
基于Cyt b序列分析黄沙鳖和日本鳖及其杂交子一代的遗传多样性
2023-09-08 09:50肖何威胡亚洲秦溱徐大建梁艺馨刘小燕王晓清
湖南农业大学学报(自然科学版) 2023年4期
肖何威,胡亚洲,秦溱,徐大建,梁艺馨,刘小燕,王晓清
基于序列分析黄沙鳖和日本鳖及其杂交子一代的遗传多样性
肖何威,胡亚洲,秦溱,徐大建,梁艺馨,刘小燕,王晓清*
(湖南农业大学动物科学技术学院,湖南 长沙 410128)
为了解中华鳖()中黄沙鳖和日本鳖及其杂交子一代的遗传特性,分别对它们的线粒体基因测序,并进行遗传多样性分析。结果表明:PCR扩增测序黄沙鳖和日本鳖及其杂交子一代3个群体共获得90条序列,18种单倍型;90个个体的基因碱基A、T、C、G平均含量分别为35.4%、26.0%、27.9%、10.7%,表现出明显的反G偏倚;日本鳖、黄沙鳖和杂交鳖分别有5、6、10种单倍型,日本鳖与杂交子代有共享的单倍型Hap 8、Hap 9和Hap 16;3个群体的变异主要存在于群体内部,变异比例为60.8%;3个群体的单倍型多样性为0.747~0.867,平均核苷酸差异数为12.922~16.262,核苷酸多样性为0.009 13~0.011 51;杂交子一代的单倍型数、单倍型多样性、平均核苷酸差异数、核苷酸多样性均最高,日本鳖的均最低,黄沙鳖、杂交子一代、日本鳖的多态位点数依次降低。可见,杂交子代遗传多样性较高,杂交可提高中华鳖的遗传多样性。
黄沙鳖;日本鳖;基因;遗传多样性;单倍型;多态位点
中华鳖隶属爬行纲龟鳖目鳖科,是中国的特色水产养殖品种。由于中华鳖分布广泛,虽无亚种分类,但也形成了各具地理特色的种群[1]。根据水域的不同,大致可将中国养殖的中华鳖分成北方品系、黄河品系、西南品系、洞庭湖品系、鄱阳湖品系、太湖品系、台湾品系、日本品系[2–3]。随着养殖业的迅速发展,养殖户开始进行不同品系间的杂交[4]。广西种(西南品系)中华鳖被称为黄沙鳖,抗性强,但产蛋率较低,生长较慢[5–6]。日本鳖是从日本引进并经过5代选育获得的,具有生长速度快、产蛋率高等特点,但其抗逆性较弱,口感较差[7]。目前,生产上养殖较多的杂交鳖为黄沙鳖(♂)与日本鳖(♀)的杂交一代,其具有日本鳖生长快和黄沙鳖抗病能力强的特点,同时口感比日本鳖的更好。线粒体DNA呈环状,具有母系遗传、几乎不发生重组、进化速度快等特点[8–9],在生物的群体遗传多样性及系统发育研究上应用广泛,是理想的遗传分子标记,已广泛应用于水生生物种群亲缘关系研究[10]。中华鳖线粒体长约17 364 bp,包括37个基因,编码13个蛋白[11],其中细胞色素b基因()进化速度适中,其变异程度足以说明种间的系统发育关系,同时其具有保守性,适用于一些亲缘关系较近群体的遗传结构研究[12]。笔者基于中华鳖线粒体DNA的序列对黄沙鳖、日本鳖及其杂交子代进行研究分析,了解该杂交种的遗传特性,旨在为中华鳖的种质保护及将来的杂交育种提供依据。
1 材料与方法
1.1 材料
供试中华鳖均为稚鳖,黄沙鳖(HS)购自广西平南县金田镇汇源龟鳖养殖场,日本鳖(RB)和杂交鳖(HR)购自江西南丰县鑫壮渔业服务部。3个群体各30只,取其裙边组织,保存于无水乙醇中,于–20 ℃冰箱保存,备用。
1.2 方法
采用全式金DNA提取试剂盒提取中华鳖的裙边DNA,利用中华鳖线粒体通用引物Cyt b–R (ATTCCGGTTTTGGGGATCGG)和Cyt b–F(GTCA ACGCCACAGAATAAGC)[13]对所提DNA进行PCR扩增。反应体系25 μL:包括上游、下游引物各1 μL;DNA模板1 μL;MagicMix3.0为12.5 μL;灭菌水9.5 μL。反应程序:94 ℃预变性3 min;94 ℃变性30 s,54.9 ℃退火150 s,72 ℃延伸5 min,运行34个循环;最后72 ℃下延伸10 min。扩增后产物经1%琼脂糖凝胶电泳,观察到清晰的目的条带后送北京擎科生物科技股份有限公司进行PCR产物双向测序。
1.3 数据分析
将所得的DNA测序结果进行正反链校正,引物剪切后用Clustal W和Chromas比对和分析。运用Dnasp 5.0统计样本的单倍型数()、单倍型多样性(d)、核苷酸多态位点数()、核苷酸多样性(i)、平均核苷酸差异数()。运用Popart构建单倍型网络图。运用MEGA11计算群体间和群体内的遗传距离,基于Jukes–Cantor(JC)模型,采用UPGMA法构建单倍型进化树,并进行1000次的Boot strap检验。运用Arlequin 3.5进行分子方差分析(AMOVA)。
2 结果与分析
2.1 3个中华鳖群体Cyt b的序列特征
通过PCR扩增测序获得90条中华鳖序列,经Clustal W对比剪切后获得1419 bp的有效片段。经分析,碱基A、G、T、C的平均含量分别为35.4%、10.7%、26.0%、27.9%,碱基G的含量明显低于其他3种碱基的,表现出明显的反G偏倚;A+T平均含量为61.4%,C+G平均含量为38.6%(表1)。90条序列共检测到18种单倍型(图1),HR、HS和RB群体分别有10、6和5种单倍型;HS群体与其他群体无共享单倍型,主要单倍型为Hap 1;RB和HR群体共享单倍型为Hap 8、Hap 9和Hap 16,主要单倍型为Hap 8。3种鳖均有不共享的单倍型,可根据特定的单倍型来对它们进行区分。
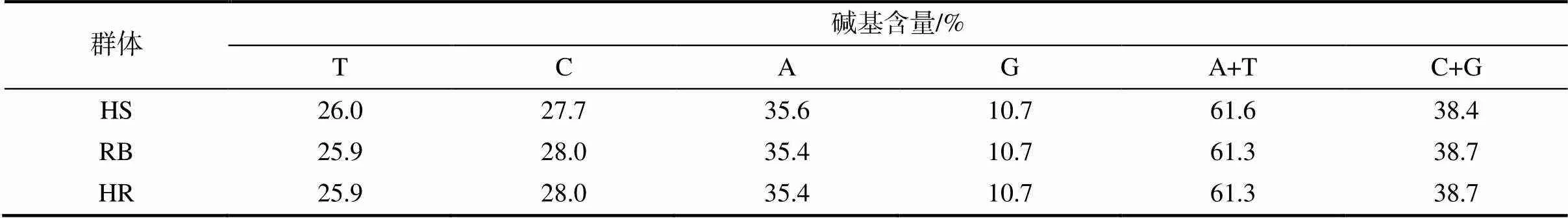
表1 3个中华鳖群体的Cyt b序列碱基组成
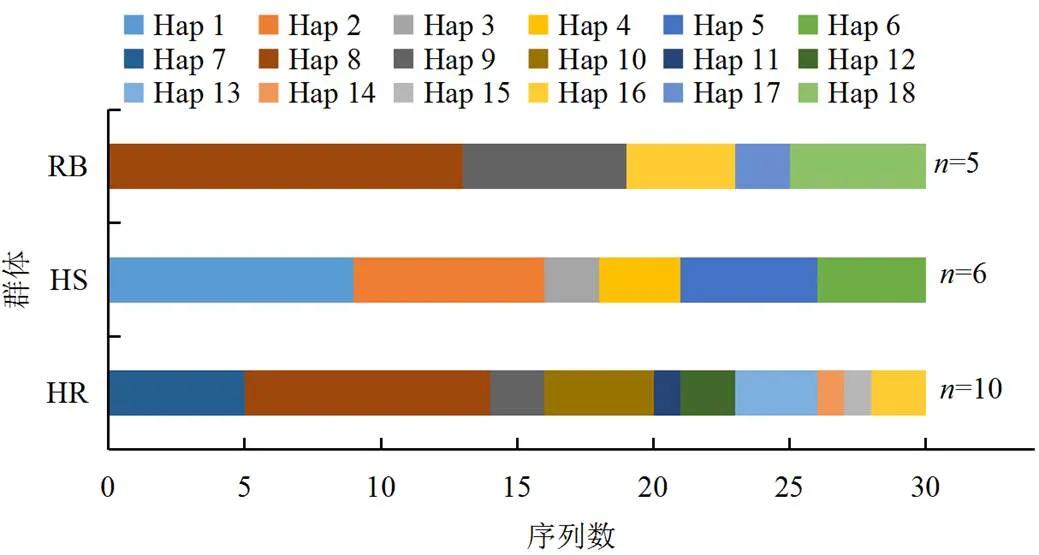
图1 3个中华鳖群体的Cyt b单倍型分布
2.2 3个中华鳖群体的遗传多样性
由表2可知,3个群体中90个个体总的d为0.905,i为0.014 71,为20.749;3个群体中,HS群体的最多,其次为HR群体的,RB群体的最低,分别为54、43、34;3个群体的d为0.747~0.867,为12.922~16.262,i为0.009 13~ 0.011 51,其中HR群体的d(0.867)、(16.262)和i(0.011 51)均最高,表明其遗传多样性较高。
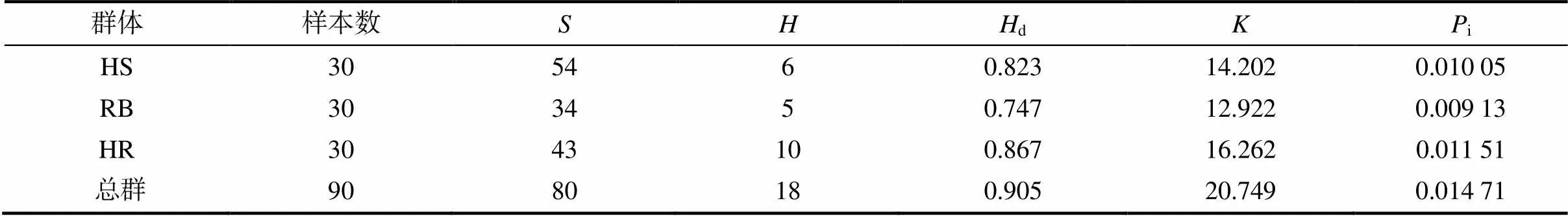
表2 3个中华鳖群体的遗传多样性参数
2.3 3个中华鳖群体的遗传差异
从中华鳖3个群体的遗传距离(表3)来看,群体内遗传距离为0.009 30~0.011 76,群体间遗传距离为0.010 47~0.020 82,其中,HR群体与HS群体间的遗传距离较HR群体与RB群体间的遗传距离更远,符合线粒体基因母系遗传的特点。AMOVA分析结果(表4)显示,3个中华鳖群体60.80%的变异源于群体内,39.20%的变异源于群体间,群体内遗传多样性更丰富。从单倍型网络图(图2)可知,其整体相对较简单,主要分为2支,其中Hap 4与其他单倍型距离较远。在UPGMA法构建的中华鳖单倍型系统发育树(图3)中,部分HR群体与RB群体聚为1支,其余的HR、HS群体与RB群体聚为1支,2支再聚在一起,跟单倍型网络图结果一致。可见,日本鳖与杂交鳖的亲缘关系更近。

表3 3个中华鳖群体间的遗传距离和群体内的遗传距离

表4 3个中华鳖群体的AMOVA分析结果

图2 3个中华鳖群体的Cyt b单倍型网络结构
3 结论与讨论
遗传多样性是衡量生物进化潜力的重要指标。遗传多样性越丰富,物种适应环境的能力越强,高遗传多样性是维持物种生存和进化的物质基础[14–15]。群体遗传多样性每减少10%,就会对该物种的生存力、适应力和繁殖力等造成巨大影响[16]。线粒体DNA特殊的结构和遗传特点使其成为研究物种遗传学的有效工具[17]。NEIGEL等[18]认为,给定群体内线粒体DNA核苷酸多样性越小,群体的遗传多样性越低。GRANT等[19]将不同核苷酸多样性和单倍型多样性分为4种组合类型:低i(<0.005)与低d(<0.500);低i(<0.005)与高d(>0.500);高i(>0.005)与低d(<0.500);高i(>0.005)与高d(>0.500)。本研究中,3种中华鳖群体在碱基的组成上基本相同,且A+T的含量明显高于G+C的含量,这与李璐等[20]的研究结果相似。本研究结果表明,3个中华鳖群体的d与i均较高,属于第4种类型,表明3个中华鳖群体的遗传多样性较丰富,种群可能有一个较大且稳定的群体。
KARTAVTSEV等[21]指出基因一般在种群内的遗传距离为0.005~0.015。本研究中,黄沙鳖与日本鳖的遗传距离为0.020 82,说明黄沙鳖种群与日本鳖种群具有较大差异,可能已分化为2个相对独立的群体。物种遗传多样性的丰富程度与该物种的生存能力相关,遗传多样性越丰富,适应环境的能力越强[22]。单倍型多样性和核苷酸多样性是评价物种多样性的标准[23]。本研究结果显示,黄沙鳖群体遗传多样性较日本鳖遗传多样性更丰富,这与刘阳等[24]的研究结果相似。可能是由于从日本引入群体较少且经过长期人工选育[25]所致。本研究共获得18个单倍型,黄沙鳖、日本鳖和杂交鳖分别有6、5和10个,其中日本鳖和杂交鳖共享3个单倍型,可能与线粒体基因随母本遗传有关[26]。庞广昌等[27]认为群体内的变异影响群体的稳定性,群体间的变异反应群体在不同环境中的适应情况。3个中华鳖群体内的变异占比为60.80%,群体间的变异占比为39.20%,表明各群体之间相对稳定,群体间差异不大。
中华鳖在长期的地理隔离中形成了适应不同环境的地理种,在形态、生长和抗病能力上都有所差异[28–29],利用不同地理种的鳖进行杂交已培育出具有生长快、抗逆性强、营养价值高等特点的杂交鳖,如绿卡鳖[30]、英明中华鳖[31]、浙新花鳖[32–33]等。杂交优势大小与亲本群体之间的遗传差异有关,在一定范围内遗传距离越大,杂交优势就越明显[34]。本研究中,杂交鳖遗传多样性高于黄沙鳖和日本鳖,表明杂交会提高中华鳖的遗传多样性,进而提高其环境适应力。在养殖生产过程中,杂交鳖的生长及抗病能力表现均优于纯种中华鳖,说明杂交优势在地理种之间的杂交是存在的:因此,对于中华鳖的杂交选育需要弄清其遗传背景,对不同地理群体进行遗传距离的测定,科学选育,避免近亲繁育,同时保证亲本群体数量,以确保群体的遗传多样性。
[1] 张振东,刘夕姿,肖友红.我国中华鳖种苗产业调查与发展战略[J].渔业信息与战略,2016,31(2):98–104.
[2] 金晶,刘子明,吕耀平,等.松阳花鳖与中华鳖四个地理种群形态差异比较研究[J].生态科学,2018,37(2):131–137.
[3] 张林,周剑光,张涛,等.3个不同品系中华鳖间外形、核型以及乳酸脱氢酶同工酶表达的差异[J].中国渔业质量与标准,2020,10(6):59–67.
[4] 阙江龙,付辉云,张燕萍,等.中华鳖鄱阳湖品系和黄沙品系及其杂交后代的主成分分析和判别分析[J].安徽农业科学,2022,50(8):77–80.
[5] 何金钊,冯鹏霏,周大颜,等.稻鳖生态种养对黄沙鳖肌肉品质的影响[J].水产养殖,2022,43(2):24–30.
[6] 马梦娇,荆慧娟,符安卫,等.中华鳖腿肉蛋白的理化性质[J].食品与发酵工业,2019,45(22):110–116.
[7] 梁宏伟,曹力欢,罗相忠,等.基于线粒体基因的中华鳖3个品系遗传多样性分析[J].基因组学与应用生物学,2021,40(Z3):2908–2915.
[8] 程磊,何苹萍,韦嫔媛,等.基于线粒体D–loop区和基因分析广西禾花鲤三个群体遗传结构[J].水生生物学报,2021,45(1):54–59.
[9] 彭敏,肖珊,陈晓汉,等.基于线粒体基因的南海北部金钱鱼种群遗传结构分析[J].南方农业学报,2021,52(10):2851–2860.
[10] 邢晶晶.分子遗传标记及其技术在水产生物中的研究与应用[J].水产学杂志,2002,15(1):61–70.
[11] 彭巧玲,蒲友光,王志方,等.中华鳖线粒体基因组序列分析[J].中国生物化学与分子生物学报,2005,21(5):591–596.
[12] 杨彦平,许萌原,马凤娇,等.基于线粒体基因的长江刀鲚群体遗传结构分析[J].江西农业学报,2021,33(8):11–16.
[13] 陈辰,朱新平,李伟,等.一组鳖科动物线粒体全基因组扩增通用引物:CN112501308 A[P].2021–03–16.
[14] 申绍祎,田辉伍,汪登强,等.长江上游特有鱼类红唇薄鳅线粒体控制区遗传多样性研究[J].淡水渔业,2017,47(4):83–90.
[15] 唐首杰,毕详,张飞明,等.基于线粒体DNA基因序列团头鲂3个选育群体的遗传变异分析[J].水产科技情报,2019,46(2):61–68.
[16] 杨伟,代应贵,安丹丹,等.北盘江光唇裂腹鱼种群基因组DNA遗传多样性的AFLP分析[J].海洋渔业,2021,43(4):385–394.
[17] 程亚欣.基于线粒体控制区和细胞色素B基因的江豚种群遗传多样性和遗传结构分析[D].上海:上海海洋大学,2016.
[18] NEIGEL J E,AVISE J C.Application of a random walk model to geographic distributions of animal mitochondrial DNA variation[J].Genetics,1993,135(4):1209–1220.
[19] GRANT W,BOWEN B.Shallow population histories in deep evolutionary lineages of marine fishes:insights from sardines and anchovies and lessons for conservation[J].Journal of Heredity,1998,89(5):415–426.
[20] 李璐,谭舜,王彪,等.基于线粒体D–loop区序列的中华鳖不同群体遗传差异分析[J].水产学杂志,2020,33(5):7–11.
[21] KARTAVTSEV Y P.Divergence atandmtDNA genes on different taxonomic levels and genetics of speciation in animals[J].Mitochondrial DNA,2011,22(3):55–65.
[22] 李大命,孙文祥,许飞,等.高邮湖大银鱼、太湖新银鱼和基因序列多态性分析[J].水产科学,2020,39(2):258–264.
[23] HOUKI S,YAMADA M,HONDA T,et al.Origin and possible role of males in hermaphroditic androgeneticclams[J].Zoological Science,2011,28(7): 526–531.
[24] 刘阳,史燕,朱新平,等.中华鳖5个群体遗传多样性的微卫星分析[J].基因组学与应用生物学,2012,31(2):141–146.
[25] 田镇,陈爱华,曹奕,等.红壳色文蛤选育群体遗传多样性的微卫星分析[J].南方农业学报,2021,52(9):2582–2589.
[26] PERMANA G N,HUTAPEA J H,MORIA S B,et al. Maternal inheritance of yellowfin tuna () in captivity[J].Journal of Environmental Science and Engineering B,2015,4(1):31–36.
[27] 庞广昌,姜冬梅.群体遗传多样性和数据分析[J].林业科学,1995,31(6):543–550.
[28] 张世水,胡火庚,王军花,等.江西省中华鳖种质状况及养殖方式调查[J].南昌大学学报(理科版),2012,36(1):77–81.
[29] 蔡完其,李思发,刘至治,等.中华鳖七群体稚鳖—成鳖阶段养殖性能评估[J].水产学报,2002,26(5):433–439.
[30] 刘阳,龙建杰,刘佳瑶,等.中华鳖洞庭湖群体、黄河群体及其杂交种稚幼体生长对比试验[J].广东农业科学,2012,39(14):156–157.
[31] 范厚勇.英明中华鳖的选育及其养殖性能研究[D].南昌:南昌大学,2013.
[32] 何中央,张海琪,周凡,等.中华鳖“浙新花鳖”[J].中国水产,2017(3):80–83.
[33] 张君,陈露,余鹏,等.中华鳖4个品系营养成分分析与比较[J].水生生物学报,2018,42(4):770–778.
[34] DONG Z,ZHOU E.Application of the random amplified polymorphic DNA technique in a study of heterosis in common carp,L.[J]. Aquaculture Research,1998,29(8):595–600.
Genetic diversity analysis ofand their hybrid from two geographical populations of usingsequence
XIAO Hewei,HU Yazhou,QIN Qin,XU Dajian,LIANG Yixin,LIU Xiaoyan,WANG Xiaoqing*
(College of Animal Science and Technology, Hunan Agricultural University, Changsha, Hunan 410128, China)
To comprehend the genetic diversity within the Chinese soft-shelled turtle(), mitochondrial genes() from two geographical populations ofand their hybrids were sequenced and analyzed. The study yielded a total of 90 sequences with 18 haplotypes among the three sequenced groups of Japanese population, Huangsha population and hybrid population ofvia PCR. Among the 90 individuals of, the average base contents for A, T, C and G was 35.4%, 26.0%, 27.9% and 10.7%, respectively, revealing a distinct anti-G bias. The Japanese population, Huangsha population and hybrid population exhibited 5, 6 and 10 haplotypes, respectively. Notably, the Japanese population shared haplotypes Hap 8, Hap 9 and Hap 16 with the hybrid population. Population variance contributed significantly to overall variation at 60.8%. Haplotype diversity for all three populations ranged from 0.747 to 0.867, average nucleotide differences ranged from 12.922 to 16.262, and nucleotide diversity ranged from 0.009 13 to 0.011 51. In terms of the three groups, the number of haplotypes, haplotype diversity, average nucleotide diversity and nucleotide diversity of the hybrid population were the highest, and those of the Japanese population were the lowwest, while the number of polymorphic loci decreased in the order of the Huangsha population, hybrid population and Japanese population. The study findings underscored the rich genetic diversity of hybrid population of, suggesting that hybridization events augment the genetic diversity within.
Huangsha population of; Japanese population of;gene; genetic diversity; haplotype; variable site
S917.4
A
1007–1032(2023)04–0486–05
10.13331/j.cnki.jhau.2023.04.017
2022–04–02
2023–08–10
国家重点研发计划项目(2018YFD0900200);湖南省水产产业技术体系
肖何威(1997—),男,湖南长沙人,硕士研究生,主要从事水产动物遗传育种研究,X42643934@163.com;*通信作者,王晓清,博士,教授,主要从事水产动物遗传育种研究,wangxiao8258@126.com
肖何威,胡亚洲,秦溱,徐大建,梁艺馨,刘小燕,王晓清.基于序列分析黄沙鳖和日本鳖及其杂交子一代的遗传多样性[J].湖南农业大学学报(自然科学版),2023,49(4):486–490.
XIAO H W,HU Y Z,QIN Q,XU D J,LIANG Y X,LIU X Y,WANG X Q.Genetic diversity analysis ofand their hybrid from two geographical populations of usingsequence [J].Journal of Hunan Agricultural University(Natural Sciences),2023,49(4):486–490.
http://xb.hunau.edu.cn

责任编辑:邹慧玲
英文编辑:柳正
猜你喜欢
贵州畜牧兽医(2023年3期)2023-06-29
农技服务(2023年2期)2023-03-15
海洋通报(2021年1期)2021-07-23
生物学通报(2021年4期)2021-03-16
园林科技(2020年2期)2020-01-18
当代陕西(2019年9期)2019-05-20
特别健康(2018年2期)2018-07-14
学生天地(2016年10期)2016-04-16
中国音乐教育(2015年4期)2015-05-20
癌变·畸变·突变(2014年1期)2014-03-01
