
RNAi沉默HvFTZ-F1对茄二十八星瓢虫幼虫存活及发育的影响
2022-11-18 08:26王亚洁刘卓琦李华丽潘慧鹏杨春晓
中国生物防治学报 2022年5期
王亚洁,刘卓琦,李华丽,潘慧鹏,杨春晓*
(1.华南农业大学植物保护学院/广东省生物农药创制与应用重点实验室,广州 510642;2.华南农业大学亚热带农业生物资源保护与利用国家重点实验室,广州 510642)
FTZ-F1(Fushi-tarazu factor 1)是昆虫体内一种典型的核受体转录因子,在昆虫的蜕皮、化蛹和成虫繁殖中起到至关重要的作用。FTZ-F1在昆虫多个组织中作为功能因子响应20E(20-hydroxyecdysone,20-羟基蜕皮酮)信号,并介导下游基因的表达[1]。20E前体ecdysone在前胸腺(prothoracic glands,PGs)中合成并分泌到血淋巴,随体液循环进入靶细胞,在细胞质内被细胞色素P450羟化酶Shade(Cyp314a1)催化生成20E[2]。20E调节昆虫的蜕皮及变态发育,是昆虫生长发育过程中的关键组分,由细胞色素P450单加氧酶基因、spook、phantom、disembodied(DIB)、shadow和shade等基因共同参与合成[3]。在完全变态昆虫中,20E通过与蜕皮激素受体(ecdysone receptor,ECR)/超螺旋体(ultraspiracle,USP)异源二聚体结合来激活broad complex(BRC)、E74(Ecdysone-induced protein 74)、E75(Ecdysone-induced protein 75)和E93(Ecdysone-induced protein 93)等蜕皮激素早期应答基因的表达[4,5]。这些早期应答基因编码的蛋白再激活HR3(hormone receptor 3)、HR4(hormone receptor 4)、HR38(hormone receptor 38)等中期应答基因的表达,随后这些中期应答基因编码的蛋白通过诱导核受体因子FTZ-F1再触发晚期基因的表达[5,6]。因此,核受体FTZ-F1在20E通路的信号转导中起着重要作用。沉默昆虫的FTZ-F1基因,昆虫体内20E相关基因的表达水平会受到影响。例如,在马铃薯叶甲Leptinotarsa decemlineata中,沉默LdFTZ-F1会影响幼虫化蛹,显著抑制20E相关基因的转录,并降低20E滴度[7]。在果蝇Drosophila melanogaster中沉默DmFTZ-F1会导致phantom、DIB、shadow表达量的降低[8]。
在蜕皮过程中,TH(tyrosine hydroxylase)、DDC(Dopa decarboxylase)、laccase2和ebony等色素合成基因调控昆虫体色的形成[9]。酪氨酸羟化酶(TH)将酪氨酸转化为多巴(Dopa);之后,多巴脱羧酶(DDC)将多巴转化为黑色素的前体多巴胺(Dopamine);laccase2通过多巴和多巴胺催化黑色素的产生[9-12]。合成的色素前体从表皮细胞中分泌出来,然后掺入表皮,形成可视化的黑色斑纹和斑点。多巴胺还可由ebony转化为N-β-丙烯基多巴胺(NBAD),参与表皮鞣化,使表皮呈现黄色或者淡黄色[13]。研究发现20E调控色素相关基因的表达,并且20E和保幼激素(JH)对昆虫体内总多巴胺水平具有特异性影响[11,14,15]。FTZ-F1首次在果蝇中发现,编码α型和β型两种蛋白质,已有研究证明βFTZ-F1参与调控昆虫蛹壳形成,是表皮形成所必须的[16]。沉默昆虫体内FTZ-F1基因会导致幼虫蜕皮失败并死亡。例如,将赤拟谷盗Tribolium castaneum5龄幼虫注射800~1000 ng dsTcFTZ-F1造成100%的幼虫在化蛹前死亡[17]。在黑腹果蝇Drosophila melanogaster中,βFTZ-F1调控蜕皮和变态相关基因,沉默DmFTZ-F1会抑制幼虫蜕皮直至死亡[18]。在德国小蠊Blattella germanica中,BgFTZ-F1控制蜕皮脉冲的时间,沉默BgFTZ-F1导致幼虫生长发育迟缓和蜕皮失败[19]。BmFTZ-F1与家蚕Bombyx mori幼虫蜕皮和羽化有关[20]。沉默棉铃虫Helicoverpa armigera4龄幼虫的HaFTZ-F1基因,导致幼虫蜕皮过程受到抑制[21]。以上研究都表明FTZ-F1在昆虫蜕皮过程中起着至关重要的作用。
茄二十八星瓢虫Henosepilachna vigintioctopunctata属于鞘翅目、瓢虫科,主要为害茄科植物,如马铃薯Solanum tuberosum、番茄Lycopersicon esculentum、茄子Solanum melongena等。自2015年起,我国推进马铃薯主粮化,近年来,随着马铃薯种植面积不断扩大,茄二十八星瓢虫的为害愈加严重[22]。目前,化学防治仍然是防控茄二十八星瓢虫的主要手段。然而由于化学药剂长期、不合理地使用会引起环境污染以及农产品质量安全等问题。因此,制定其他防控策略来对茄二十八星瓢虫进行综合治理非常有必要[23]。RNA干扰(RNA interference,RNAi)指外源或内源的双链RNA(doublestranded RNA,dsRNA)在生物体内被核酸内切酶 Dicer切割成多个具有特定长度和结构的小片段siRNA,之后通过序列配对的模式干扰靶标基因翻译蛋白过程或降解 mRNA序列,从而产生基因沉默的现象[24]。近十年,RNAi在害虫防治中的应用得到了广泛的研究[25-28]。RNAi已成为了通过抑制基因表达进行害虫生物防治的有效手段。以RNAi为基础的害虫防治是控制茄二十八星瓢虫的一个潜在可行的解决方案[28-32]。
为筛选基于RNAi技术的茄二十八星瓢虫绿色防控的候选靶标基因,通过饲喂茄二十八星瓢虫1龄幼虫100 ng/μL的dsHvFTZ-F1处理过的叶片,使得幼虫无法蜕皮进入2龄,直至死亡,死亡率为90%[32]。在本研究中,我们利用饲喂法RNAi,探究低浓度处理下沉默HvFTZ-F1基因对茄二十八星瓢虫存活和发育的影响,为降低防控成本、实现RNA生物农药产业化提供理论基础。
1 材料与方法
1.1 供试植物与虫源
本研究所用的茄二十八星瓢虫于2018年4月采集自华南农业大学校内的龙葵,将瓢虫与叶片均置于放有滤纸和棉球保湿的培养皿中,在人工气候箱中继代饲养。饲养条件:温度(25±1)℃,相对湿度70%~80%,光周期14L∶10D。
1.2 RNA的提取与cDNA的合成
采用TRIzol法提取样品总RNA,使用仪器NanoDrop OneC分光光度计(Thermo Fisher Scientific,Waltham,MA,USA)测定RNA浓度,所有样本RNA的OD260/OD230在1.8~2.2。使用试剂盒PrimeScriptTMRT reagent Kit with gDNA Eraser(TaKaRa,Dalian,China)将RNA反转录为cDNA。所得cDNA稀释10倍用于后续试验。
1.3 dsRNA的体外合成
使用包含T7启动子序列的特异性引物[33]进行HvFTZ-F1和绿色荧光蛋白(green fluorescent protein,GFP)dsRNA 的合成。PCR 反应体系:10 μL 5×PCR buffer,1.0 μL dNTP mix,5.0 μL 上游引物(10 μmol/L),5.0 μL 下游引物(10 μmol/L),0.25 μLTaq(5 U/μL)(TaKaRa,China)。PCR 反应程序:94 ℃预变性3 min;94 ℃变性30 s,55 ℃退火30 s,72 ℃延伸1 min,35个循环,72 ℃延伸10 min。PCR产物采用1%的琼脂糖凝胶电泳进行检测,使用 DNA胶回收试剂盒(Tiangen,Beijing,China)进行回收,作为合成dsRNA的模板。使用MEGAscript T7 RNAi试剂盒(Thermo Fisher Scientific,USA)合成dsRNA,分别得到dsHvFTZ-F1和dsGFP。使用1%琼脂糖凝胶电泳检测dsRNA质量并使用仪器NanoDrop OneC分光光度计(Thermo Fisher Scientific,USA)测定dsRNA浓度。
1.4 取食dsHvFTZ-F1对茄二十八星瓢虫1龄幼虫存活和发育的影响
用5 ng/μL的dsHvFTZ-F1溶液浸泡直径为12 mm的圆形茄子叶片1 min,待叶片自然风干后饲喂茄二十八星瓢虫1龄初幼虫,每隔24 h更换一次叶片,饲喂dsHvFTZ-F1处理的叶片48 h后,更换为新鲜的未经dsRNA处理的茄子叶片。将幼虫放入有滤纸和加湿棉球的培养皿中进行培养,用相同方法以5 ng/μL的dsGFP作为对照组。每个dsRNA浓度设置5个重复,每个重复含有10头1龄幼虫,培养皿置于人工气候箱中(温度25 ℃±1 ℃,相对湿度70%~80%,光周期14L∶10D)。每隔12 h统计培养皿中茄二十八星瓢虫的死亡数目,拍摄并记录dsRNA处理下茄二十八星瓢虫的存活和发育情况。
1.5 取食dsHvFTZ-F1对茄二十八星瓢虫1龄幼虫20E和色素沉积相关基因表达的影响
用5 ng/μL的dsHvFTZ-F1处理1龄幼虫。每10头幼虫为1个生物学重复,设置3个重复,处理后48 h收样。所有样本均用液氮迅速冷冻,用1.5 mL无RNA酶的离心管保存于-80 ℃,之后进行总RNA的提取并反转录为cDNA。所得cDNA用于检测dsHvFTZ-F1处理后对茄二十八星瓢虫1龄幼虫HvFTZ-F1基因、20E相关基因(HvSPOOK,HvSHADOW,HvSHADE,HvDIB,HvE75,HvECR,HvHR3)和色素沉积相关基因(HvDDC,HvTH,Hvebony)表达量的影响。
1.6 取食dsHvFTZ-F1对茄二十八星瓢虫1龄幼虫20E滴度的影响
为研究HvFTZ-F1沉默对20E滴度的影响,首先通过连续稀释标准20E溶液构建标准曲线。分别用5 ng/μL的dsHvFTZ-F1和5 ng/μL的dsGFP(对照组)处理1龄幼虫,每10头幼虫为1个生物学重复,设置3个重复,处理后48 h收样。所收样本以1∶9(w/v)比例加入PBS缓冲液(phosphate buffered saline,PBS),使用研磨器(Tiangen,China)在冰上充分研磨并稀释。所得匀浆以5000 r/min离心10 min,使用昆虫蜕皮酮酶联免疫吸附测定试剂盒(Insect Ecdysone ELISA Kit,MLBIO,China)测定上清液中20E滴度。使用酶标仪(BIO-RAD,CA,USA)测定光密度(optical densities,ODs)。
1.7 取食dsHvFTZ-F1对茄二十八星瓢虫1龄幼虫多巴胺滴度的影响
为研究HvFTZ-F1沉默对多巴胺滴度的影响,首先通过连续稀释标准多巴胺溶液构建标准曲线。分别用5 ng/μL的dsHvFTZ-F1和5 ng/μL的dsGFP(对照组)处理1龄幼虫,每10头幼虫为1个生物学重复,设置3个重复,处理后48 h收样。所收集样本以1∶9(w/v)的比例加入PBS缓冲液,使用研磨器在冰上充分研磨并稀释。所得匀浆以3000 r/min离心20 min,使用昆虫多巴胺酶联免疫吸附测定试剂盒(Insect Dopamine ELISA kit,NJJCBIO,China)对上层清液进行多巴胺浓度测定。使用酶标仪(iMark,BIO-RAD,USA)检测OD值。
1.8 RT-qPCR分析
对相关基因的表达模式进行RT-qPCR分析,RT-qPCR的反应体系:cDNA模板1.0 μL,引物F和R(10 μmol/L)各 0.625 μL,TB Green 7.5 μL,ddH2O 5.25 μL,共 15 μL。RT-qPCR 反应程序为 3 个阶段,分别为变性阶段95 ℃,30 s;定量分析阶段(95 ℃,5 s;60 ℃,30 s)40个循环;熔解曲线分析阶段(65 ℃~95 ℃,每升温0.5 ℃检测一次荧光信号,每次检测持续5 s)[33]。反应在96孔板Microseal PCR plates中进行,RT-qPCR的反应仪器为Bio-Rad C1000 Real-Time PCR system(Bio-Rad C1000 Real-Time PCR system,BIO-RAD,USA)。使用HvRPS18和HvRPL13作为内参基因,各基因的相对表达量用2-ΔΔCt法进行计算[34],所用引物序列参照之前研究[33]。
1.9 数据统计与分析
使用独立样本T检验分析HvFTZ-F1及相关基因在处理组和对照组中的表达水平差异,以及比较不同处理下 20E滴度和多巴胺滴度的差异(P<0.05)。百分比数据在统计分析前进行反正弦平方根变换。使用IBM SPSS Statistics v.24.0软件(IBM Corp.,Armonk,NY,USA)进行所有数据分析。
2 结果与分析
2.1 dsHvFTZ-F1处理对茄二十八星瓢虫1龄幼虫生存及发育的影响
经5 ng/μL的dsRNA处理后,dsGFP对照组幼虫在处理3 d后发育至2龄。dsHvFTZ-F1处理的幼虫无法蜕皮进入2龄直至死亡,并且所有幼虫的死亡状态均为后肠从肛门处伸出(图1)。

图1 茄二十八星瓢虫1龄幼虫经dsHvFTZ-F1处理3 d后表型的变化Fig.1 Phenotypic changes of the H.vigintioctopunctata 1st instar larvae treated with dsHvFTZ-F1 for 3 days
取食dsHvFTZ-F1处理的叶片,可导致茄二十八星瓢虫1龄幼虫的死亡率达65.00%(t=16.57,df=8,P<0.0001)(图2)。

图2 取食5 ng/μL dsHvFTZ-F1处理的叶片对茄二十八星瓢虫1龄幼虫存活率的影响Fig.2 The impact of ingetion of 5 ng/μLdsHvFTZ-F1 treated leaves on the survival rate of the 1st instar larvae of H.vigintioctopunctata
2.2 dsHvFTZ-F1处理对茄二十八星瓢虫1龄幼虫20E和色素沉积相关基因表达的影响
经dsHvFTZ-F1处理2 d后,1龄幼虫的HvFTZ-F1基因表达水平显著降低了63.69%(t=4.426,df=4,P=0.011);HvE75(t=11.173,df=2,P=0.008)的表达水平降低了 20.76%;HvHR3(t=3.636,df=2,P=0.068)、HvECR(t=0.14,df=2,P=0.901)的表达量无显著变化(图3A)。
同样,经dsHvFTZ-F1处理后HvDIB、HvSPOOK、HvSHADOW、HvSHADE表达水平显著降低,分别下调 85.36%(t=118.238,df=2,P<0.0001),66.32%(t=49.844,df=2,P<0.0001),52.74%(t=20.336,df=2,P=0.002),31.12%(t=10.993,df=2,P=0.008)(图 3B)。
HvDDC、HvTH、Hvebony表达水平在5 ng/μL dsHvFTZ-F1处理后显著降低,分别降低了88.74%(t=36.913,df=4,P<0.0001),91.97%(t=55.982,df=2,P<0.0001),86.15%(t=47.937,df=2,P<0.0001)(图3C)。
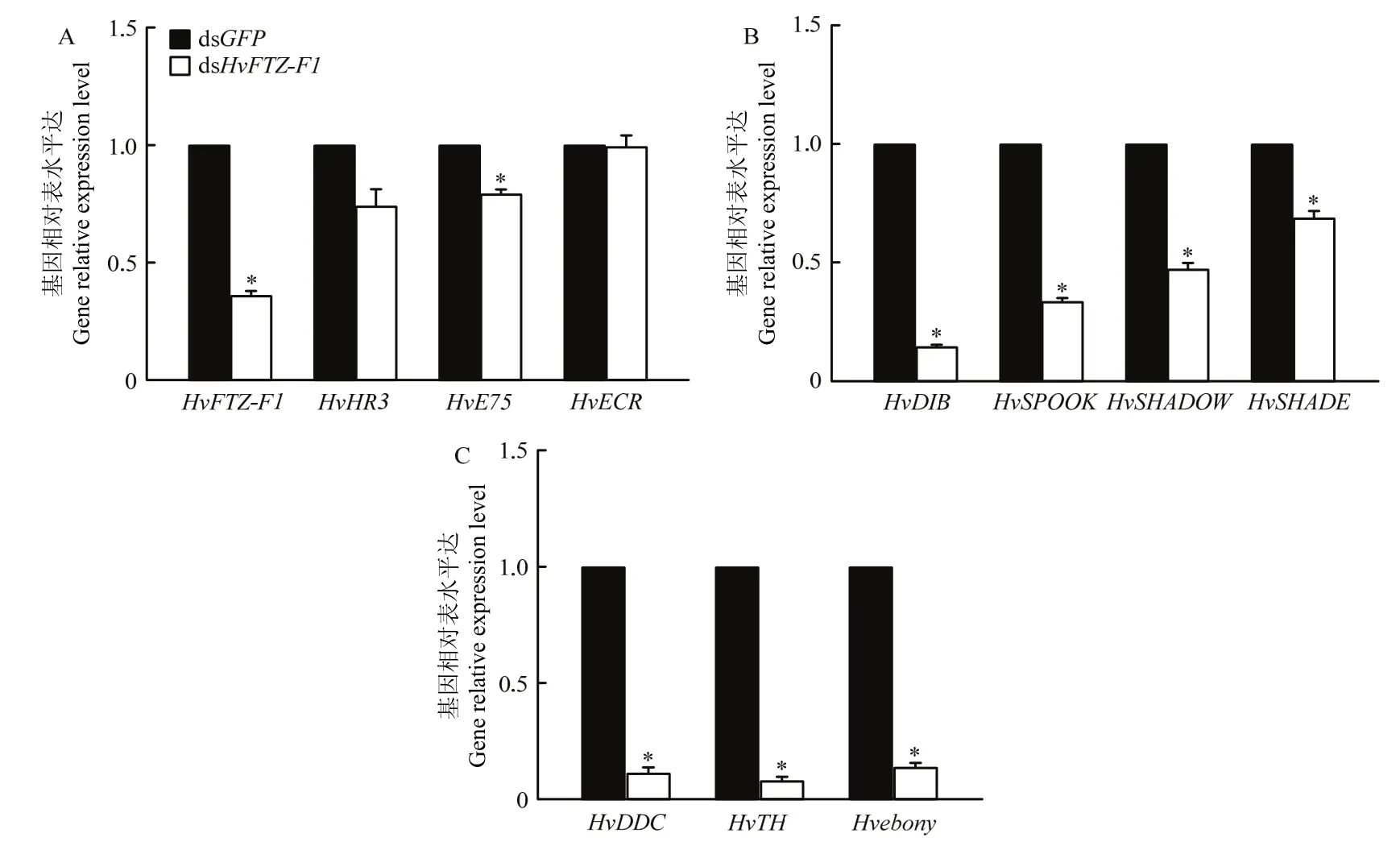
图3 取食5 ng/μL dsHvFTZ-F1处理的叶片对茄二十八星瓢虫1龄幼虫中HvFTZ-F1及相关基因表达水平的影响Fig.3 The impact of ingetion of 5 ng/μL dsHvFTZ-F1 treated leaves on the gene expression of HvFTZ-F1 and related genes in the 1st instar larvae of H.vigintioctopunctata
2.3 dsHvFTZ-F1处理对茄二十八星瓢虫1龄幼虫20E滴度的影响
经dsHvFTZ-F1处理2 d后,1龄幼虫的20E滴度显著降低了15.97%(t=3.693,df=4,P=0.021)(图4)。

图4 取食5 ng/μL dsHvFTZ-F1处理的叶片对茄二十八星瓢虫1龄幼虫中20E滴度的影响Fig.4 The impact of ingetion of 5 ng/μL dsHvFTZ-F1 treated leaves on the 20E titer in the 1st instar larvae of H.vigintioctopunctata
2.4 dsHvFTZ-F1处理对茄二十八星瓢虫1龄幼虫多巴胺滴度的影响
1龄幼虫的多巴胺滴度在dsHvFTZ-F1处理2 d后显著降低了21.38%(t=44.281,df=4,P<0.0001)(图5)。

图5 取食5 ng/μL dsHvFTZ-F1处理的叶片对茄二十八星瓢虫1龄幼虫中多巴胺滴度的影响Fig.5 The impact of ingetion of 5 ng/μL dsHvFTZ-F1 treated leaves on the dopamine titer in the 1st instar larvae of H.vigintioctopunctata
3 讨论
近期研究发现HvFTZ-F1基因在茄二十八星瓢虫1龄幼虫和2龄幼虫中的表达量显著高于3龄和4龄幼虫中的表达量,在幼虫阶段每次蜕皮前或蜕皮时高表达[32]。并且HvFTZ-F1在预蛹期高表达,与FTZ-F1基因在赤拟谷盗[17]、马铃薯叶甲[7]、棉铃虫[21]等昆虫中的研究结果相似。HvFTZ-F1在不同发育阶段的特异表达模式,表明HvFTZ-F1是茄二十八星瓢虫生存发育过程中的关键基因。
为了探究HvFTZ-F1的功能,我们通过饲喂dsHvFTZ-F1抑制茄二十八星瓢虫HvFTZ-F1基因的表达,结果表明HvFTZ-F1基因沉默会导致茄二十八星瓢虫1龄幼虫出现发育障碍并死亡。经处理的1龄幼虫不能正常蜕皮进入2龄,表明HvFTZ-F1在茄二十八星瓢虫蜕皮中发挥重要作用。在黑腹果蝇、淡色库蚊Culex pipiens pallens等昆虫中,FTZ-F1对于表皮形成均有影响,进一步说明HvFTZ-F1可能参与茄二十八星瓢虫表皮的形成[35,36]。处理组幼虫的死亡状态均为后肠携带新鲜粪便从肛门伸出,之前的研究表明,100 ng/μL dsHvFTZ-F1也会导致1龄幼虫出现同样的死亡状态,同时伴随渗透压基因的显著升高,因此我们推测,低剂量的dsHvFTZ-F1处理同样能影响茄二十八星瓢虫1龄幼虫的渗透压调节[32]。
HvFTZ-F1沉默导致1龄幼虫Halloween基因(HvDIB、HvSPOOK、HvSHADOW、HvSHADE)和20E相关基因HvE75表达水平不同程度的下调,显著降低了幼虫体内的20E滴度。在果蝇、马铃薯甲虫和棉铃虫等昆虫中,RNAi介导的FTZ-F1沉默导致蜕皮激素滴度显著降低[7,8,21],我们的研究结果与在这些昆虫中的研究结果一致,表明HvFTZ-F1参与茄二十八星瓢虫的20E信号通路。并且,HvFTZ-F1沉默降低了色素沉着相关基因(HvTH,HvDDC,Hvebony)的表达水平。有研究报道,在亚洲玉米螟Ostrinia furnacalis和西方蜜蜂Apis mellifera中,20E相关基因影响了色素沉积相关基因的表达。向亚洲玉米螟注射20E会导致Oflaccase2表达水平升高[37];西方蜜蜂Amlaccase2的表达水平受蜕皮激素的调节,且Amlaccase2在成虫外骨骼分化过程中起重要作用,其表达的改变可导致蜕皮激素释放受阻和外骨骼异常[38]。类似的其他报道称,向棉铃虫注射20E促进了TH的转录和表达,进一步加强了色素相关基因与20E的相关性[39]。20E可能诱导转录因子参与黑色素生化模块的基因调控网络,促进昆虫蜕皮后的黑化和硬化。
另外,HvFTZ-F1沉默降低了茄二十八星瓢虫1龄幼虫多巴胺滴度,因此我们推测FTZ-F1可通过调控20E信号,控制色素合成基因的表达和多巴胺滴度,并影响昆虫蜕皮,或HvFTZ-F1直接参与茄二十八星瓢虫多巴胺的合成,但需要进一步的实验去证实。在烟草天蛾Manduca sexta中,注射20E会抑制表皮黑色素沉积[11],说明蜕皮激素滴度的降低会启动黑色素合成信号通路,还需进一步探究茄二十八星瓢虫体内的20E滴度下降对多巴胺滴度的变化的影响。
本研究首次采用低浓度 5 ng/μL的 dsHvFTZ-F1处理茄二十八星瓢虫 1龄幼虫,以探究低浓度dsHvFTZ-F1对茄二十八星瓢虫1龄幼虫存活和发育的影响。相较于之前采用100 ng/μL dsHvFTZ-F1的实验结果[32],较低浓度的dsRNA处理同样也能够造成生长发育障碍及较高的死亡率。HvFTZ-F1基因可被认为是一个潜在的靶标基因,用于开发基于RNAi技术的生物农药防治茄二十八星瓢虫。
猜你喜欢
读者(2022年9期)2022-04-22
昆明医科大学学报(2022年2期)2022-03-29
老年医学研究(2021年5期)2022-01-19
华侨大学学报(自然科学版)(2021年4期)2021-07-30
红蜻蜓·低年级(2020年9期)2020-09-26
疯狂英语·新读写(2020年3期)2020-06-06
时代英语·高一(2019年5期)2019-09-03
小学生作文(低年级适用)(2018年4期)2018-11-29
小朋友·快乐手工(2015年10期)2015-11-02
小朋友·快乐手工(2015年2期)2015-03-13
