特异性F1抗体进行IgG亚型检测(图7),EV76、EV76ΔsORF34、201ΔsORF34免疫组IgG1抗体滴度均较IgG2a、IgG2b抗体滴度高(FEV76=45.96,P<0.001;FEV76ΔsORF34=20.47,P<0.001;F201ΔsORF34=7.399,P<0.01),EV76 ΔsORF34免疫组中IgG2b抗体滴度较201ΔsORF34免疫组低(qIgG2b=3.875,P<0.05),而IgG1、IgG2a抗体滴度与201ΔsORF34免疫组差异无统计学意义(qIgG1=1.187,P>0.05;qIgG2a=0.297,P>0.05)。

A:抗F1-IgG抗体滴度;B:抗LcrV-IgG抗体滴度;①:P<0.05;②:P<0.01;③:P>0.05图6 3种鼠疫菌减毒株免疫后小鼠血清抗F1、LcrV IgG抗体滴度Fig.6 F1-IgG and LcrV-IgG antibodies titer in mice immunized with Y.pestis EV76,EV76ΔsORF34 or 201ΔsORF34

①:P<0.05;②:P<0.01图7 小鼠血清中特异性抗鼠疫菌F1抗体IgG 亚型Fig.7 IgG isotypes of antibodies specific to F1 antigen in mouse sera
3 讨 论
鼠疫是《中华人民共和国传染病防治法》规定的甲类传染病,自然疫源地面积广泛,致病性强,病死率高。在经济开发和交通便利的背景下,远距离传播鼠疫的风险增加。同时,鼠疫的多重抗生素耐药株已经出现[12-13],因此亟待研制更为安全有效的鼠疫疫苗。国内外基于F1和(或)LcrV抗原的亚单位疫苗大多已经进入临床实验[14-17],展现出了良好的保护效果。但是F1抗原阴性鼠疫菌对人依然强毒,LcrV蛋白在自然界具有多态性,导致亚单位疫苗不能覆盖所有致病菌。鼠疫减毒活疫苗由于携带多种抗原,理论上能够覆盖更多致病菌株,并且能激发机体细胞免疫,比亚单位疫苗具有明显优势。本研究旨在现有鼠疫EV76疫苗株的基础上,通过敲除sORF34基因,进一步降低其残存毒力,同时观察其对鼠疫菌感染的保护作用。
首先评价小肽基因缺失株的安全性。本研究组在之前的研究中发现鼠疫菌201株中sORF34基因在37 ℃培养下较26 ℃转录水平升高,敲除后鼠疫菌201株较野生株毒力下降明显[8]。此次研究使用λ-Red一步法,将鼠疫菌201株和EV76疫苗株中sORF34基因敲除,构建了鼠疫菌201ΔsORF34和EV76ΔsORF34。本研究测定了皮下感染途径鼠疫菌201株和201ΔsORF34株的LD50,分别为3.1 CFU和4.75×104CFU,鼠疫201ΔsORF34株毒力较鼠疫201株下降1.5×104倍。虽然鼠疫菌201株对人不致病[18],但有研究表明在非人灵长类动物中鼠疫201株残存毒力高于EV76疫苗株[19]。鼠疫201ΔsORF34株在非人灵长类中的安全性,仍然需要进一步研究。由于EV76疫苗株对小鼠皮下途径LD50约为6.3×107CFU[20],本研究选择2×107CFU不同菌株皮下感染,观察生存率,比较两者毒力差别,EV76ΔsORF34株生存率为100%,而EV76疫苗株生存率为70%,二者差异有统计学意义,表明敲除sORF34后EV76疫苗株残存毒力进一步下降。
有研究报道,鼠疫菌减毒活疫苗可诱导体液免疫和细胞免疫反应[21-22]。我们对免疫后小鼠血清中主要保护性抗体(F1抗体和LcrV抗体)进行了检测。发现鼠疫201ΔsORF34株、EV76株、EV76ΔsORF34株免疫组可以产生较高滴度的抗F1-IgG,EV76疫苗株免疫组与201ΔsORF34株免疫组比较,差异无统计学意义(P>0.05),均高于EV76ΔsORF34株免疫组(P<0.05);而鼠疫201ΔsORF34株免疫剂量较EV76疫苗株和EV76ΔsORF34株低,提示鼠疫201ΔsORF34株可能具有更好的免疫原性。EV76免疫组不能有效刺激机体产生抗LcrV-IgG,这与已有文献报道结论一致[23-25],而EV76ΔsORF34免疫组可产生较低滴度的抗LcrV-IgG。之后本研究组对IgG抗体不同亚型滴度进行了检测,发现不同免疫组中特异性IgG1抗体的滴度均高于IgG2a和IgG2b。IgG1抗体的产生与Th2细胞相关,而IgG2a、IgG2b抗体的产生与Th1细胞相关。这种差异提示机体在针对鼠疫减毒活疫苗免疫应答中Th2占主导。
综上所述,本研究揭示鼠疫EV76ΔsORF34株比EV76疫苗株毒力更弱,且短期免疫保护效果差异无统计学意义。为鼠疫EV76ΔsORF34株作为候选的鼠疫减毒活疫苗提供了依据。同时201ΔsORF34株相较于201株毒力下降,相较于EV76株有更好的免疫原性,但其安全性需要进一步研究。
利益冲突:无
引用本文格式:郭晓,曹诗洋,谭亚芳,等.鼠疫耶尔森菌小肽基因sORF34缺失株毒力及短期免疫保护效果评价[J].中国人兽共患病学报,2022,38(9):757-763.DOI:10.3969/j.issn.1002-2694.2022.00.109
猜你喜欢
浙江临床医学(2022年8期)2022-11-22
中国兽医学报(2022年4期)2022-06-17
天津农业科学(2022年5期)2022-05-31
昆明医科大学学报(2022年2期)2022-03-29
特别健康·下半月(2020年12期)2020-12-14
江苏农业科学(2019年14期)2019-09-23
中外医疗(2016年29期)2016-11-30
中国现代医生(2016年25期)2016-11-19
江苏农业科学(2016年4期)2016-06-14
中国当代医药(2014年23期)2014-09-25
