
导入酵母NDI1基因减少鱼藤酮诱导的分化型帕金森病细胞模型的损伤
2023-10-11 01:29沈露茜徐学静叶逸繁申钰琪李红智
基础医学与临床 2023年10期
沈露茜,徐学静,叶逸繁,陈 卓,陈 兰,申钰琪,李红智*
1.首都医科大学附属北京友谊医院 神经内科,北京 100050;2.西南医科大学附属医院 医学检验部,四川 泸州 646000;3.温州医科大学 检验医学与生命科学学院 教育部检验医学重点实验室,浙江 温州 325035
帕金森病(Parkinson’s disease, PD)在神经系统变性疾病的发病中占第二位,目前主要的治疗手段是多巴类药物,但只能改善部分症状,并不能阻止病情进展,更不能治愈。帕金森病的细胞病理特征主要为多巴胺能神经元变性,其发病可能是遗传和环境因素共同作用的结果,分为家族性和散发性,其中约95%为散发性的。线粒体呼吸链复合体Ⅰ(简称复合体1)缺陷可导致帕金森病的病理特征和临床症状[1-3]。线粒体功能障碍学说认为,帕金森病(特别是散发性)的发病机制是由于线粒体复合体Ⅰ活性降低所致[4-5]。这种在散发性帕金森病患者中普遍存在的复合体Ⅰ活性降低显然不全是由于复合体Ⅰ亚基编码基因的某个突变所致[6]。所以相应的治疗策略应该是功能上弥补整个复合体Ⅰ的缺陷。
酵母的线粒体复合体Ⅰ是内NADH脱氢酶(internal NADH dehydrogenase,NDI1)基因表达的NDI1蛋白,虽然由单亚基构成,但有可能同源替代哺乳动物细胞中由45个亚基构成的复合体Ⅰ[7-8]。
本研究采用酵母NDI1的重组腺相关病毒(recombinant adeno-associated virus with NDI1, rAAV-NDI1),以鱼藤酮诱导经分化的人神经细胞系SH-SY5Y建立帕金森病细胞模型,研究酵母NDI1蛋白对人线粒体复合体Ⅰ功能缺陷和细胞病理特征的改善作用。本研究可以为散发型帕金森病的酵母NDI1基因治疗提供研究基础。
1 材料与方法
1.1 材料
1.1.1 细胞:人神经母细胞瘤细胞系SH-SY5Y(美国菌种保藏中心)。
1.1.2 试剂(盒):5型重组腺相关病毒rAAV5-NDI1(委托武汉枢密脑科学技术有限公司包装);抗人α-突触核蛋白(α-synuclein)抗体(BD公司);抗人pS129 α-突触核蛋白抗体(Abcam公司);抗LC3B抗体、抗HA抗体(CST公司);荧光素/荧光素酶化学发光ATP测定试剂盒、MitoSOXTMRed(Thermo Fisher Scientific公司)。
1.2 方法
1.2.1 细胞的分组及处理:分为3组,DMSO+空载组(正常对照组)、鱼藤酮+空载组(诱导模型组)、鱼藤酮+NDI1组(诱导模型的治疗组)。2×105个SH-SY5Y细胞铺于6孔板中,待贴壁后,换为含10 μmol/L全反式维甲酸(all-trans retinoic acid, ATRA)的细胞分化培养液(第0天)。于第2天,将rAAV5-NDI1,以感染复数(multiplicity of infection,MOI)为4×104,加入含ATRA的培养液中,进行感染。于第4天,去病毒,换含ATRA的培养液,此后于第6天、第8天各换1次含ATRA的培养液。于第8天在含ATRA的培养液中加入1 μmol/L鱼藤酮处理24 h。于第9天,收获细胞用于后续的检测。
1.2.2 Western blot检测标签蛋白HA(NDI1)、α-突触核蛋白和pS129 α-突触核蛋白水平:收集细胞,裂解,离心取上清。测定蛋白浓度后,SDS聚丙烯酰胺凝胶电泳,转膜。封闭后,加一抗(1∶1 000的抗HA抗体或α-突触核蛋白(α-synuclein)抗体或pS129 α-突触核蛋白抗体)于4 ℃孵育过夜。然后二抗孵育。最后显影、曝光。
1.2.3 免疫荧光法检测HA(NDI1)、MitoTracker、LC3B的定位及水平:细胞爬片上的细胞,加100 nmol/L的MitoTrackerTMRed于37℃避光孵育30 min。经固定、通透、封闭后加一抗(1∶100的抗人LC3B抗体或HA抗体)于4 ℃ 避光孵育过夜。加Alexa fluo 647标记的二抗室温避光孵育1 h后加DAPI染色。激光共聚焦显微镜观察。
1.2.4 复合体依赖性氧耗的检测:于Oxygraph-2K细胞呼吸仪的仓内加入2×106个细胞/2 mL检测液。加2% 洋地黄皂苷通透细胞膜后,先加入复合体Ⅰ的底物混合物记录复合体Ⅰ依赖性的氧耗,再加入100 μmol/L的哺乳动物(不包括酵母的)复合体Ⅰ抑制剂鱼藤酮,记录抑制内源性复合体Ⅰ后的氧耗,再加入0.225 mol/L的通用性(包括酵母的)复合体Ⅰ抑制剂黄酮,记录抑制外源性酵母复合体Ⅰ后的氧耗,然后加入复合体Ⅱ和Ⅲ的底物混合物,记录复合体Ⅱ和Ⅲ依赖性的氧耗。用DatLab 软件进行数据分析。
1.2.5 ATP合成的检测:收集的细胞(约1×106个)在100 μL ATP提取液中100 ℃加热90 s。10 μL上清液样品或ATP标准品与100 μL ATP检测液混合,用多功能酶标仪进行化学发光检测,根据标准曲线计算样品的ATP水平。除测定未经处理细胞的基础ATP合成外,另外将细胞与15 mg/L的寡霉素在37 ℃下孵育1 h后,测定抑制ATP合酶后的ATP合成。
1.2.6 线粒体内ROS的测定:收集的细胞(约1×106个),加入5 μmol/L的MitoSOXTMRed于37 ℃避光孵育25 min。洗涤、重悬细胞,流式细胞仪采集5 000个细胞分析每个样本的中位数荧光强度(median fluorescence intensity, MFI)。
1.3 统计学分析
2 结果
2.1 分化后SH-SY5Y细胞的增殖状态、转导后NDI1的表达及亚细胞定位
SH-SY5Y细胞ATRA处理2 d后增殖变慢,在处理4 d后增殖基本停止(图1B)。
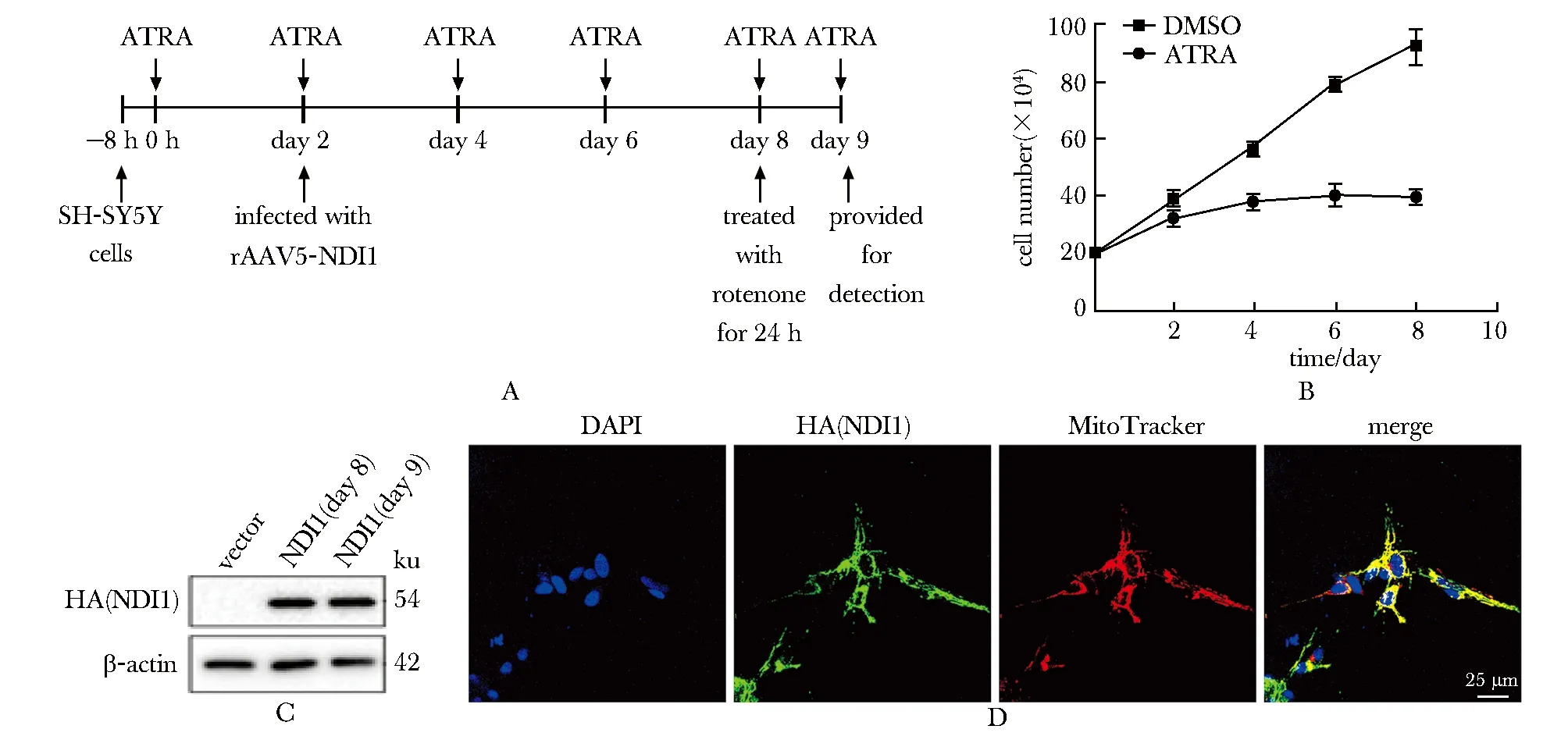
A.scheme for all-trans retinoic aicd (ATRA) differentiation, rotenone treatment and rAAV5-NDI1 transduction of SH-SY5Y cells; B.proliferation status of ATRA treated SH-SY5Y cells; C.expression of HA (NDI1) on day 6 and 7 after transduction(day 8 and 9); D.subcellular co-localization of MitoTracker and HA (NDI1) on day 6 after introduced, MitoTracker(red), mitochondria;HA(green), HA (NDI1) protein; DAPI(blue), nucleus(scale bar:25 μm).图1 分化后SH-SY5Y细胞的增殖状态、感染后NDI1的表达及亚细胞定位Fig 1 Proliferation status of SH-SY5Y cells after differentiation, and the expression and subcellular localization of
在感染后第6、7天,感染NDI1组的HA(NDI1)的表达水平均较高(图1C)。
HA(NDI1)与MitoTracker亚细胞共定位,显示 NDI1蛋白定位于线粒体(图1D)。
2.2 NDI1导入鱼藤酮诱导的分化型帕金森病细胞模型后改善细胞病理学特征
鱼藤酮+空载组细胞形态发生异常改变,胞体变圆并聚集在一起,轴突缩短,细胞间连接变少;鱼藤酮+NDI1组形态明显改善,胞体皱缩现象改善,数量增多,细胞伸展,突起变长(图2A)。
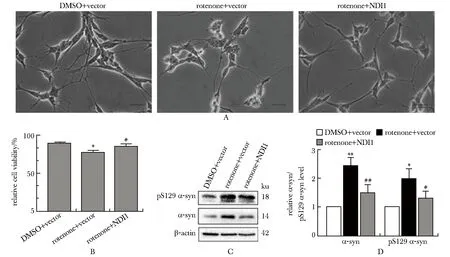
A.cell morphology (scale bar: 100 μm); B.cell viability; C.the α-synuclein (α-syn) and pS129 α-synuclein (pS129 α-syn) levels; D.quantitation of α-synuclein and pS129 α-synuclein levels; *P<0.01, **P<0.001 compared with DMSO+vector; #P<0.05, ##P<0.01 compared with rotenone+vector.图2 NDI1导入鱼藤酮诱导的分化型帕金森病细胞模型后能改善细胞病理学特征Fig 2 NDI1 improved the cytopathological characteristics after introduced into rotenone-induced differentiated Parkinson’s disease cell n=3)
鱼藤酮+空载组与DMSO+空载组相比,细胞存活率显著下降(P<0.01),鱼藤酮+NDI1组较鱼藤酮+空载组显著回升(P<0.05)(图2B)。
鱼藤酮+空载组较DMSO+空载组α-突触核蛋白和pS129 α-突触核蛋白水平显著增加(P<0.001,P<0.01),鱼藤酮+NDI1组较鱼藤酮+空载组显著回落(P<0.01,P<0.05)(图2D)。
2.3 NDI1导入鱼藤酮诱导的分化型帕金森病细胞模型后恢复线粒体复合体Ⅰ依赖性的氧耗水平
鱼藤酮+空载组较DMSO+空载组复合体Ⅰ依赖性氧耗显著降低(P<0.001),鱼藤酮+NDI1组较鱼藤酮+空载组显著回升(P<0.01)(图3A)。
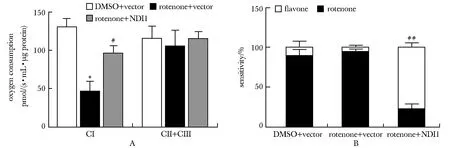
A:CI-dependent and CⅡ+CⅢ-dependent oxygen consumption levels, CI.complex Ⅰ, CⅡ.complex Ⅱ, CⅢ.complex Ⅲ; B:proportion of flavone or rotenone sensitive in complex Ⅰ-dependent oxygen consumption; *P<0.001 compared with DMSO+vector; #P<0.01, ##P<0.001 compared with rotenone+vector.图3 NDI1导入鱼藤酮诱导的分化型帕金森病细胞模型后能恢复线粒体复合体Ⅰ依赖性的氧耗水平Fig 3 NDI1 restored mitochondrial complex Ⅰ-dependent oxygen consumption level after transduction into rotenone-induced differentiated Parkinson’s disease cell n=3)
鱼藤酮+NDI1组较鱼藤酮+空载组,复合体Ⅰ依赖性氧耗中对黄酮敏感的比例显著升高(P<0.001),占复合体Ⅰ依赖性氧耗的大部分(图3B)。
2.4 NDI1导入鱼藤酮诱导的分化型帕金森病细胞模型后能恢复ATP水平
鱼藤酮+空载组较DMSO+空载组细胞整体ATP合成量(即base)显著降低(P<0.001),鱼藤酮+NDI1组较鱼藤酮+空载组显著回升(P<0.01)(图4A)。

A.total cellular ATP content (base), oligomycin-sensitive ATP content (oligo-sensitive); B.proportion of oligomycin-sensitive ATP content; *P<0.001 compared with DMSO+vector; #P<0.01 compared with rotenone+vector.图4 NDI1导入鱼藤酮诱导的分化型帕金森病细胞模型后能恢复ATP水平Fig 4 NDI1 restored ATP level after transduction into the rotenone-induced differentiated Parkinson’s disease
鱼藤酮+空载组较DMSO+空载组对寡霉素敏感的ATP合成量(即oligo-sensitive)及其比例显著降低(P<0.001),鱼藤酮+NDI1组较鱼藤酮+空载组显著回升(P<0.01)(图4A,B)。
2.5 NDI1导入鱼藤酮诱导的分化型帕金森病细胞模型后能降低ROS水平
鱼藤酮+空载组较DMSO+空载组线粒体内ROS水平显著升高(P<0.001),鱼藤酮+NDI1组较鱼藤酮+空载组显著回落(P<0.01)(图5)。
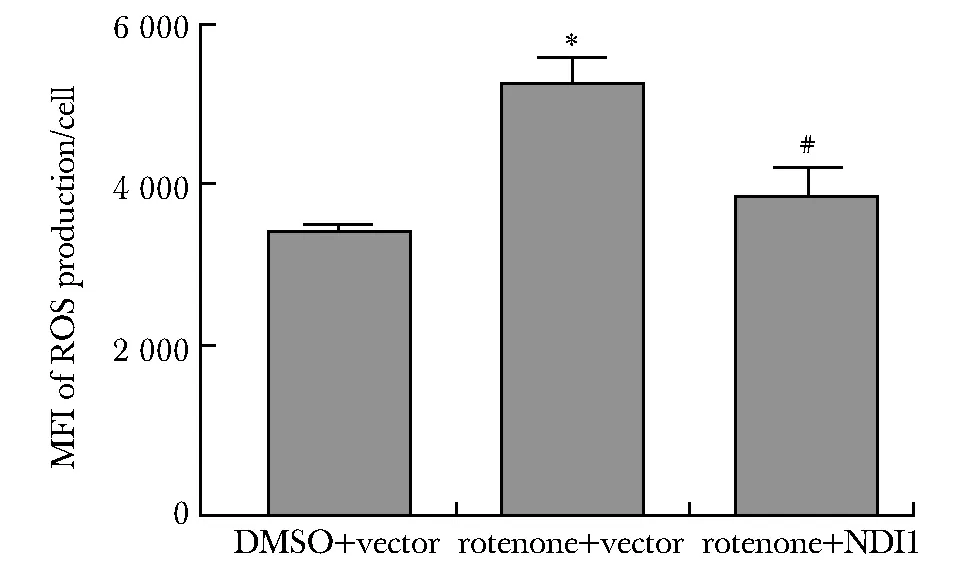
MFI.median fluorescence intensity; *P<0.001 compared with DMSO+vector; #P<0.01 compared with rotenone+vector.图5 NDI1导入鱼藤酮诱导的分化型帕金森病细胞模型后能降低ROS水平Fig 5 NDI1 reduced the level of ROS after transduction into the rotenone-induced differentiated Parkinson’s
2.6 NDI1导入鱼藤酮诱导的分化型帕金森病细胞模型后能降低细胞自噬和线粒体自噬水平
鱼藤酮+空载组较DMSO+空载组自噬标志物LC3B免疫荧光强度显著提高(P<0.001),鱼藤酮+NDI1组较鱼藤酮+空载组显著回落(P<0.001)(图6B)。
鱼藤酮+空载组较DMSO+空载组LC3B与Mito-Tracker共定位的Pearson相关系数显著提高(P<0.001),鱼藤酮+NDI1组较鱼藤酮+空载组显著回落(P<0.001)(图6C)。
3 讨论
鱼藤酮是常用的构建帕金森病细胞、动物模型的神经毒素,其主要机制是抑制线粒体复合体Ⅰ[9]。本研究采用鱼藤酮来构建帕金森病细胞模型,研究结果显示复合体Ⅰ相关的线粒体功能低下并出现了帕金森病的细胞病理学特征(图2~6),表明构建成功。
目前针对线粒体复合体Ⅰ缺陷的基因治疗,如 Leber氏视神经病的ND4基因治疗已完成临床前研究并进入临床试验[10],还有对Ndufs4全身性敲除的Leigh综合征模型小鼠采用AAV-Ndufs4进行基因治疗[11],但已报道基因治疗策略只针对人复合体Ⅰ某一个亚基功能缺陷。本研究的基因治疗策略能够在功能上弥补整个复合体Ⅰ的缺陷,有望应用于所有线粒体复合体Ⅰ缺陷的疾病。
已有一些研究证实了酵母NDI1或其蛋白产物用于复合体Ⅰ缺陷相关疾病治疗的有效性。采用体外制备的细胞通透性TAT-NDI1蛋白,能够使心肌缺血损伤模型大鼠的心肌复合体Ⅰ功能恢复,心梗减小[7]。在多发性硬化模型小鼠中采用NDI1靶向功能缺陷的复合体Ⅰ,能够缓解轴突受损和神经元缺失问题,从而改善视觉功能[8]。本研究结果表明, 细胞水平的NDI1治疗可以抵抗鱼藤酮诱导的细胞形态改变和存活率下降,使复合体Ⅰ(主要是外源性的)依赖性氧耗、线粒体有关的ATP显著回升,使线粒体ROS显著下降并抵抗线粒体自噬(图2~6),具有与上述文献报道类似的改善效果。
本研究采用带有线粒体导肽序列的酵母NDI1,先进入细胞核中,在细胞质中翻译后转运到线粒体中。已有研究表明,酵母NDI1在大鼠中没有引起免疫反应,这可能因为外来蛋白位于线粒体中,可以躲避免疫监测之故[12]。
临床试验已证明腺相关病毒AAV在帕金森病等神经组织基因治疗中具有令人鼓舞的安全性、有效性以及表达的持久性[13-14],故本研究采用AAV作为基因治疗表达载体。本研究先采用全反式维甲酸分化SH-SY5Y细胞,停止细胞分裂,然后采用rAAV-NDI1感染分化型帕金森病细胞模型。因为重组腺相关病毒在被感染的细胞中主要是以非整合的形式存在,细胞分裂后的子代细胞中重组基因含量将被稀释掉[15]。
猜你喜欢
保健医苑(2022年6期)2022-07-08
活力(2019年22期)2019-03-16
深圳职业技术学院学报(2018年3期)2018-07-23
中国野生植物资源(2018年1期)2018-04-23
原子与分子物理学报(2015年1期)2015-11-24
医学研究杂志(2015年5期)2015-06-10
中国中医药现代远程教育(2014年11期)2014-08-08
中国药理学通报(2014年2期)2014-05-09
湖南农业科学(2014年5期)2014-02-27
食品科学(2013年15期)2013-03-11
