
乳酸对人非小细胞肺癌细胞A549生物学特性影响的研究
2023-12-27 09:09黄登亮张耀刚田美媛李志琴童思贤
解放军医学院学报 2023年9期
孙 莉,江 源,黄登亮,侯 静,张耀刚,田美媛,李志琴,童思贤,姜 军
1 青海大学附属医院中心实验室,青海西宁 810001;2 青海大学附属医院科研管理部,青海西宁 810001;3 青海大学附属医院肿瘤内科,青海西宁 810001
肺癌是一种常见的呼吸道肿瘤,在我国肺癌新增病例和死亡人数均居所有恶性肿瘤之首[1]。肺癌分为小细胞肺癌和非小细胞肺癌(non-small cell lung cancer,NSCLC),其中NSCLC最为常见,占所有肺癌患者的80% ~ 85%[2]。虽然手术、放化疗、生物免疫疗法、分子靶向药物等疗法可延长部分肺癌患者的生存期,但治疗的效果较为有限,肺癌的总体5年生存率目前仍在15%左右[3],因此亟须探索肺癌诊疗的新思路和新策略。肿瘤的重要特征之一是代谢异常[4],肿瘤细胞代谢重塑引起乳酸累积,肿瘤组织中乳酸浓度可高达20 ~30 mmol/L,平均浓度约10 mmol/L[5]。研究显示,乳酸可作为代谢供能物质参与NSCLC细胞三羧酸循环。在急性炎症模型小鼠中,乳酸诱导内质网中Mg2+释放而参与急性炎症过程中的组织损伤[6]。此外乳酸通过组蛋白乳酸化修饰调节巨噬细胞基因表达且改变细胞特性[7],上调小胶质细胞中糖酵解基因表达促进阿尔茨海默病进展[8]。因此,乳酸可作为代谢底物、信号分子和表观遗传修饰因子发挥重要生物学功能。乳酸作为代谢产物,对NSCLC细胞糖酵解是否具有反馈调节作用还不明确。本研究将探索乳酸对NSCLC细胞A549糖酵解的影响,并分析乳酸处理后A549细胞生物学特性的变化情况。
材料与方法
1 材料 人非小细胞肺癌细胞A549细胞株购于上海中科院细胞库。乳酸(货号:L6402)购自默克生命科学公司;DMEM(货号:2230805)购自Gibco公司;糖酵解压力检测试剂盒(货号:103020-100)和细胞增殖E-Plate检测板(货号:00300600890)购自安捷伦公司;HK1一抗(货号:#2024)和PKM2一抗(货号:#4053)购自CST公司;G6PD一抗(货号:ab210702)和LDHA一抗(货号:ab52488)购自Abcam公司;β-actin一抗(货号:sc-47778)购自Santa Cruz公司。
2 细胞培养及处理 人非小细胞肺癌细胞A549用含10%胎牛血清的RPM 1640培养基在37°C、5% CO2培养箱中培养。细胞贴壁后,分别加入终浓度为5 mmol/L和10 mmol/L的乳酸处理A549细胞24 h后,收集细胞并检测糖酵解压力、线粒体底物选择以及细胞增殖、迁移和周期等生物学特征。
3 细胞代谢分析仪(Seahorse XFe96)检测糖酵解压力和线粒体底物选择 每孔接种80 µL A549细胞悬液(细胞数为1.2 × 104),糖酵解压力测定实验乳酸浓度设置为0 mmol/L、5 mmol/L、10 mmol/L;分析线粒体底物选择时乳酸浓度设置为0 mmol/L、10 mmol/L,每组设置3个复孔;37°C、5% CO2培养24 h后,按照糖酵解压力检测试剂盒和线粒体底物选择分析试剂盒说明书实时测定各组活细胞在生理温度(37°C)下的耗氧率和细胞外酸化率。细胞外酸化率是糖酵解的关键指标,可在不损伤细胞的情况下了解培养细胞糖酵解压力水平;线粒体底物选择分析实验对于参与细胞氧化的3种主要线粒体底物(葡萄糖、谷氨酰胺和长链脂肪酸),利用特异性抑制剂的不同组合,分析其依赖性、容量和灵活性。检测完成后,用Hoechst 33258标记细胞核并计数,对糖酵解压力分析结果进行标准化处理。
4 实时无标记细胞分析(real time cellular analysis,RTCA)系统检测细胞的迁移与增殖能力 细胞株在进行细胞增殖实验前1 d进行细胞传代,实验时满足细胞满度在60% ~ 80%,从培养箱中取出细胞,进行消化、离心后,使用完全培养基配制成实验需要的细胞浓度;检测E-Plate 16 VIEW板基线后,将细胞加入E-Plate 16 VIEW孔板;用于增殖和迁移检测的细胞接种量分别为1.2 × 104/孔和5 × 104/孔。乳酸浓度设置为0 mmol/L、5 mmol/L、10 mmol/L;每组设置3个复孔。接种细胞后,将E-Plate 16 VIEW孔板在超净台中室温放置30 min使细胞充分沉降,再将E-Plate 16 VIEW孔板放到RTCA仪器中。编辑样本信息和测定条件,开始实时动态检测细胞迁移和增殖曲线。
5 qPCR和Western blot检测相关基因和蛋白表达 提取RNA后,以RNA为模板反转录合成cDNA,进行PCR扩增反应。引物序列由上海生工公司合成,并读取Ct值,以2-△△Ct表示目的基因mRNA的相对表达水平;qPCR引物序列见表1。Western blot测定相关蛋白表达水平,细胞经裂解后取总蛋白,进行10% SDS-PAGE电泳,湿转法将蛋白转移至PVDF膜,5%脱脂奶粉封闭,4℃孵育过夜,TBST洗膜,加入含5%脱脂奶粉的PBS和一抗,4℃孵育过夜;TBST洗膜,加二抗,37℃孵育1 h;TBST洗膜,ECL化学发光显影,成像系统采集图像并分析目标条带和内参条带。
6 流式细胞仪检测细胞周期 每支流式管中加入1 × 106个细胞,加入1 mL 70%乙醇,涡旋混匀,4℃固定30 min,450 g离心5 min,弃上清,加1 mL PBS重悬细胞,450 g离心5 min后倒掉上清,加100 µL PBS,再加入1 µL RNase A (10 mg/mL)和5 µL PI(终浓度50 µg/mL),涡旋混匀,避光孵育15 min,加1 mL PBS重悬细胞,450 g离心5 min,弃上清,加入400 µL PBS重悬上机。
7 统计学分析 利用SPSS 19.0软件进行数据统计。计量资料以表示,所有实验均重复3次,符合正态分布与方差齐性的两组间计量资料比较采用独立样本t检验,多组间差异比较采用单因素方差分析,进一步两两比较采用Tukey’s检验,依时间变化资料采用重复测量方差分析。P<0.05为差异有统计学意义。
结 果
1 乳酸对A549细胞糖酵解水平和线粒体底物选择的影响 由Seahorse XFe96细胞代谢分析仪实时监测细胞外酸化率所得的糖酵解功能曲线(图1A)和参数指标的计算公式(图1B)可计算出细胞的糖酵解水平、糖最大酵解能力和糖酵解储备等参数。结果显示,5 mmol/L和10 mmol/L乳酸处理后A549细胞的糖酵解水平(F=84.85,P<0.000 1;图1C)、最大糖酵解能力(F=334.6,P<0.000 1;图1D)和糖酵解储备(F=233.7,P<0.000 1;图1E)均降低,且加入乳酸浓度越高,糖酵解能力下降越显著。因此,乳酸对A549细胞的糖酵解代谢具有负性调节作用。由于糖酵解和线粒体代谢紧密关联,在观察到乳酸对糖酵解的影响后,进一步通过线粒体底物选择分析乳酸对线粒体代谢的影响;线粒体底物选择的结果显示:10 mmol/L乳酸处理后A549细胞对葡萄糖(图1F)的利用无显著改变,对脂肪酸的依赖性增强(图1H)(P=0.018),对谷氨酰胺(图1G)的氧化能力减弱(P=0.019);提示在A549细胞代谢中,乳酸可能在有些方面具有与谷氨酰胺相似的功能。因此,乳酸对A549细胞的糖酵解有负性调节作用,且引起线粒体代谢底物改变。
2 qPCR和Western blot检测糖酵解相关基因表达水平 为探索乳酸影响糖酵解水平的分子机制,我们分析了部分葡萄糖代谢相关基因的表达水平。蛋白免疫印迹结果显示,乳酸处理后的A549细胞中G6PD的蛋白水平显著下降(P=0.007),且10 mmol/L乳酸处理组下降较5 mmol/L乳酸处理组更为显著。 随着乳酸处理的浓度升高,PKM2和HK1的蛋白水平微弱降低,LDHA蛋白水平略微升高(图2A ~ 图2B),但差异无统计学意义。qPCR结果显示,乳酸处理后的A549细胞中HK3(P=0.048)、LDHA(P<0.001)、LDHB(P<0.001)(图2C)基因mRNA水平均降低,且乳酸处理浓度越高,上述基因表达下降越显著。qPCR实验中10 mmol/L乳酸处理的A549细胞HK3基因Ct值太大无法进行统计分析,但也说明10 mmol/L乳酸处理后A549细胞中HK3基因mRNA水平下调。而PKM基因mRNA水平在两个乳酸处理组的A549细胞中均上调(P<0.001),可能是PKM蛋白水平下降引起的反馈调节所致。上述结果显示,乳酸引起葡萄糖代谢酶基因表达下调,可能是乳酸处理组A549细胞糖酵解水平下降的原因。
3 乳酸对A549细胞的迁移、增殖和周期的影响增殖和迁移能力是肿瘤细胞的重要生物学特征。RTCA系统检测结果显示,与对照组相比,5 mmol/L和10 mmol/L乳酸处理后,A549细胞迁移能力减弱(F=177.3,P<0.001;图3A);随乳酸处理浓度升高,A549细胞增殖减弱(F=646.7,P<0.001;图3B)。以上结果提示,乳酸能够降低A549细胞的增殖和迁移能力。进一步通过流式细胞术测定比较正常培养A549细胞(图3C)和10 mmol/L乳酸处理组A549细胞(图3D)的细胞周期变化,结果显示相对于正常培养A549细胞,10 mmol/L乳酸处理的A549细胞G1期细胞增多(t=5.145,P=0.001),S期(t=4.994,P=0.001)和M期(t=3.569,P=0.007)细胞减少(图3E)。因此,A549细胞的迁移和增殖能力随乳酸处理浓度增加而下降。
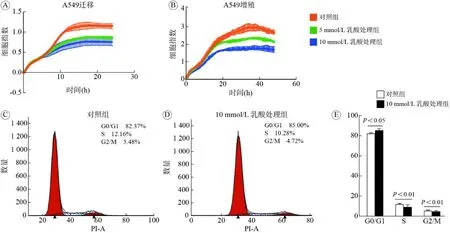
图3 乳酸处理后A549细胞迁移、增殖曲线和细胞周期分析结果A:细胞迁移曲线;B:细胞增殖曲线;C:对照组细胞周期图;D:10 mmol/L乳酸处理组细胞周期图;E:细胞周期分析柱状图Fig.3 Migration and proliferation curve and cell cycle analysis results of A549 cells after lactic acid treatmentA: Cell migration curve; B: Cell proliferation curve; C: The distribution of cell cycle in the control group; D: The distribution of cell cycle in the 10 mmol/L lactate treatment group; E: Analysis of the proportion of cells in the G0/G1 phase, S phase and G2/M phase in the control group and the treatment group
4 细胞周期与迁移相关基因mRNA水平 采用qPCR对细胞周期与迁移相关基因mRNA水平进行分析,发现5 mmol/L乳酸处理组参与细胞周期G1/S转换的CCND1(图4B)以及参与细胞周期G2/M转换的CCNB1(图4A) mRNA水平受到抑制(P<0.05),10 mmol/L乳酸处理组参与细胞周期G1/S转换的CCNE1(图4C)、CCND1和CCNB1的mRNA表达下降更为显著(P<0.05);而两个乳酸处理组中CDK2较对照组mRNA水平均上调,尤其是5 mmol/L上调最为显著(P<0.05;图4D)。CDK2是由G1期进入S期的关键性细胞周期蛋白依赖性激酶,其mRNA水平升高可能是CCNE1、CCND1和CCNB1抑制所引起的。分析细胞迁移相关基因mRNA表达水平发现,与对照组相比,两个乳酸处理组A549细胞中CDH1基因mRNA水平均升高(P<0.05;图4E),10 mmol/L乳酸处理组中TWIST1基因mRNA水平显著下调(P<0.05;图4G),三组SNAI2基因mRNA水平无显著变化(图4F)。CDH1基因编码的E-钙黏蛋白抑制细胞迁移,而TWIST1促进细胞迁移。以上结果显示,乳酸对A549细胞增殖和迁移的抑制作用可能通过调控细胞增殖、迁移相关基因表达而实现。
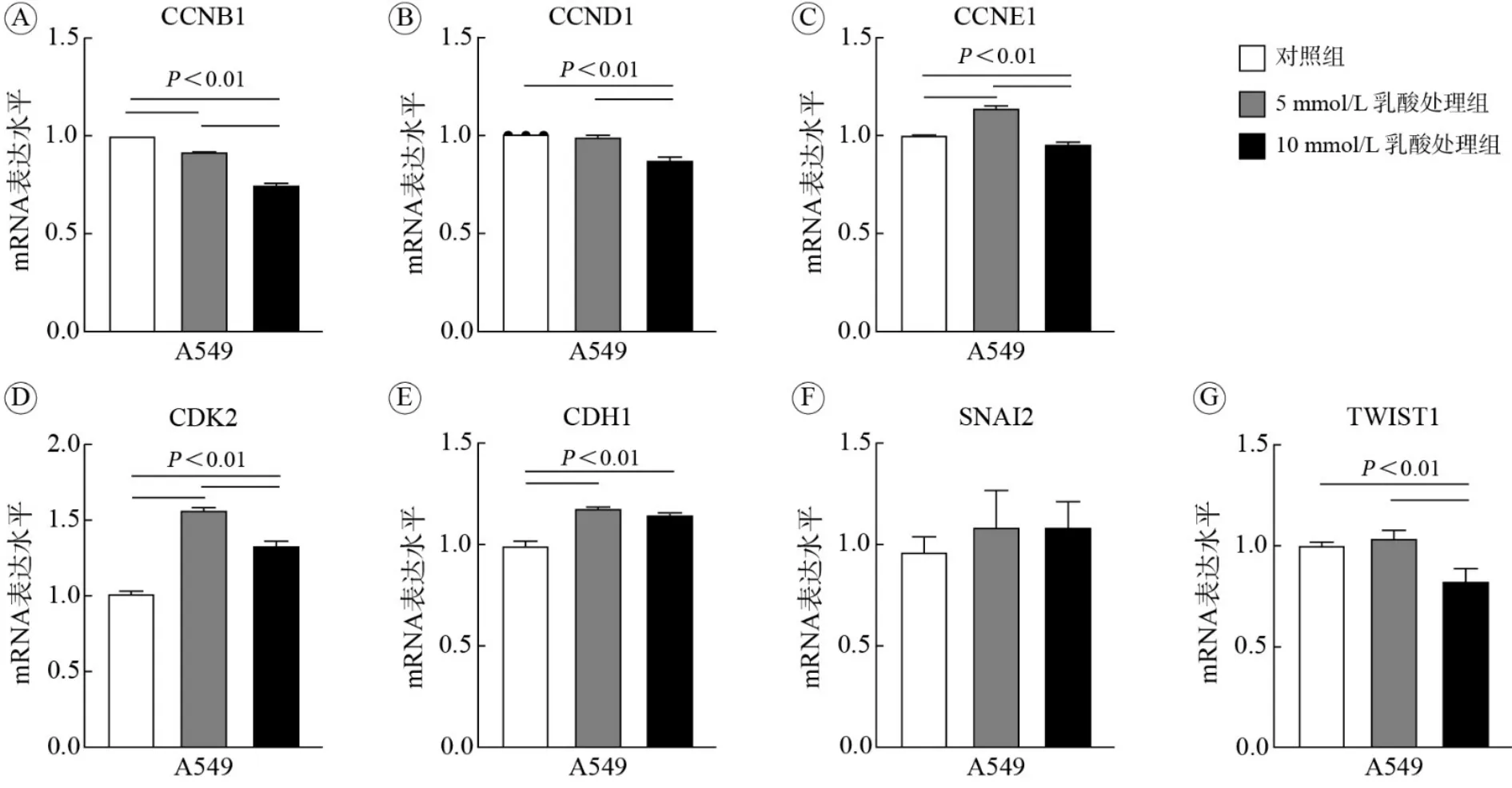
图4 乳酸处理后细胞周期与迁移基因mRNA表达水平Fig.4 Cell cycle and migration gene mRNA expression levels after lactate treatment
讨 论
当前,肿瘤已成为危害人类健康和生命安全的头号杀手,基于性别、地域和年龄等流行病学因素,肺癌是全球癌症相关死亡的首要原因[9],代谢重编程长期被认为是肿瘤的标志之一[10]。约100年前,有研究者发现肿瘤细胞的糖代谢方式就是在有氧条件下的高糖酵解水平,产生大量乳酸,这一现象被称为有氧糖酵解或Warburg效应,大量科学家在癌症糖酵解和乳酸代谢领域进行了更深入的研究。尽管糖酵解途径产生的能量要比三羧酸循环产生的能量少得多,但糖酵解依然是肿瘤细胞的主要代谢方式,为合成代谢提供了多种糖酵解中间体,满足了癌细胞快速增长的需求[11],与正常细胞相比这种代谢方式的改变导致的结果就是糖酵解的产物乳酸在肿瘤中的积聚,而长期以来乳酸对肿瘤细胞的作用并不明确。近年大量研究揭示了乳酸在肿瘤中的作用,其在肿瘤的增殖、侵袭、转移、免疫逃逸、代谢、衰老以及干细胞分化等过程中扮演着重要的角色[12-18]。乳酸还可以刺激内皮细胞产生血管内皮生长因子,促进肿瘤细胞迁移和刺激血管生成,乳酸的浓度与放疗抵抗呈正相关。乳酸与疾病预后呈负相关,高乳酸水平是晚期肺癌患者死亡的独立危险因素[19-20]。
本研究以人非小细胞肺癌 A549 细胞为研究对象,检测并研究乳酸对A549糖酵解压力、细胞增殖、迁移和周期的影响。结果发现,乳酸负调控A549细胞糖酵解水平,并引起线粒体代谢底物选择变化。最新研究显示,巨噬细胞中乳酸通过共价修饰糖酵解、磷酸戊糖途径和三羧酸循环等代谢途径中的酶调节细胞代谢过程[21],负反馈调节糖酵解代谢酶活性,本研究发现乳酸可能通过调节基因表达负调控A549细胞糖酵解;因此,乳酸对糖酵解的负调控作用具有普遍性,并可能通过不同调节方式实现。线粒体底物选择分析实验中,乳酸处理后A549细胞对谷氨酰胺的利用减少,谷氨酰胺是回补三羧酸循环的重要代谢物[18],由于乳酸也可通过代谢转化进入三羧酸循环,所以乳酸会减少A549细胞对谷氨酰胺的利用。除影响细胞代谢外,本研究还观察到乳酸抑制A549细胞增殖和迁移,这似乎与乳酸对肿瘤发生发展的促进作用相矛盾,但肿瘤细胞具有持续增殖、侵袭能力增强、免疫逃逸和代谢重塑等多种重要特征,仅根据增殖迁移能力的改变并不能反映乳酸对肿瘤细胞的整体作用。此外,处于增殖、迁移等不同状态的细胞对营养物质和能量的需求不同,而物质能量变化通过细胞代谢实现,所以细胞的增殖和迁移能力与细胞代谢状态密切关联,本研究中糖酵解水平下降与细胞增殖迁移能力减弱之间的联系值得进一步探究。此外,我们观察到乳酸引起细胞代谢、增殖迁移和相关基因表达的变化,而乳酸引起基因表达变化的分子机制还不明确。
综上所述,外源乳酸引起A549细胞糖酵解能力降低、线粒体代谢底物选择发生改变、细胞增殖及迁移能力减弱,这种变化可能通过调节相关基因表达实现。但糖酵解水平下降与细胞增值迁移能力减弱之间的联系,以及乳酸影响相关基因表达的分子机制还需进一步探索。
作者贡献姜军:项目监督指导;孙莉:总体构思,撰写初稿;江源:方法设计;黄登亮:调查设计;候静:资源提供;张耀刚:数据管理;田美媛:调查设计;李志琴、童思贤:项目管理。
利益冲突所有作者声明无利益冲突。
数据共享声明本论文相关数据可依据合理理由从作者处获取,Email:747495351@qq.com。
猜你喜欢
中南医学科学杂志(2022年6期)2022-12-04
云南化工(2021年6期)2021-12-21
科学(2020年2期)2020-08-24
中国循证心血管医学杂志(2020年11期)2020-01-08
国际呼吸杂志(2019年4期)2019-03-12
中华老年口腔医学杂志(2016年4期)2017-01-15
中国医学装备(2016年6期)2016-12-01
中华老年多器官疾病杂志(2016年7期)2016-04-28
癌症进展(2016年10期)2016-03-20
医学研究杂志(2015年5期)2015-06-10
