
不同养殖参数对克氏原螯虾生长及生理生化的影响
2023-06-30 14:27覃宝利王信海王宣朋姜爱兰叶建勇吴春单金峰
江苏农业科学 2023年5期
覃宝利 王信海 王宣朋 姜爱兰 叶建勇 吴春 单金峰

摘要:为获知不同养殖参数放养密度、水草覆盖率及饲料蛋白水平对克氏原螯虾生长及生理生化的影响,采用 L9(34) 正交设计探讨其在不同放养密度(8、12、16尾/m2)、水草覆盖率(20%、40%和60%)及饲料蛋白水平(28%、30%和32%)组合条件下的生长、血清生化指标含量及非特异性免疫酶活性的變化。结果表明,克氏原螯虾生长性能与放养密度呈负相关。放养密度为16尾/m2时,虾末体质量和增长率显著低于低密度处理组(8、12尾/m2)(P<0.05)。成活率在最大密度组显著降低(P<0.05),2个低密度组无显著差异(P>0.05)。水草覆盖率为40%时克氏原螯虾体质量增长率、特定生长率和成活率显著高于20%和60%组(P<0.05)。40%水草覆盖率处理下克氏原螯虾血清白蛋白(ALB)含量均在16尾/m2密度组显著下降,其他血清生化指标含量均无显著差异。20%、60%水草覆盖率处理下白蛋白(ALB)含量变化趋势与40%水草覆盖率处理相同,但甘油三酯(TG)含量和谷丙转氨酶(ALT)活性均在 16尾/m2 密度组显著上升(P<0.05)。同一水草覆盖率处理下,最大密度组虾体血清超氧化物歧化酶(SOD)、过氧化氢酶(CAT)和碱性磷酸酶(AKP)活性最高,显著高于2个低密度组(P<0.05),溶菌酶(LZM)和酸性磷酸酶(ACP)活性最低,显著低于2个低密度组(P>0.05)。饲料蛋白水平对克氏原螯虾生长性能、成活率、血清生化指标含量和非特异性免疫酶活性均无明显相关性。研究提示,放养密度过高可显著抑制克氏原螯虾生长、成活率及生理生化指标,适宜的水草覆盖率可显著提高克氏原螯虾生长性能和成活率。综合评价筛选出克氏原螯虾围网最佳组合养殖方案为水草覆盖率为40%,放养密度为8尾/m2或12尾/m2,饲料蛋白水平为28%。
关键词:克氏原螯虾;放养密度;水草覆盖率;饲料蛋白水平;生长性能;血清生化;非特异性免疫酶
中图分类号:S966.12 文献标志码:A
文章编号:1002-1302(2023)05-0183-08
克氏原螯虾(Procambarus clarkii),俗称淡水小龙虾,属节肢动物门甲壳纲十足目螯虾亚目螯虾科原螯虾属[1]。因其风味鲜美、营养价值高、商业效益高而在我国内陆被广泛养殖。2019年全国克氏原螯虾养殖产量达 208.96 万t,养殖总面积达 128.6 万hm2[2]。但随着养殖规模逐年扩大,养殖问题日益凸显,如各种病害疫情频发、规格提升困难及生长速度慢等,使龙虾养殖业效益大大降低,严重制约其养殖业健康稳定持续发展。这与市场有关,更与养殖技术有关,养殖技术涉及的问题很多[3]。在实际生产中,养殖者为追求更大产量,往往进行高密度养殖。而甲壳类动物的高密度养殖通常会加剧资源竞争,如生存空间、隐蔽物、食物[4]、同类相残[5];拥挤胁迫,如食欲低下、生理状态差、抗病能力弱[6],导致成活率、规格、生长速度下降。另一方面,克氏原螯虾生性喜斗,有同类相残习性,是典型具有社会行为的动物。克氏原螯虾在蜕壳时,为避免被捕食,需要找隐蔽物隐藏。有研究表明,隐蔽物不仅可显著减少甲壳类动物的同类相残,还能改善甲壳类动物的生存和生长[7-9]。伊乐藻作为一种大型耐寒沉水植物,不仅为克氏原螯虾提供栖息和隐蔽的场所,还为克氏原螯虾提供营养丰富的饵料,净化水质,防暑降温、防风避寒、提升免疫力,已被广泛用于克氏原螯虾养殖中。一般来说,甲壳类动物隐蔽物的有效性随其密度和复杂性的增加而增强[7],但实际生产中往往会出现伊乐藻面积过高,大量腐烂、恶化水质等问题,需合理控制栽种面积。再者,克氏原螯虾在本质上属于肉食性动物,饲料中适宜的蛋白质水平对维持其良好的生长性能和健康至关重要。饲料蛋白水平不足或过量不仅会对养殖对象的生长、蜕壳和免疫产生负面影响,还会增加养殖水体的氮排放,提高养殖成本[10]。目前,关于克氏原螯虾对蛋白质适宜需求量的研究结果差异较大。因此,有必要对克氏原鳌虾养殖的适宜密度、水草覆盖率和饲料蛋白水平进行精准评估。
本研究以克氏原螯虾亚成体虾作为试验对象,通过45 d池塘围网养殖生长试验分析克氏原螯虾的生长性能、存活率、血清生化指标及非特异性免疫酶活性的变化,明确克氏原鳌虾养殖的适宜密度、水草覆盖率和饲料蛋白水平组合参数,旨在为克氏原螯虾健康养殖提供数据支撑。
1 材料与方法
1.1 试验材料
试验用幼虾采自江苏省泗阳县庆江水产养殖专业合作社克氏原螯虾养殖基地,选择身体健壮,无疾病克氏原螯虾,规格为250~300尾/kg,所植水草为伊乐藻。
1.2 试验设计
2020年4月2日,在江苏省宿迁市宿豫区来龙镇陵园村玲龙水产养殖专业合作社克氏原螯虾养殖基地6 hm2小龙虾精养塘口内,架设9个围网(边长6 m×6 m,高度1.2 m),设定水草覆盖率(20%、40%、60%)、投放密度(8、12、16尾/m2)和饲料蛋白水平(28%、30%、32%)3个因素进行正交试验,组成9个组合,在池塘内按这9个组合架设围网(面积为36 m2/个)。在围网内按照L9(34)正交试验法对水草覆盖率(因素A)、放养密度(因素B)、饲料蛋白水平(因素C)进行组配,由表1可知,各试验组克氏原螯虾的放养量、水草覆盖比例和饲料蛋白水平,按组配进行水草种植、虾苗投放和不同蛋白饲料投喂。由表2可知,试验分9个处理组,每组设3个重复。试验期间每日08:00和17:00投喂人工配合饲料,日投饵率为3%~5%,试验时间为45 d。试验结束后取样测定克氏原螯虾的生长及生理生化指标。
1.3 采样与指标测定
1.3.1 生长性能测定
饲养试验结束后,试验虾停饲24 h,然后将各围网中克氏原螯虾计数并称取其终末体质量,计算体质量增长率、特定生长率、成活率,公式如下:
成活率=Nt/N0×100%;
体质量增长率=(Wt-W0)/W0×100%;
特定生长率=(lnWt-lnW0)/t×100%。
式中:Nt为试验虾终末尾数,N0为初始尾数,尾;Wt为终末体质量,W0为初始体质量,g;t为试验天数,d。
1.3.2 血液生化指标的测定
采用無菌注射器取虾体围心腔的血液,将血液放入无菌离心管4 ℃冰箱,过夜后离心,取血清,用于总蛋白(TP)、白蛋白(ALB)、总胆固醇(TCH)、甘油三酯(TG)的含量及谷丙转氨酶(ALT)、超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、碱性磷酸酶(AKP)、酸性磷酸酶(ACP)和溶菌酶(LZM)活性的测定,方法参照南京建成生物工程研究所试剂盒说明书。
1.4 数据处理
试验结果数据以“平均值±标准差”表示,采用SPSS 19.0软件进行单因素方差分析,双重比较或Duncans多重比较,P<0.05表明差异显著。
2 结果与分析
2.1 水草覆盖率、放养密度和饲料蛋白水平对克氏原螯虾生长性能和成活率的影响
由表2、表3可知,放养密度最大组(16尾/m2)克氏原螯虾成活率显著低于2个低密度组(12、8尾/m2)(P<0.05),2个低密度组间无显著差异(P>0.05)。40%水草覆盖率组克氏原螯虾成活率明显高于20%和60%组(P<0.05),60%组明显高于20%组(P<0.05)。40%水草覆盖率组体质量增长率和特定生长率显著高于20%和60%组(P<0.05),60%组与20%组无显著差异(P<0.05)。16尾/m2组克氏原螯虾体质量增长率和特定生长率显著低于8尾/m2密度组(P<0.05),2个低密度间无明显差异。由表3可知,4#组虾末体质量与5#和7#组无显著差异(P>0.05),但显著高于其他组(P<0.05)。4#组虾体质量增长率与5#组无显著差异(P>0.05),但显著高于其他组(P<0.05)。4#、5#、7#组特定生长率与1#、6#、8#组无显著差异,但显著高于2#、3#和9#组。4#、5#组间成活率差异不明显,但明显高于其他组。
由表4可知,水草覆盖率和放养密度对克氏原螯虾生长性能和成活率均有显著影响(P<0.05),饲料蛋白水平对克氏原螯虾生长性能和成活率无显著影响(P>0.05)。3个因素中,水草覆盖率对克氏原螯虾体质量增长率和特定生长率的影响作用最大,放养密度次之,饲料蛋白水平最小。放养密度对克氏原螯虾成活率的影响作用最大,水草覆盖率次之,饲料蛋白水平最小。
2.2 水草覆盖率、放养密度和饲料蛋白水平对克氏原螯虾血清生化指标和非特异性免疫酶活性的影响
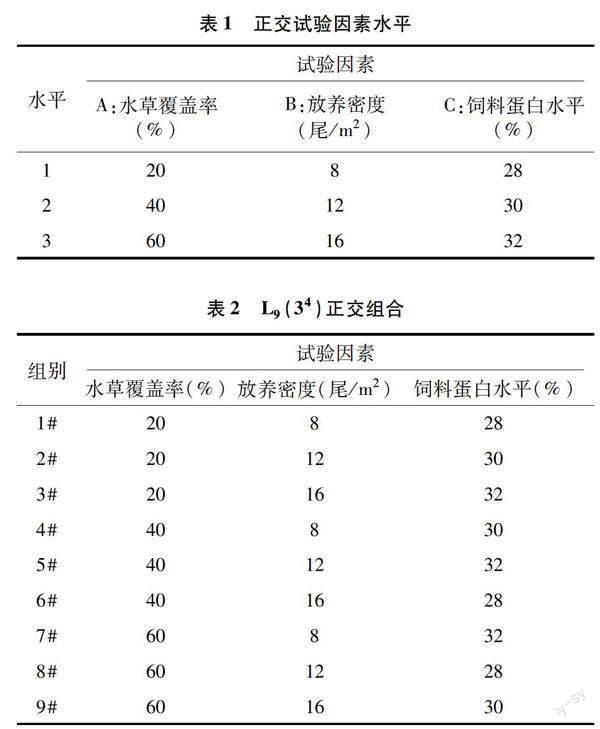
由表5可知,3#、6#、9#组TP含量虽有明显下降,但不同处理组间无显著差异(P>0.05),白蛋白含量显著下降(P<0.05)。5#组总胆固醇含量与 1#、4#、7#组无显著差异(P>0.05),但显著高于2#、3#、6#、8#、9#组(P<0.05),后者之间无显著差异(P>0.05)。3#、9#组之间甘油三酯含量无显著差异(P>0.05),但均显著高于除8#之外的其他组(P<0.05)。3#、9#组谷丙转氨酶活性分别显著高于1#和2#组、7#和8#组(P<0.05),4#、5#、6#组之间谷丙转氨酶活性无显著性差异(P>0.05)。
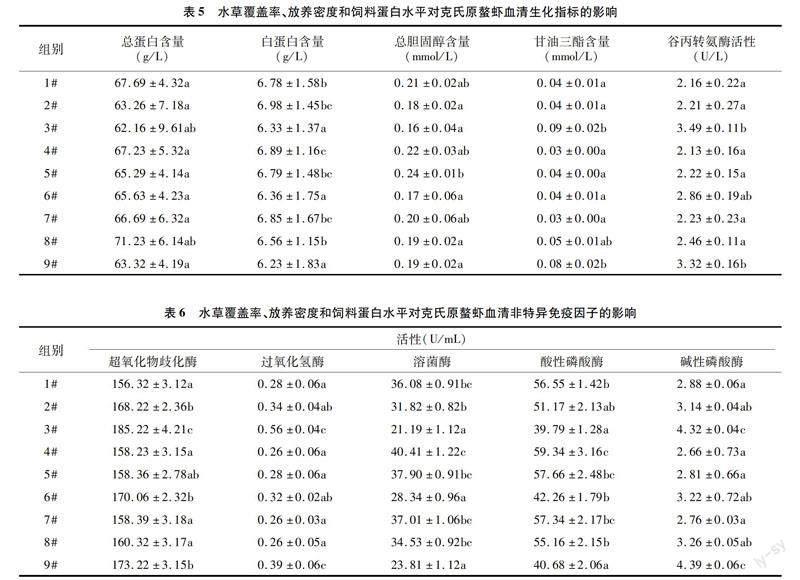
由表2、表6可知,在相同水草覆盖率水平内,随放养密度增大,克氏原螯虾超氧化物歧化酶、碱性磷酸酶和过氧化氢酶活性逐渐升高,溶菌酶和酸性磷酸酶活性逐渐降低。1#、4#、7#、8#组超氧化物歧化酶活性无显著差异(P>0.05),但显著低于2#、3#、6#、9#组,4#与5#无显著差异(P>0.05)。1#、4#、5#、8#组过氧化氢酶活性无显著差异(P>0.05),显著低于3#、9#(P<0.05),6#与4#、5#组无显著差异。1#、4#、5#、7#、8#组溶菌酶活性无显著差异(P>0.05),但显著高于3#、6#、9#组(P<0.05)。4# 组酸性磷酸酶含量与5#、7#组含量无显著差异(P>0.05),但显著高于其他组(P<0.05)。3#、9#组碱性磷酸酶活性无显著差异(P>0.05),但显著高于其他组(P<0.05),其他组间无显著差异(P>0.05)。其中,4#组溶菌酶和酸性磷酸酶活性最高,碱性磷酸酶活性最低。
3 讨论
3.1 放养密度、水草覆盖率和饲料蛋白水平对克氏原螯虾生长性能和成活率的影响
水产养殖中高密度养殖通常会增加水体生产力,但密度过高会降低水产动物存活率和生长性能。肖鸣鹤等发现,随着放养密度的增大,克氏原螯虾幼虾末体质量减小[11]。刘国兴等研究发现,随放养密度增大,克氏原螯虾终末体质量、增重率和特定生长率逐渐降低[12]。Yu等发现,较高的养殖密度对克氏原螯虾生长有负面影响[7]。本研究中,克氏原螯虾生长性能随放养密度的增加而降低,其中虾体末质量和体质量增长率在密度最大组(16尾/m2)显著降低。Romaire等在野外围隔试验中发现,克氏原螯虾放养密度在5、10尾/m2时个体体质量均超过20 g,但在15尾/m2以上时虾体均发生显著的发育不良[13]。Naranjo-Prano等发现,红螯螯虾养殖密度≤11尾/m2时,虾体终末体质量在 25 g,养殖密度增加终末体质量下降[14]。另刘明明也发现,在伊乐藻环境中,当放养密度高于 15尾/m2 时,克氏原螯虾的增重率显著降低[15],本研究结果与之类似。生物生长率不仅由物种本身决定,还由环境因素决定。虾蟹类属于排氨型代谢动物,其蛋白质代谢的最终产物氨是直接排出体外的[16]。因此,在空间区域一定的高密度环境下,幼虾相互接触抵抗增加,能耗增加,用于生长的部分减少,代谢废物增多,生存环境不健康,从而抑制生长[17]。同样,养殖密度对成活率也有负面影响。有研究表明,克氏原螯虾成活率随养殖密度增大而减小[7,18-20]。本研究中克氏原螯虾养殖密度最大组(16尾/m2)成活率明显低于低密度组(8、12尾/m2),2个低密度组间差别较小。廖锐等发现,每种鱼均有一个临界养殖密度,超过这个临界密度时,死亡率上升,而在这个临界密度内,成活率不受影响[21]。因此,本研究中8、12尾/m2 2个密度值可能在该生存条件下的密度临界值内,而16尾/m2的养殖密度可能超过了该规格的临界密度,引起密度胁迫,个体竞争加剧,能耗增多,导致自相残食率增加,死亡率上升。
水草对克氏原螯虾养殖非常重要,因为它不仅是克氏原螯虾喜食的饵料,还是克氏原螯虾蜕壳和躲避的隐蔽场所。而隐蔽场所的有效性通常会随其密度和复杂性的升高而增加[22-23],但隐蔽场所覆盖率过高时,克氏原螯虾获得食物耗费的时间和能量可能会大大增加[24]。因此,在克氏原螯虾养殖过程中,维持适宜的水草覆盖率很有必要。本研究中水草覆盖率为40%时,克氏原螯虾的生长性能和成活率的整体水平明显高于20%和60%组。Yu等从不同水草覆盖率对克氏原螯虾幼虾生存、生长和蜕皮的影响结果,综合判断得出养殖克氏原螯虾适宜的水草覆盖率是50%,本研究结果与之接近[7]。方春林等在研究克氏原螯虾仿生态繁殖池水草布局时,发现水草栽植面积占池塘面积的40%,比20%和80%组更有利于克氏原螯虾亲虾的成熟度、抱卵量、抱卵率和存活率的提高[18]。秦伟等在进行克氏原螯虾养殖试验时,发现水草覆盖率35%试验组池塘底泥微生物种类比25%试验组丰富[25]。通过不同角度提示40%可能是克氏原螯虾养殖水草覆盖率的最佳数值。
大多数研究表明,虾类生长性能随饲料中粗蛋白水平的增加呈先上升后下降的趋势。Miao等研究发现,克氏原螯虾[(5.17±0.32) g]在32%和34%蛋白水平下虾最终体质量、增重率和特定生长率显著高于26%和28%组[26]。夏思瑶等的研究表明,饲料蛋白水平为30.09%时,克氏原螯虾幼虾[(3.69±0.05) g]增重率、特定生长率显著高于其他蛋白水平组[27]。王自蕊等研究发现,克氏原螯虾[(9.45±1.33) g]生长需求适宜的蛋白质质量分数为31.37%~32.20%[28]。但本研究中不同饲料蛋白水平下克氏原螯虾的生长性能无显著差异,可能是不同试验所用虾规格大小、养殖环境及试验设置的蛋白水平梯度范围不同所致。徐维娜等对均体质量为(7.03±0.05) g的克氏原螯虾幼虾饲喂8周,24%、27%和30%的饲料蛋白水平下,克氏原螯虾的生长性能无显著差异[29],本研究结果与之类似。不同饲料蛋白水平下克氏原螯虾的成活率无显著差异,与Miao等的研究结果[26-29]一致。苏时萍等指出,克氏原螯虾能消化的饲料蛋白水平最高为 35%~40%[30],说明本试验设置的蛋白水平梯度在克氏原螯虾适宜需求范围内,28%的蛋白水平即可满足克氏原螯虾的生长需求。
本研究通过不同处理因素对克氏原螯虾生长性能和成活率影响的显著性和3因素的主次分析发现,水草覆盖率和放养密度对克氏原螯虾生长性能和成活率均有显著影响,其中放养密度对克氏原螯虾的成活率影响最大,水草覆盖率对克氏原螯蝦的生长性能指标影响最大。Shao等研究水温、饲养密度和鱼粉豆粕比例对克氏原螯虾早熟影响时,发现饲养密度始终是影响成活率、体质量增加、性腺指数和成熟率的最重要因素[31]。提示本研究的3个因素中,放养密度是决定虾生存的最重要因素,水草覆盖率是决定虾生长的最重要因素,进而说明放养密度和水草覆盖率在小龙虾的水产养殖中具有重要作用。
3.2 水草覆盖率、放养密度和饲料蛋白水平对克氏原螯虾血清生化指标的影响
血液生化指标能够指示动物的生理和代谢水平,与动物的营养变化、健康状况和环境状况密切相关[32]。胆固醇甘油三脂主要在肝脏合成,是血脂的主要成分[33]。本研究不同密度处理组克氏原螯虾血清总胆固醇含量差异不显著。胆固醇作为一种非极性脂质成分,属于类固醇化合物,广泛存在于动物体内,是动物组织和细胞中许多具有生理活性化合物的化学框架[34]。它是甲壳类动物必需的营养物质,被用来维持生理功能,并进一步参与新陈代谢,或通过肠道排出,不太可能被吸收作为能量来利用[35]。这可能是本试验中虾血清总胆固醇受密度变化影响较小的原因。血脂水平的高低与机体的代谢、生理状态密切相关,被广泛用于评估鱼类对环境的适应能力和健康状况[36]。摄食脂类食物过多或肥胖可引起血清甘油三酯含量升高,而运动会降低其含量。本研究中,20%水草覆盖率和60%水草覆盖率水平内放养密度最大组甘油三酯含量显著高于低密度组,40%水草覆盖率水平内不同密度组含量差异不显著。这可能是20%水草覆盖率和60%水草覆盖率水平内放养密度最大组虾运动低于其他组的原因。该结果反映40%水草覆盖率的环境可能更适合克氏原螯虾生长。肖鸣鹤等发现,克氏原螯虾幼虾肝胰腺中甘油三酯随着养殖密度的增大呈不断增多的趋势[13]。张海恩等发现,中国明对虾血清甘油三酯含量在高密度组比低密度组显著下降[37]。倪蒙等研究得出,不同放养密度对罗氏沼虾血清甘油三酯含量变化影响不显著[38]。以上研究与本研究结果不同,可能是不同养殖条件、不同物种血脂代谢对养殖密度变化的响应不同。谷丙转氨酶是肝脏受损的重要指示酶,通常存在于肝细胞中,若肝细胞受损或死亡,它便会进入到血液中,造成血清中该酶活性增加。本研究中虾体血清谷丙转氨酶活性均在放养密度最大时显著升高,说明高密度组克氏原螯虾肝脏细胞受损。血清总蛋白分为白蛋白和球蛋白,其浓度是鱼体健康状况的基本指标[39],其中白蛋白含量越高标志着生长速度越快,球蛋白含量越高,标志着免疫能力越强[40]。尽管结果显示各处理组血清总蛋白含量差异不显著,但是密度最大组虾体血清总蛋白含量明显下降,白蛋白的含量均在密度最大组显著降低。说明放养密度在16尾/m2时克氏原螯虾处于亚健康状态,生长受到明显抑制,这与虾体末质量、体质量增长率、特定生长率和成活率的结果表现一致。
本研究中饲料蛋白水平与血清总蛋白、白蛋白、甘油三酯、总胆固醇、谷丙转氨酶含量无明显相关性。杨弘等对尼罗罗非鱼幼鱼的饲料蛋白水平(25%~45%)研究中也发现类似的结果[41]。说明本试验条件下饲料蛋白水平为28%即可满足克氏原螯虾的生长需求。姜松等研究发现,斑节对虾血液中的谷丙转氨酶和总蛋白、白蛋白的含量表现出随着蛋白水平的增加而上升的趋势,且在各饲料组中均有显著性差异[42]。Lu等发现,较高饲料蛋白水平(30.56%~44.64%)可诱导克氏原螯虾[(12.86±0.10) g]血液中总蛋白、白蛋白、总胆固醇含量和丙氨酸转氨酶活性显著增加[43],本研究结果与之不同。这可能是试验虾的种类、规格大小、饲料蛋白源、蛋白水平梯度及养殖环境不同,导致虾体血液生理生化响应差异较大。
本研究中,水草覆盖率水平对克氏原螯虾血清总蛋白、白蛋白、总胆固醇含量及谷丙转氨酶活性均无显著影响。说明水草覆盖率并非是该条件下影响克氏原螯虾血清生化指标的关键因素,更大可能是发挥了环境生态功能。
3.3 水草覆盖率、放养密度和饲料蛋白水平对克氏原螯虾血清非特异性免疫酶活性的影响
甲壳类动物以非特异性免疫为主[20]。本研究中水草覆盖率和饲料蛋白水平对虾体非特异免疫酶活性变化无明显影响,放养密度有显著影响。其中,高密度组SOD、CAT、AKP活性随放养密度的增加有显著上升趋势,ACP、LZM活性随放养密度的增加有显著下降趋势。说明密度条件对虾体血清非特异性免疫酶活性变化的影响占主导因素,且不同类型的非特异性免疫酶对密度变化引起环境变化的响应不完全相同。密度胁迫导致机体内产生大量活性氧自由基,而SOD、CAT活性升高可清除这些自由基,保护机体免受氧化损伤。本研究中,高密度组SOD、CAT活性显著高于低密度组,2个低密度组之间无显著差异。可能是高密度组个体受到密度胁迫时,新陈代谢加快,能量消耗升高,一定程度上促进了氧自由基的产生,进而导致SOD和CAT含量升高。在机体免疫过程中,LZM可催化细菌细胞壁水解,导致细菌溶解死亡,也可诱导调节其他免疫因子产生[44]。本研究中高密度组克氏原螯虾血清溶菌酶活性显著低于2个低密度组。其他研究也发现,克氏原螯虾肝胰腺溶菌酶活性随养殖密度的升高呈下降趋势[20,45]。俄罗斯鲟幼鱼也出现高密度组溶菌酶水平显著低于中、低密度组的现象[46],说明密度过高导致克氏原螯虾抗菌能力下降,进而机体免疫力下降。ACP是细胞在发挥吞噬和包裹作用过程中所释放的物质。本研究结果显示,高密度组血清 ACP和LZM活性变化相同,可能是因为吞噬细胞杀菌的物质基础LZM活性显著降低,吞噬细胞的吞噬功能显著减弱,进而伴随产生ACP显著减少。血清AKP主要源于肝脏[47],是构成甲壳动物解毒系统的组成之一,在非特异性免疫中发挥重要作用[48]。研究结果显示,AKP活性随密度增加呈上升趋势,16尾/m2组显著高于12、8尾/m2组。范文浩等发现,克氏原螯虾肝脏碱性磷酸酶活性随密度增加有上升趋势[45],本研究结果与之类似。吴垠等研究发现,患病中国明对虾血清碱性磷酸酶活性明显高于正常虾,且发病初期该酶活性远低于重症期[49]。在电镜酶细胞化学技术下,正常中国明对虾血清几乎不显示AKP活性,但病理状态下反而升高[50]。因此,推测本研究中AKP活性越高说明克氏原螯虾体质可能越差。吴宗凡等对鲫鱼[51]、陈亚坤等对凡纳滨对虾[52]及王博等對墨吉明对虾[53]的研究中发现密度对血清AKP活性有负面影响。这可能是因为不同水产动物碱性磷酸酶理化性质和功能基团的差异性导致其在不同条件下酶活力不同[54]。
4 结论
本研究中放养密度对克氏原螯虾成活率影响作用最大,水草覆盖率次之,饲料蛋白水平最小;水草覆盖率对克氏原螯虾体质量增长率和特定生长率的影响作用最大,放养密度次之,饲料蛋白水平最小。
综合分析放养密度、水草覆盖率、饲料蛋白水平3个因子对克氏原螯虾存活、生长及血清生化指标和非特异性免疫酶活性的影响,得出最佳组合方案为水草覆盖率为40%,放养密度为8尾/m2或 12尾/m2,饲料蛋白水平为28%。
参考文献:
[1]Gherardi F. Crayfish invading Europe:the case study of Procambarus clarkii[J]. Marine and Freshwater Behaviour and Physiology,2006,39(3):175-191.
[2]农业农村部渔业渔政管理局,全国水产技术推广总站,中国水产协会. 2020中国小龙虾产业发展报告[J]. 中国水产,2020,7:8-17.
[3]舒新亚. 浅析小龙虾养殖业的几个重要问题(五)[J]. 渔业致富指南,2021(16):42-45.
[4]McClain W R. Effects of population density and feeding rate on growth and feed consumption of red swamp crawfish Procambarns clarkii[J]. Journal of the World Aquaculture Society,1995,26(1):14-23.
[5]Romano N,Zeng C S. Cannibalism of decapod crustaceans and implications for their aquaculture:a review of its prevalence,influencing factors,and mitigating methods[J]. Reviews in Fisheries Science & Aquaculture,2017,25(1):42-69.
[6]Nga B T,Lürling M,Peeters E T H M,et al. Chemical and physical effects of crowding on growth and survival of Penaeus monodon Fabricius post-larvae[J]. Aquaculture,2005,246(1/2/3/4):455-465.
[7]Yu J X,Xiong M T,Ye S W,et al. Effects of stocking density and artificial macrophyte shelter on survival,growth and molting of juvenile red swamp crayfish (Procambarus clarkii) under experimental conditions[J]. Aquaculture,2020,521:735001.
[8]Olsson K,Nystrm P. Non-interactive effects of habitat complexity and adult crayfish on survival and growth of juvenile crayfish (Pacifastacus leniusculus)[J]. Freshwater Biology,2009,54(1):35-46.
[9]Mirera O,Moksnes P O. Cannibalistic interactions of juvenile mud crabs Scylla serrata:the effect of shelter and crab size[J]. African Journal of Marine Science,2013,35(4):545-553.
[10]Naylor M A,Kaiser H,Jones C L W. The effect of dietary protein level on total ammonia nitrogen and free ammonia nitrogen concentrations in a serial-use raceway used to farm South African abalone,Haliotis midae Linnaeus,1758[J]. Journal of Shellfish Research,2011,30(2):337-341.
[11]肖鳴鹤,肖英平,吴志强,等. 养殖密度对克氏原螯虾幼虾生长、消化酶活力和生理生化指标的影响[J]. 水产学报,2012,36(7):1088-1093.
[12]刘国兴,李 玲,彭 刚,等. 放养密度对克氏原螯虾生长和养殖水质的影响[J]. 江西农业学报,2014,26(4):86-89,93.
[13]Romaire R P,Villágran E R. Evaluation of stocking density and feeding regime on production of red swamp crawfish in outdoor mesocosms[J]. Journal of the World Aquaculture Society,2010,41(3):298-307.
[14]Naranjo-Páramo J,Hernandez-Llamas A,Villarreal H. Effect of stocking density on growth,survival and yield of juvenile redclaw crayfish Cherax quadricarinatus (Decapoda:Parastacidae) in gravel-lined commercial nursery ponds[J]. Aquaculture,2004,242(1/2/3/4):197-206.
[15]刘明明. 4种水草用于克氏原螯虾生态养殖的初步研究[D]. 上海:上海海洋大学,2020.
[16]王克行. 虾蟹类增养殖学[M]. 北京:中国农业出版社,1997:13-14.
[17]刘广斌,马 甡. 凡纳滨对虾高密度养殖实验[J]. 海洋湖沼通报,2004(3):54-58.
[18]方春林,王庆萍,余智杰,等. 克氏原螯虾仿生态繁殖池布草技术研究[J]. 水产科技情报,2012,39(1):30-33.
[19]邓梦颖,吴志强,肖英平,等. 养殖密度对克氏原螯虾幼虾生长、摄食和饵料利用影响[J]. 淡水渔业,2010,40(3):13-17.
[20]陈 勇. 饲养密度对克氏原螯虾成活率和肝胰腺三种免疫酶的影响[J]. 湖北农业科学,2016,55(16):4237-4240.
[21]廖 锐,区又君,勾效伟. 养殖密度对鱼类福利影响的研究进展Ⅰ.死亡率、生长、摄食以及应激反应[J]. 南方水产,2006,2(6):76-80.
[22]Sáez-Royuela M,Carral J,Celada J,et al. Effects of shelter type and food supply frequency on survival and growth of stage-2 juvenile white-clawed crayfish (Austropotamobius pallipes Lereboullet) under laboratory conditions[J]. Aquaculture International,2001,9(6):489-497.
[23]Amaral V,Paula J,Hawkins S,et al. Cannibalistic interactions in two co-occurring decapod species:effects of density,food,alternative prey and habitat[J]. Journal of Experimental Marine Biology and Ecology,2009,368(1):88-93.
[24]程慧俊. 克氏原螯虾稻田养殖生态学的初步研究[D]. 武汉:湖北大学,2014.
[25]秦 伟,周 鑫,徐增洪,等. 不同放养密度和水草覆盖率下克氏原螯虾池塘底泥中微生物群落特征[J]. 水产科学,2015,34(10):621-628.
[26]Miao S Y,Han B,Li J Q,et al. Effects of dietary protein level on the growth performance,feed utilization and immunity of red swamp crayfish Procambarus clarkia[J]. Aquaculture Reports,2020,18:100540.
[27]夏思瑶,万金娟,孟祥龙,等. 饲料中蛋白质水平对克氏原螯虾生长性能及肌肉品质的影响[J]. 水产养殖,2021,42(3):6-12.
[28]王自蕊,李永安,许志城,等. 饲料蛋白质水平对克氏原螯虾生长性能的影响[J]. 水产科技情报,2020,47(4):202-205.
[29]徐维娜,刘文斌,沈美芳,等. 饲料中不同蛋白质和脂肪水平对克氏螯虾(Procambarus clarkii)生长性能、体组成和消化酶活性的影响[J]. 海洋与湖沼,2011,42(4):521-529.
[30]苏时萍,施培松,杨启超,等. 饲料蛋白质水平对克氏原螯虾幼体消化酶活性和肌肉成分的影响[J]. 安徽农业大学学报,2009,36(2):231-235.
[31]Shao G M,Tan H Y,Wang Y F. Effect of combined water temperature,stocking density and fish meal:soybean meal ratio in diet on precocity of the crayfish Procambarus clarkii (Girard)[J]. Aquaculture Research,2018,49(5):2081-2083.
[32]Lermen C L,Lappe R,Crestani M,et al. Effect of different temperature regimes on metabolic and blood parameters of silver catfish Rhamdia quelen[J]. Aquaculture,2004,239(1/2/3/4):497-507.
[33]徐 濱,魏开金,马宝珊,等. 不同克氏原螯虾养殖模式下幼虾生长、体成分和血淋巴生化指标的比较[J]. 淡水渔业,2021,51(5):84-90.
[34]Shyn-Shin S,Ping-Chung L,Shiu-Nan C,et al. Cholesterol requirement of juvenile tiger shrimp (Penaeus monodon)[J]. Aquaculture,1994,125(1/2):131-137.
[35]Tian H Y,Yang C,Yu Y B,et al. Dietary cholesterol level affects growth,molting performance and ecdysteroid signal transduction in Procambarus clarkii[J]. Aquaculture,2020,523:735198.
[36]刘 群,温海深,李吉方,等. 网箱养殖密度对虹鳟甲状腺激素及血脂指标的影响[J]. 水生生物学报,2014,38(6):1076-1083.
[37]张海恩,何玉英,李 健,等. 不同养殖密度对中国明对虾生长和能量代谢的影响[J]. 渔业科学进展,2021,42(5):70-76.
[38]倪 蒙,陈雪峰,高 强,等. 放养密度对温棚养殖罗氏沼虾生长、生化指标、水质及养殖效益的影响[J]. 大连海洋大学学报,2021,36(3):423-429.
[39]Misra S,Sahu N P,Pal A K,et al. Pre-and post-challenge immuno-haematological changes in Labeo rohita juveniles fed gelatinised or non-gelatinised carbohydrate with n-3 PUFA[J]. Fish & Shellfish Immunology,2006,21(4):346-356.
[40]Li X F,Liu W B,Lu K L,et al. Dietary carbohydrate/lipid ratios affect stress,oxidative status and non-specific immune responses of fingerling blunt snout bream,Megalobrama amblycephala[J]. Fish & Shellfish Immunology,2012,33(2):316-323.
[41]杨 弘,徐起群,乐贻荣,等. 饲料蛋白质水平对尼罗罗非鱼幼鱼生长性能、体组成、血液学指标和肝脏非特异性免疫指标的影响[J]. 动物营养学报,2012,24(12):2384-2392.
[42]姜 松,杨其彬,黄建华,等. 饲料蛋白水平对斑节对虾生长、血液生化指标及体成分的影响[J]. 海洋渔业,2014,36(1):44-50.
[43]Lu X,Peng D,Chen X R,et al. Effects of dietary protein levels on growth,muscle composition,digestive enzymes activities,hemolymph biochemical indices and ovary development of pre-adult red swamp crayfish (Procambarus clarkii)[J]. Aquaculture Reports,2020,18:100542.
[44]刘永涛,董 靖,夏京津,等. 不同饲料对稻田养殖克氏原螯虾生长、非特异性免疫酶及体成分的影响[J]. 中国渔业质量与标准,2020,10(1):43-51.
[45]范文浩,方 刘,周 锦,等. 养殖密度对克氏原螯虾生长及消化酶、免疫酶活性的影响[J]. 水产科学,2021,40(2):261-266.
[46]曹 阳,李二超,陈立侨,等. 养殖密度对俄罗斯鲟幼鱼的生长、生理和免疫指标的影响[J]. 水生生物学报,2014,38(5):968-974.
[47]王冰心,叶均安. 虾类血清中免疫相关酶的研究进展[J]. 中国饲料,2009(3):27-28,36.
[48]邹广众,孙虎山. 水产甲壳动物免疫学研究进展与前景展望[J]. 生命科学仪器,2009,7(6):17-21.
[49]吴 垠,邢殿楼,祝国芹,等. 中国对虾暴发性流行病的血液病理研究[J]. 中国水产科学,1998,5(3):53-57.
[50]刘晓云,张志峰,马洪明. 中国对虾血细胞酶细胞化学的初步研究[J]. 青岛海洋大学学报(自然科学版),2002,32(2):259-265.
[51]吴宗凡,时 旭,程果锋,等. 养殖密度对温室湿地循环水系统中鲫生长、生理及免疫指标的影响[J]. 南方水产科学,2014,10(5):39-44.
[52]陈亚坤,郭 冉,夏 辉,等. 密度胁迫对凡纳滨对虾生长、水质因子及免疫力的影响[J]. 江苏农业科学,2011,39(3):292-294.
[53]王 博,秦海鵬,廖栩峥,等. 养殖密度对生物絮团养殖系统中墨吉明对虾免疫、生长和水质的影响[J]. 渔业现代化,2019,46(6):61-67.
[54]张 辉,张海莲. 碱性磷酸酶在水产动物中的作用[J]. 河北渔业,2003(5):12-13,32.
收稿日期:2022-04-02
基金项目:江苏省农业科技自主创新资金[编号:CX(19)3018];江苏省宿迁市自主创新项目(编号:SQCX202108)。
作者简介:覃宝利(1986—),女,河南鹤壁人,硕士,助理研究员,主要从事水产养殖方面的研究。E-mail:906963793@qq.com。
通信作者:王信海,硕士,副研究员,主要从事稻田综合种养方面的研究。E-mail:20141602@jaas.ac.cn。
猜你喜欢
安徽农业科学(2020年14期)2020-08-04
农村农业农民·B版(2017年6期)2017-06-23
湖北农业科学(2016年18期)2016-12-08
河北渔业(2016年10期)2016-11-22
安徽农学通报(2016年8期)2016-05-17
江苏农业科学(2015年11期)2016-01-27
江苏农业科学(2015年8期)2015-09-10
家禽科学(2015年5期)2015-05-30
河北渔业(2015年2期)2015-03-26
当代畜禽养殖业(2013年4期)2013-08-15
