
泥石流滩地引洪淤地改造后种植玉米对土壤细菌群落结构特征的影响
2023-12-29 01:11刘永红陈冠陶闫洋洋张建华
西南农业学报 2023年10期
王 谢, 刘 勤, 刘永红, 李 芹, 李 明, 陈冠陶,闫洋洋, 唐 甜, 张建华
(1. 四川省农业科学院农业资源与环境研究所/农业农村部西南山地农业环境重点实验室,成都 610066;2. 中国科学院、水利部成都山地灾害与环境研究所,成都 610299;3. 四川省农业科学院,成都 610066)
【研究意义】泥石流滩地是泥石流暴发后形成的泥石流堆积物,也是山区一种特殊的土地资源。在条件允许和需要的情况下,泥石流滩地值得开发利用,可为生态建设提供有效的缓冲空间,成为山区破解耕地保护和发展保障难题的有效途径之一[1]。我国山区众多,泥石流广泛分布于我国各大山区。特别在我国西南山区,泥石流分布尤为广泛。泥石流滩地主要分布于流域内地势平缓、海拔高度相对较低的区域,具有地势平坦、灌溉便利、光照充足、热量丰富等优点,其生产潜力较高、开发前景较大[2]。【前人研究进展】近年来对泥石流滩地的研究引起了国内外学者的广泛关注,对国内尤其是云南小江流域蒋家沟泥石流滩地的开发利用研究较多[3-4]。中国地学学者对泥石流滩地的研究起步较晚,20世纪80年代主要集中于泥石流滩地的风险判别与分析阶段,研究滩地利用环境的安全性,后期分析和论证了滩地土壤性质及其作为耕地的生产潜力[5-7]。滩地演化一般经历3个时期:初始形成期、发展期和成熟期(又称稳定期)[8],初始形成的未经改造的泥石流滩地,土壤层次结构不合理,物理结构性差,保水保肥能力极差,养分贫瘠,无法作为耕地利用。针对泥石流滩地砾石含量多,黏粒及粉砂粒含量低,土壤结构性和可耕性差等特点,主要有以下3个措施改良土壤:①耕种前人工捡出石块,增加土壤的易耕性,改善土壤组成结构。②整地以后,从附近搬运熟土进行客土改良,增加壤质土成分,形成初步可耕作的土壤。③在汛期降雨后,利用沟道水流中挟带的细颗粒泥沙和养分进行淤地改良,增加滩地的细颗粒成分,改良土壤结构,并补充养分。20世纪90年代,云南蒋家沟泥石流滩地经开发利用,增加耕地超280 hm2,水稻产量超1×104kg/hm2,人均粮食增加近7倍,极好地解决了温饱问题[9]。可见,泥石流滩地经过科学地改良、利用,具有巨大的生产潜力和经济效益。【本研究切入点】山区人民由于缺乏对泥石流滩开发利用的深刻认识,在滩地开发利用过程中,特别是作为农耕地的使用过程中,往往只是进行粗糙利用,缺乏科学的整地、改土以及培肥策略。目前,仅有少数学者对泥石流滩地改造利用过程中土壤理化性质、养分特征等做了初步探讨,对指导滩地利用有一定价值,但缺乏系统深入的阐述。土壤微生物在根土界面种对土壤发育和作物生长都有重要影响。泥石流滩地改造后微生物的响应是深入揭示滩地土壤发育机理和生态系统元素循环的关键环节,然而该部分研究近乎空白。【拟解决的关键问题】本研究从泥石流滩地土壤微生物学的角度,通过客土、引洪漫淤的方式对泥石流滩地土壤进行改造后种植玉米,研究泥石流滩地土壤细菌群落结构的变化及内在机理,寻求提高泥石流滩地土壤耕性和生产潜力的改造方式,为泥石流滩地的开发利用提供科学依据。
1 材料与方法
1.1 研究区概况
四川凉山州喜德县红莫镇桃源村泥石流滩地(102.28° E,28.11° N)海拔1844 m,该地气温年差较小,常年平均气温15.0 ℃,7月平均气温21.60 ℃,1月平均气温8.80 ℃,四季不分明,但气温日差较大。研究区降水集中在5—9月,占全年总降雨量73.9%;常年平均降雨量1006.10 mm,最高达1237.80 mm;1/6、1、6和24 h的最大暴雨量平均值分别为7.50、21.00、32.00和63.00 mm。
热水河流域面积161.40 km2,河长36 km,比降67.3%,年平均流量2.46 m3/s。研究地所在泥石流滩地位于主河道热水河左侧,属于老泥石流灾害点。该泥石流灾害点包括3条泥石流沟,分别为邓家湾沟、杨湾沟和桃源沟,均为热水河右岸的支沟。这3条支沟泥石流物源丰富,皆超过7×105m3,可能参与泥石流活动的动储量近1×105m3,历史上均曾发生多起大型泥石流。研究区所在泥石流滩地右岸主河道流量一般在1.82~5.40 m3/s,左岸小河沟流量,枯水期为干沟,丰水期流量可达2.00 m3/s以上。
研究区主要分布于新生界第四系地层,有冲积阶地、冲洪积扇地、台地,岩性以褐黄色、暗紫色粘质粉土、粘土和砂卵砾石层为主,地层厚2~120 m。主要土壤类型包括水稻土、黄壤、黄棕壤和紫色土。
1.2 试验设计与样品采集
2022年1月依据地形构建2块面积超过667 m2的试验地,其中667 m2试验地,客土10 cm,所客土壤为周边山地黄壤。2022年2月对滩地左侧河沟进行建议的生态拦截,以保障2022年5月3日汛期发生时将大量泥沙带入试验地实现引洪漫淤效果。2022年5月10日破开生态拦截坝,试验地漫淤泥石厚度为8~14 cm。利用日光晒土1周后播种夏玉米。
2022年8月10日在玉米生长旺盛期调查土壤细菌群落结构。在客土后引洪漫淤(用“TY”代表该处理,T表示客土,Y表示引洪漫淤)和仅引洪漫淤(用“SY”代表该处理,S表示不客土,Y表示引洪漫淤)土地上中随机布设3个1 m×1 m的试验小区,按照5点法采集0~10 cm泥沙样品,按照四分法混合取样100 g,保存在4 ℃车载冰箱中带回,送检测定土壤细菌群落结构。此外,按照相同的布点方法和采样方法采集周边无改造的泥石流河滩地(用“SS”代表该原生生境)的土壤样品作为对照。
1.3 土壤生物16S测序
称取500 mg的样品,放入2 mL无菌离心管中,加入PBS溶液(1X,pH 7.4),震荡混匀,离心3 min后取沉淀物,利用E.Z.N.ATMMag-Bind Soil DNA Kit提取试剂盒(厂商:OMEGA,货号:M5635-02)提取DNA。经电泳质检后利用Qubit3.0 DNA检测试剂盒(厂商:ThermoFisher,货号:Q32854)对基因组DNA精确定量。使用带Barcode的特异引物进行PCR扩增,扩增细菌16S rRNA基因的V3~V4区域。测序引物为16S V3~V4通用引物341F(序列CCTACGGGNGGCWGCAG)和805R(序列GACTA CHVGGGTATCTAATCC)[10]。纯化扩增产物并制备序列文库,然后用Qubit3.0荧光定量仪进行文库浓度测定。最后利用Illumina MiseqTM/HiseqTM平台进行高通量测序。测序下机数据依次按照序列质控、拼接、去除嵌合体、操作分类单元聚类、分类学注释进行预处理,得到OTU数据集[11]。
1.4 统计与分析
为揭示不同处理间土壤OTU丰度的数值分布差异,对OTU的频度计算并绘制密度图。为揭示土壤细菌群落结构组成,在统计分析门水平上细菌结构的基础上,利用随机森林模型[12]预测各处理的关键OTU,并进行重要度排序。为揭示土壤细菌群落的alpha多样性和beta多样性特征[13],本研究基于R的{vegan}函数包计算群落丰度的发现物种数、ACE指数、Chao指数、香浓多样性指数、辛普森指数、香浓-威尔均匀度指数、Goods-coverage指数,以及玉米种植与否、引洪漫淤与否和客土与否对OTU丰度的方差解释率。为揭示关键OTU在表征各处理之间的关系,利用聚类分析和三元分布结构图进行展示。数据分析基于软件R project 4.2.1。
2 结果与分析
2.1 土壤OTU丰度的密度分布情况
由图1可知,各处理OTU丰度的数值分布密度均表现为偏正态分布。其中,无改造的泥石流河滩地(SS)的丰度众数的数量级在千位水平,而仅引洪漫淤(SY)和客土后引洪漫淤(TY)的数量级在百位水平。并且,OTU丰度数值分布的众数与中位数之间的偏离程度表现为SS明显大于TY和SY,表明引洪漫淤后种植玉米对土壤细菌物种丰度的数量级影响较大;而TY和SY的偏离程度十分接近,表明两者土壤细菌物种丰度的数量级差异相对较小。

图1 各处理物种丰度数值分布密度Fig.1 Distribution density of species abundance values of different treatments
2.2 土壤OTU的相对丰度
由图2可知,试验地中,在门水平上物种相对丰度表现为变形菌门(Proteobacteria)最多,相对丰度超过40%。放线菌门(Actinobacteria)的相对丰度在15%左右,拟杆菌门(Bacteroidetes)和酸杆菌门(Acidobacteria)的相对丰度在10%左右,浮霉菌门(Planctomycetes)和蓝菌门(Cyanobacteria)的相对丰度在5%左右,绿弯菌门(Chloroflexi)、芽单胞菌门(Gemmatimonadetes)和疣微菌门(Verrucomicrobia)的相对丰度在1%左右,而其余各门的相对丰度均低于1%。此外,从相对丰度上看,SS、SY和TY之间在菌门之间都存在一定差异。

图2 各处理在门分类水平上的物种差异Fig.2 Species differences at the phylum taxonomic level across the treatments
2.3 土壤OTU的关键物种
由图3-A可知,本研究共计确定1869个OTU。SS、SY和TY土壤中的OTU分别为1219、1362和1115个。其中,SS、SY和TY土壤中共有的OTU数量占总OTU数量的29.75%。SS、SY和TY独有OTU数量分别占总OTU数量的15.57%、8.61%和7.81%,表明SS、SY和TY处理间土壤细菌群落存在一定差异。且这种差异整体表现为组间差异显著大于组内差异(图3-B)。其组间差异表现在以下两方面:一是物种的多样。由表1可知,SS和TY的香浓-威尔多样性指数、辛普森指数和均匀度指数均显著高于SY(P<0.05)。二是关键物种的组成。土壤细菌物种预测种类表现为SY>TY>SS(表1)。利用随机森林预测模型预测OTU对处理的重要性(图3-C)可知,有50个OTU(仅占总OTU的2.68%)对于解释SS、SY和TY的组间差异具有重要意义。其中,对于SS、SY和TY分组的重要性排名前6个OTU分别是OTU0084(变形菌门下的未分类乙型变形菌)、OTU0154(酸杆菌门下尚未明确的难培养Gp1类属细菌)、OTU0007(变形菌门下的交替红色杆菌属细菌)、OTU1426(疣微菌门下未分类疣微菌科细菌)、OTU0660(浮霉菌门下的小梨形菌属细菌)、OTU0214(Candidatus_Saccharibacteria菌门下的Saccharibacteria_genera_incertae_sedis属细菌),其随机森林平均递减准确性值均高于1.70,重要度排名前6(图4-A)。
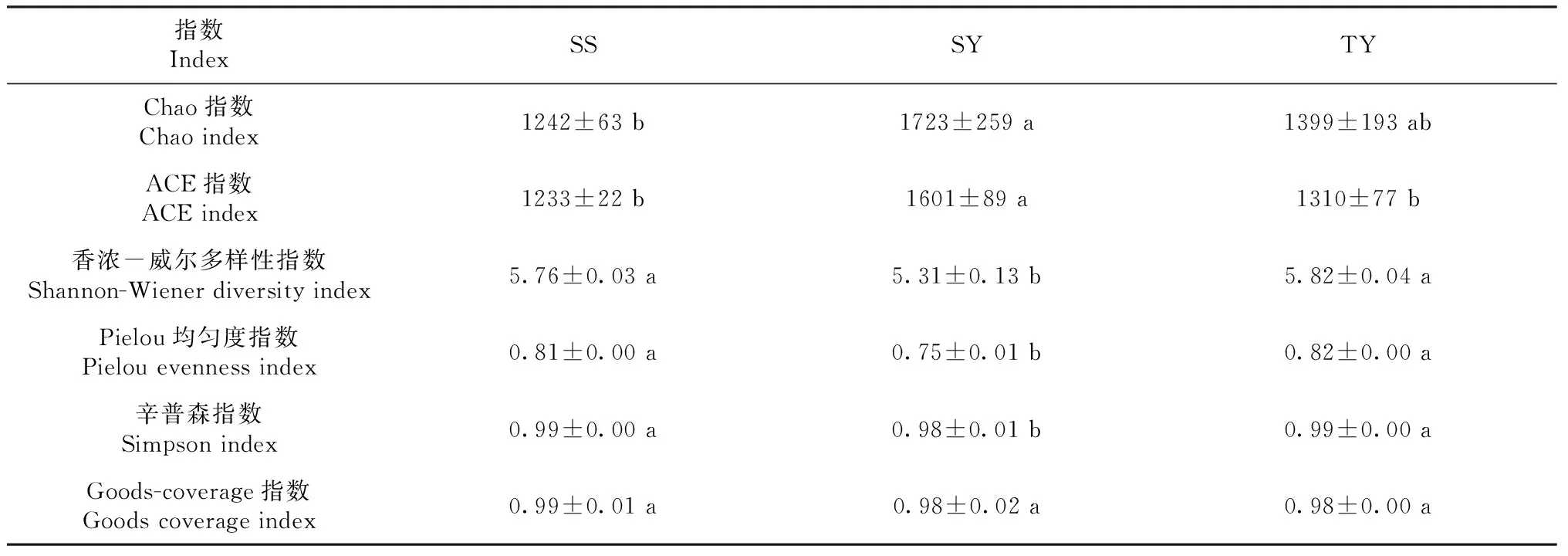
表1 各处理细菌域和真菌域物种分布多样性和均匀性Table 1 Diversity and uniformity of species distribution in bacterial and fungal domains in each treatment

图3 土壤细菌群落结构中的关键OTUFig.3 Key OTU in soil bacterial community structure

OTU0006为叶绿体(Chloroplast);OTU0007为赤杆菌科(Erythrobacteraceae);OTU0022为未归类的酸杆菌GP4(norank Acidobacteria Gp4);OTU0041为未分类的酸微菌亚目(unclassified Acidimicrobidae);OTU0084为未分类的β变形杆菌(unclassified Betaproteobacteria);OTU0087为红杆菌科(Rhodobacteraceae);OTU0097为未归类的噬纤维菌目(norank Cytophagales);OTU0099为草酸杆菌科(Oxalobacteraceae);OTU0109为鞘脂单胞菌科(Sphingomonadaceae);OTU0116为未归类的酸杆菌GP4(norank Acidobacteria Gp4);OTU0122为类诺卡氏菌科(Nocardioidaceae);OTU0127为甲基杆菌科(Methylobacteriaceae);OTU0141为噬几丁质菌科(Chitinophagaceae);OTU0154为未归类的酸杆菌GP1(norank Acidobacteria Gp1);OTU0164为黄单胞菌科(Xanthomonadaceae);OTU0184为丰祐菌科(Opitutaceae);OTU0198为未分类细菌(unclassified Bacteria);OTU0204为嗜甲基菌科(Methylophilaceae);OTU0214为未归类的念珠菌糖杆菌(norank Candidatus Saccharibacteria);OTU0250为未归类的无名候选门WPS-1菌 (norank candidate division WPS-1);OTU0256为未分类的酸微菌科(unclassified Acidimicrobineae);OTU0304为未归类的酸杆菌GP2(norank Acidobacteria Gp2);OTU0313为未分类细菌(unclassified Bacteria);OTU0330为未分类放线菌科(unclassified Actinobacteridae);OTU0350为未分类细菌(unclassified Bacteria);OTU0379为未归类的酸杆菌GP7(norank Acidobacteria Gp7);OTU0395为噬几丁质菌科(Chitinophagaceae);OTU0397为浮霉菌门(Planctomycetaceae);OTU0405为未分类细菌(unclassified Bacteria);OTU0450为黄单胞菌科(Xanthomonadaceae);OTU0471为(Iamiaceae);OTU0570为未归类的酸杆菌GP4(norank Acidobacteria Gp4);OTU0583为未分类蓝藻(unclassified Cyanobacteria);OTU0589为未分类的β变形杆菌(unclassified Betaproteobacteria);OTU0610为黄单胞菌科(Xanthomonadaceae);OTU0660为浮霉菌门(Planctomycetaceae);OTU0708为未归类的酸杆菌GP7(norank Acidobacteria Gp7);OTU0727为厌氧绳菌科(Anaerolineaceae);OTU0747为芽单胞菌科(Gemmatimonadaceae);OTU0748为芽单胞菌科(Gemmatimonadaceae);OTU0759为未分类的β变形杆菌(unclassified Betaproteobacteria);OTU0820为未归类的酸杆菌GP4(norank Acidobacteria Gp4);OTU0852为未归类的酸杆菌GP6(norank Acidobacteria Gp6);OTU0856为丛毛单胞菌科(Comamonadaceae);OTU0880为未归类的酸杆菌GP6(norank Acidobacteria Gp6);OTU0928为黄单胞菌科(Xanthomonadaceae);OTU0977为小单孢菌科(Micromonosporaceae);OTU1145为浮霉菌门(Planctomycetaceae);OTU1426为疣微菌科(Verrucomicrobiaceae);OTU1712为噬几丁质菌科(Chitinophagaceae)。图4 随机森林重要度排名前50的OTU递减准确性值及其聚类Fig.4 OTU decreasing accuracy values for the top 50 random forest significance rankings and their clustering
2.4 关键OTU与处理之间的相关性
由图5可知,TY处理关键OTU的聚集性较高,与SY和SS处理之间区分明显,表明TY处理的关键OTU相对SY和SS处理具有一定的特异性,暗示TY处理重新构建了土壤细菌群落结构。而SS和SY处理之间关键OTU相对分散,表明SY相对SS处理只是改变了原有群落中关键物种的丰度,并没有对群落结构进行重新配置。图4-B中的聚类结果也较好印证了这一结果。此外,图4-B中关键OTU被归为6个类型,而最关键的6个OTU(OTU0154、OTU1426、OTU0214、OTU0660、OTU0084和OTU00 07)则属于4个不同的类型,代表66.67%的OTU类型,具有较高的覆盖度。结合图4~5可知,OTU0154是TY的代表菌,OTU1426和OTU0214是SY的代表菌,OTU0660、OTU0084和OTU0007是SS的代表菌。
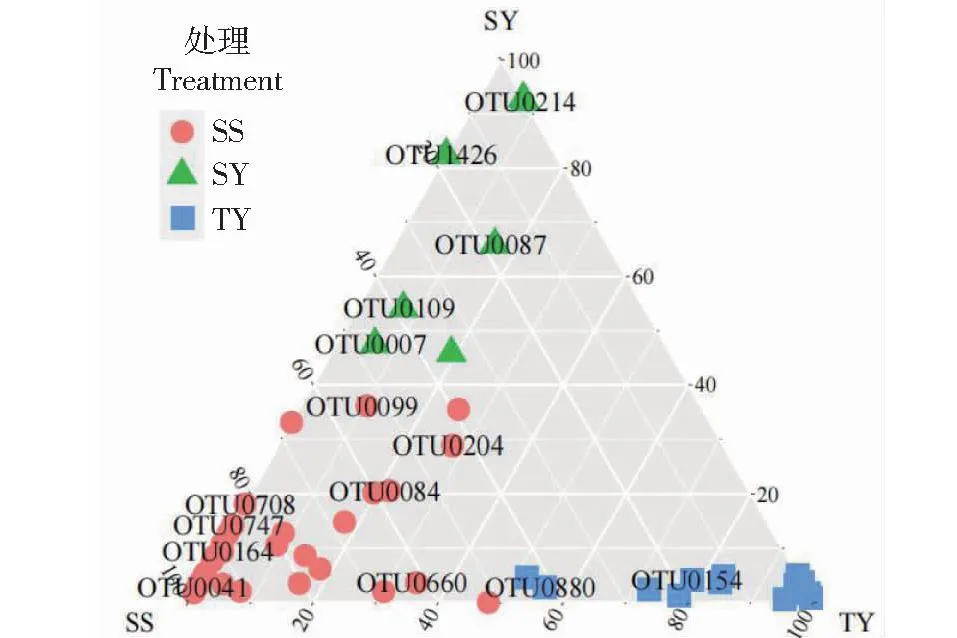
图5 随机森林重要度排名前50的OTU在3个处理上的分布关系Fig.5 Distributional relationship of the top 50 OTU over the three treatments
上述结果表明,本研究的试验处理显著影响了土壤细菌群落结构特征,试验处理可解释87.71%的方差变异(图6)。玉米种植与否、引洪漫淤与否和客土与否分别可解释53.34%、53.92%和53.34%的总方差。其中引洪漫淤与否对于总方差的独立解释比例最大(34.38%),直接效应显著。

图6 方差分解情况Fig.6 The decomposition of variance
3 讨 论
泥石流滩地是山区一种特殊的土地资源,具有地势平坦、灌溉便利、光照充足、热量丰富等优点,生产潜力较高,但砾石含量多、土壤结构不合理、养分贫瘠等不利因素限制其生产潜力的发挥,不加以改造基本上不能作为耕地使用。滩地整治和土地生产力培育的关键是降低砾石含量,改善土壤结构,提高土壤肥力[14]。在泥石流滩地开发利用中,通常先采用人工整地,配合客土、漫灌停淤(引洪漫淤或引浑停淤)等方式增加耕层土壤厚度,改良土壤质地;待滩地开垦出土地后用于种植作物或其他植被,通过人工培肥有效提高土壤养分,土壤物理性质和养分特征的改变与土地利用类型紧密相关[15]。不同改造方法和历时的土壤,其土壤结构、养分含量和生产潜力有所区别。客土对泥石流滩地原始土壤的质地、孔隙度、养分特征等有显著影响,改良效果取决于客土土壤的理化性质及客土比例[16]。漫灌停淤可使滩地增加一定厚度的淤泥层,再通过翻耕,使粗颗粒分布于犁底层之下,细颗粒分布于犁底层之上,经过几次翻耕后,产生质地层次分化,形成上虚下实的“蒙金土”,可提高滩地土壤的保肥蓄水能力。相比于其他开发利用方式,漫灌停淤可显著提高小粒径团聚体的数量,提高土壤养分,土壤有效系数和自然生产潜力高出15%[4,14]。据调查,蒋家沟泥石流滩地经过连续12年的水旱轮作+引洪停淤,滩地土层厚度增加了6倍,土壤机械组成和质地发生明显改变,随着保肥蓄水能力的提升,耕作层全氮含量提高146.6%,有效氮含量提高60.4%,有机质含量提高57%,作物产量大幅提高[2]。漫灌停淤和人工培肥相结合,有效改良泥石流滩地土壤质地、结构和养分,是泥石流滩地的合理开发和可持续利用的重要措施。
土壤微生物群落结构是土壤生态功能的基础,土壤中存在着大量微生物,其中以细菌的种类和数量最多,对土壤营养元素循环、生态环境的改善、植物的生长发育等方面均起着极其重要的作用。大量研究表明,无论是河漫滩、农田、草原、林地还是矿区土壤,变形菌门、放线菌门、酸杆菌门、绿弯菌门和厚壁菌门都是优势类群,但相对丰度相差较大[17-18]。
变形菌门是土壤生态系统中最主要的门类,许多类群与土壤固氮作用相关,能够将有机质分解产生的NH3和CH4等物质用于自身的生长繁殖,并且能够适应大多数复杂的生态环境,广泛存在于各类土壤中[19]。四川省凉山州喜德县红莫镇桃源村泥石流滩地土壤本土细菌变形菌门相对丰度达40%。客土、引洪漫淤后种植玉米对泥石流滩地土壤细菌群落有显著影响,泥石流滩地土壤细菌群落结构改变与其中2.68%的OTU有紧密关联,依据重要性排序,前6个OTU属于变形菌门、酸杆菌门、疣微菌门、浮霉菌门、糖杆菌门,两种改造方式下土壤酸杆菌门、疣微菌门和Candidatus_Saccharibacteria优势明显。酸杆菌门细菌是嗜酸菌,分布较为广泛,可能与该地区土壤pH有关。疣微菌门,兼性好氧,能利用各种糖,在自然界中分布极广,依据共-寡营养理论,疣微菌对资源可利用性采用K选择策略,其数量与土壤营养水平呈显著正相关,说明疣微菌更适合在营养比较丰富的土壤中生存[20-21]。Saccharibacteria是一种无法独立存在的球状细菌,直径只有200~300 nm,需要依赖放线菌门的宿主[22]。在人体和动物的口腔和肠道中,与健康状况有关,在土壤、沉积物和污水中也有存在,具体功能未知[23]。泥石流滩地土壤引洪漫淤结合玉米种植下,土壤细菌群落关键物种丰度改变,结合客土处理可重新配置土壤细菌群落,对土壤性质和养分变化有重要影响。
4 结 论
土壤改造后种植玉米对泥石流滩地土壤细菌群落结构有显著影响,引洪漫淤后种植玉米可促进土壤细菌物种数增加,改变土壤细菌关键物种的丰度;客土后引洪漫淤种植玉米可重新配置土壤群落结构,使土壤细菌物种数显著降低。泥石流滩地土壤细菌群落结构改变与其中2.68%的OTU有紧密关联,依据重要性排序,前6个OTU属于变形菌门(2个)、酸杆菌门、疣微菌门、浮霉菌门、Candidatus_Saccharibacteria。
猜你喜欢
中国农村水利水电(2023年6期)2023-07-04
热带作物学报(2022年8期)2022-09-16
中国比较医学杂志(2020年4期)2020-05-26
水生生物学报(2019年4期)2019-07-20
生物安全学报(2019年3期)2019-02-15
川北医学院学报(2019年6期)2019-02-10
中国农村水利水电(2018年12期)2018-12-28
中国水土保持科学(2016年4期)2016-09-21
建筑工程技术与设计(2015年20期)2015-10-21
铁道标准设计(2014年1期)2014-01-03
