
卷丹百合LlARR1及其在珠芽形成过程中的表达分析
2024-01-01 01:33陈妍竹张雪敏俞诗音王梦迪杜运鹏杨凤萍张秀海李玉舒曹丽
河南农业大学学报 2023年6期
陈妍竹,张雪敏,俞诗音,王梦迪,杜运鹏,杨凤萍,张秀海,李玉舒,曹丽
(1.延边大学农学院,吉林 延吉 133002; 2.北京市农林科学院草业花卉与景观生态研究所,北京 100097;3.北京林业大学园林学院,北京 100083; 4.北京农业职业学院,北京 102442)
百合是百合科(Liliaceae)百合属(Lilium)植物,属野百合的一个变种,具备极高的观赏价值[1]。百合属植物有110~115个种,其中只有卷丹百合(Liliumlancifolium)、淡黄花百合(Liliumsulphureum)、通江百合(Liliumsargentiae)和珠芽百合(Liliumbulbiferum)这4个种在自然条件下可在茎的叶腋处形成珠芽。珠芽繁殖具有繁殖系数高、低毒,生产的种球质量高,育种周期短等优势,已成为卷丹重要的繁殖策略之一[2]。在有珠芽百合中,卷丹表现出更多的特殊性。卷丹是在中国分布范围最广的百合科百合属植物,分布于中国17个省、自治区、直辖市。其花大色艳,抗性强,兼具观赏、食用、药用价值,为中国食用百合的主要野生种[3]。截至2019年,中国食药用百合产业规模超过100亿元,而卷丹是中国药典记载的三大药用百合之一,种植面积约30 km2,产值约45亿元,是食药用百合种植面积最多的种。卷丹一般为三倍体,难以通过有性繁殖方式扩大种群。目前,制约中国食药用百合产业发展的主要阻碍之一是百合种球退化严重。因此脱毒种球的生产是百合产业提质增效、绿色可持续发展的核心。珠芽发生机制的解析,对提高百合的栽培及繁殖效率有着重要的生产意义,对促进百合产业发展具有重要理论和应用价值。
细胞分裂素作为重要的植物激素,几乎参与植物生长发育的各个方面,如顶端优势、延迟衰老、顶端分生组织维持、从头器官再生、根系增殖、顶端分生组织功能和营养信号转导等[4-5]。研究表明,细胞分裂素参与珠芽的形成,并在珠芽的形成过程中起促进作用[6]。在薯蓣(Dioscoreapolystachya)体外球茎诱导过程中,增加培养基中细胞分裂素的浓度可以显著提高球茎诱导率[7]。NAVARRO等[8]报道,外施细胞分裂素能够促进番茄(SolanumlycopersicumL.)腋下块茎的形成。反应调节因子家族(response regulators,RRs)被认为是植物细胞分裂素响应的主要调节因子,对拟南芥基因组数据进行鉴别分析,发现该家族共有24个基因,被分为了3类:A型、B型和C型[9]。在结构上A型只带有可接受磷酸的接收器结构域,由细胞分裂素转录诱导,是主要的细胞分裂素应答基因,负调控细胞分裂素信号通路,降低对细胞分裂素的敏感性[10-14]。C型ARRs与A型ARRs相似,但它们的表达不是由细胞分裂素诱导的[9,15]。B型ARRs带有保守的核定位信号(nuclear localization signal, NLS)区域,在亚细胞定位研究中全部定位于细胞核[15]。而B型除具有接收器结构域之外,还包含与DNA结合的MYB-like结构域与反式激活区域,是细胞分裂素信号通路中的正调控转录因子,可直接与靶DNA序列结合,激活靶基因的表达[16-17]。ARR1属于B型ARR基因,是最先被认为是转录因子的RRs家族成员之一。研究表明,ARR1在细胞分裂素信号传递的通路中起着核心的调控作用[16]。且干细胞分化和腋芽的形成与细胞分裂素信号通路通过B-ARRs激活WUSCHEL基因的表达相关[18-19]。
通过北京市农林科学院草叶花卉与景观生态研究所百合组前期珠芽形成过程的转录组数据发现,LlARR1基因在卷丹S0(珠芽准备发生期)和S1(珠芽小白点时期)[6]的表达中具有显著差异。对LlARR1基因在卷丹百合珠芽形成过程中表达分析进行研究,以期能够进一步阐述卷丹珠芽发生的性状,为卷丹珠芽发生机制研究提供理论支持。
1 材料与方法
1.1 材料与试剂
1.1.1 植物材料 卷丹独头种球及不同品种有珠芽、无珠芽百合种植于北京市农林科学院国家百合种质资源圃中。根据北京市农林科学院草叶花卉与景观生态研究所百合组前期对珠芽的观察,将其发生发育阶段划分了4个时期分别为S0、S1、S2、S3[6]。每个时期取9株卷丹,每株卷丹取同一部位的2个相邻叶腋,每3株卷丹的叶腋作为一个混合样本混样;同时取每株卷丹百合上下部叶腋处;不同品种百合取其植株上部叶腋处混合样本锡箔纸包裹后用液氮速冻,在-80 ℃冰箱中冻存,用于后续总RNA的提取及qRT-PCR的检测。
1.1.2 不同激素处理卷丹百合 随机选取30株卷丹百合进行外源激素处理并观察。喷洒蒸馏水作为对照,且每3 d喷洒100 mg ·L-1生长素IAA和100 mg·L-1生长素转运抑制剂NPA以及100 mg·L-1赤霉素GA3溶液。每个处理中均使用了10个独立的植株。
1.1.3 菌株、载体和试剂 EASYspin Plus多糖多酚植物RNA快速提取试剂盒(爱博森生物科技有限公司,北京),大肠杆菌(DH5α)感受态BC102细胞(博迈德生物技术有限公司,北京),pCE2 TA/Blunt-Zero Vector载体5 min TM TA/Blunt-Zero Cloning Kit,2×Phanta® Flash Master Mix(Dye Plus),2×Rapid Taq Master Mix和反转录试剂HiScriptⅢ SuperMix for qPCR(+gDNA wiper)(诺唯赞,南京),DNA通用纯化回收试剂盒(天根生化科技有限公司,北京),荧光定量PCR试剂TB Green® Premix ExTaqTM(宝生物工程有限公司,大连)。
1.2 试验方法
1.2.1 卷丹百合中LlARR1来源 通过对处于不同发育时期的卷丹叶腋和珠芽混样进行转录组测序分析,并基于前人[20]对细胞分裂素通路研究的基础上,在转录组测序数据中筛选细胞分裂素信号途径相关所有基因。
1.2.2LlARR1基因克隆及定量引物设计 利用卷丹珠芽发生发育过程转录组数据筛选获得目的基因,利用目的基因的转录本序列在ORF Finder(https://www.ncbi.nlm.nih.gov/orffinder/)获得其开放阅读框(open reading frames, ORF)序列,将获得的长度大于200 bp的ORF序列在NCBI protein BLAST(https://blast.ncbi.nlm.nih.gov/-Blast.cgi)依次进行比对,与转录组注释结果相吻合即视为比对成功,保存比对成功的ORF序列用于定量引物设计。将保存的ORF序列提交至GenScript(https://www.genscript.com/tools/real-time-pcr-taqman-primer-design-tool?page_no= 1& position_no=2&sensors=googlesearch),调整参数orga-nism:other,size range:180~220 bp,Tm≥60 ℃进行定量引物设计,根据引物设计的原则选择最好的1~2个设计结果作为定量引物,后续再进行定量引物的筛选。
克隆引物和病毒检测引物的设计则是将该ORF序列提交至NCBI(https://www.ncbi.nlm.nih.gov/)查找获得基因的CDS序列,利用Primer Premier 5软件设计克隆引物,荧光定量引物,克隆引物和病毒检测引物均由生工生物工程(上海)股份有限公司合成(表1)。
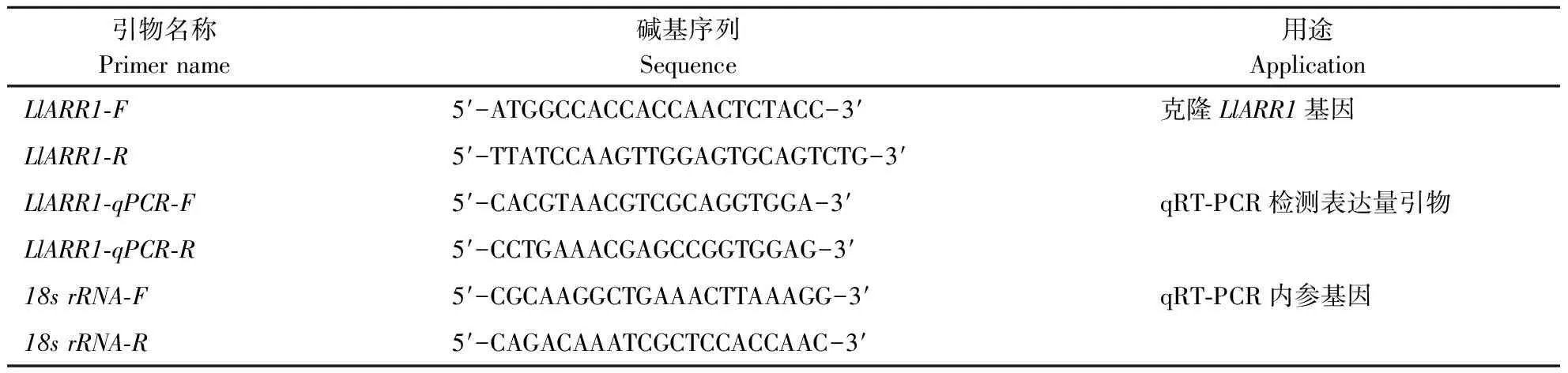
表1 LlARR1基因克隆及定量试验所用引物Table 1 Primers used in this study
1.2.3 总RNA的提取及逆转录 试验中选取的材料包括卷丹百合叶腋、卷丹百合不同组织、不同品种百合(有珠芽和无珠芽)开花前的茎秆上部叶腋以及不同激素处理(IAA、NPA、GA3,喷施质量浓度均为100 mg·L-1)的卷丹百合叶腋,其总RNA利用EASYspin Plus多糖多酚植物RNA快速提取试剂盒提取;根据反转录试剂HiScript Ⅲ SuperMix for qPCR(+gDNA wiper)将提取的RNA反转录合成cDNA,反应体系为:5×HiScriptIII qRT SuperMix 4 μL,4×gDNA wiper Mix 4 μL,RNA 1 μg,RNase Free ddH2O添加至20 μL;反应条件为:37 ℃ 15 min;85 ℃ 5 s;4 ℃永久保存。反应结束后,样品于-20 ℃冰箱保存备用。
1.2.4 转录组测序样品制备及文库构建 卷丹百合S0、S1、S2、S3,4个时期叶腋样品,液氮冷冻,-80 ℃冻存。由北京诺禾致源科技股份有限公司进行转录组测序及初步数据分析。NCBI SRA (顺序读取存档)登录号为PRJNA916842。
采用EASYspin Plus多糖多酚植物RNA快速提取试剂盒(爱博森生物科技有限公司,北京)提取总RNA。首先通过Oligo(dT)磁珠富集带的mRNA,随后加入Fragmentation Buffer随机打断得到的mRNA。以片段化的mRNA为模板,在M-MuLV逆转录酶体系中合成cDNA第一条链,随后用RNaseH降解RNA链,并在DNA polymerase I体系下,以dNTPs为原料合成cDNA第二条链。纯化后的双链cDNA经过末端修复,加A尾并连接测序接头,用AMPure XP beads筛选约370~420 bp的cDNA,进行PCR扩增并再次使用AMPure XP beads纯化PCR产物,最终获得文库。文库构建完成后,首先使用Qubit 2.0 Fluorometer进行初步定量,稀释文库至1.5 ng·μl-1,随后使用Agilent 2100 bioanalyzer对文库大小进行检测,而后,qRT-PCR对文库有效浓度进行准确定量(文库有效浓度高于1.5 nmol·L-1),以保证文库质量。
1.2.5LlARR1基因克隆 利用卷丹叶腋cDNA为模板进行PCR扩增,扩增体系包含2×Phanta Master Mix 25 μL,上、下游引物各1 μL,cDNA 2 μL,ddH2O 19 μL,总体系50 μL。反应程序为98 ℃ 30 s,98 ℃ 10 s,55 ℃ 5 s,72 ℃ 15 s,72 ℃ 5 min,共35个循环。扩增后的PCR产物用1.5%琼脂糖凝胶电泳检测,并用DNA回收试剂盒(TIANGEN,北京)回收目的片段;将目的片段连接至pCE2 TA/Blunt-Zero Vector载体,并转化到DH5α(博迈德,北京)感受态中,涂布于LB固体培养基(含50 mg·L-1卡那霉素),37 ℃培养12 h后,随机挑取单克隆并进行菌液PCR检测,将条带位置正确的阳性单克隆菌液送往生工生物工程(上海)有限公司。
1.2.6LlARR1序列分析、蛋白结构预测及系统进化树构建 通过ProtParam对蛋白理化性质进行分析;利用ProtScale对蛋白亲疏水性进行分析;采用TMHMM Serverv 2.0分析编码氨基酸的跨膜结构域;利用NetPhos 2.0 Server进行磷酸化位点预测;利用NCBI ORF finder在线程序及Conserved domains数据库对测序获得的cDNA序列进行开放阅读框及保守功能结构域分析;利用PredictProte预测蛋白一级结构域;通过SOPMA及SWISS.MODEL预测蛋白的二级和三级结构;利用Cell-PLoc 2.0分析亚细胞定位情况;利用JAlview进行编码蛋白多重序列比对;利用MEGA7.0进行Neighbor.joining系统进化树构建。系统发育树中序列的模体结构采用MEME(https://meme-suite.org/me-me/tools/meme)在线网站进行分析。
1.2.7LlARR1的表达分析 实时荧光定量PCR反应体系:5 μL TB green、1 μL cDNA模板、0.4 μL上下游引物、3.6 μL ddH2O。扩增程序:95 ℃ 30 s,95 ℃ 5 s,55 ℃ 15 s,72 ℃ 30 s,95 ℃ 5 s,循环39次。以卷丹18S为内参基因,采用2-ΔΔCt法计算LlARR1的相对表达量。利用Excel 2019、SPSS17.0、GraphPad Prism8等软件进行统计学分析。
2 结果与分析
2.1 卷丹百合珠芽发生发育时期划分
根据卷丹百合珠芽发生发育过程将其划分为4个时期,将第1个叶腋处产生小白点时期记为S1时期,将该叶腋上部1~3个叶片的叶腋记为S0时期,即珠芽准备发生时期。随着珠芽发育,依次将珠芽绿球期记为S2时期,珠芽成熟期记为S3时期(图1)。

A. S0:珠芽准备发生期;B. S1:珠芽小白点期;C. S2:珠芽绿球期;D. S3:珠芽成熟期。A.S0: Preparation stage; B. S1: White dot stage; C. S2: Green bulbil stage; D. S3: Mature bulbil stage.
2.2 卷丹ARR1在珠芽发生过程转录组中的表达
基于细胞分裂素在腋芽发生过程中的重要性[6],为探究卷丹百合珠芽发生过程中的细胞分裂素相关的关键转录因子,对S0和S1时期的转录组数据进行分析,在细胞分裂素信号通路中,筛选到了一个注释为ARR1的差异表达基因。相比于S1时期,该基因在S0时期有较高的表达(图2-A),并且荧光定量的结果与转录组数据一致(图2-B)。

**表示差异极显著(p<0.01)。下同。 ** indicates a significant difference (p<0.01). The same as below.
2.3 卷丹LlARR1基因CDS全长克隆及其编码氨基酸序列分析
为获得LlARR1基因CDS序列,将其转录本序列提交至ORF Finder(https://www.ncbi.nlm.nih.gov/orffinder/)找到一条全长2 148 bp的序列,使用引物LlARR1-F和LlARR1-R对卷丹LlARR1基因进行PCR扩增,获得1条长度约为2 000 bp的条带(图3-A),纯化回收后连接TA/Blunt-Zero Vector载体转化大肠杆菌后,通过菌液PCR挑选条带正确的阳性克隆单克隆菌液送测序。测序结果表明,LlARR1基因开放阅读框(ORF)长2 148 bp,共编码715个氨基酸。如图3-B中,红色方框为起始密码子,蓝色方框为终止密码子。
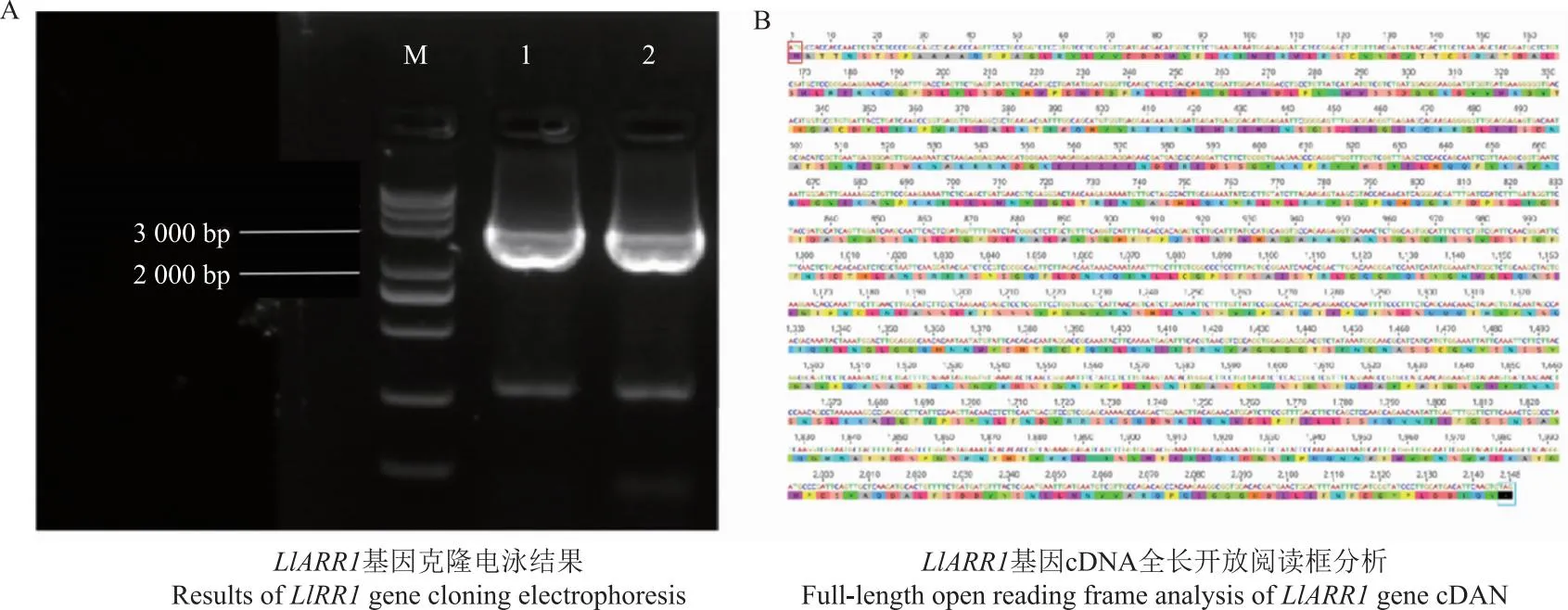
注:M代表DNA marker trans2K plus; 1,2指LIARR1基因的PCR产物。Note:M represents for DNA marker trans2K plus,1,2 refers to the PCR product of the LIARR1 gene.
2.4 LlARR1基因系统进化树构建
经分析,16种ARR1蛋白聚为5个小分支,具有较高的支持率,LlARR1与卷丹(Liliumlancifolium)、杂交百合(Liliumhybrid division)的ARR1序列处于同一个进化分支上,表明它们的亲缘关系最近(图4)。模体结构分析显示,基序1,2,3比较保守,不同物种的16个ARR1蛋白均含有1,2,3基序,且均无特有基序,说明ARR1蛋白是比较保守的。此外,聚合在系统进化树同一分支内的不同植物ARR1的模体基序是相似的,关系越近,模体基序越相似。
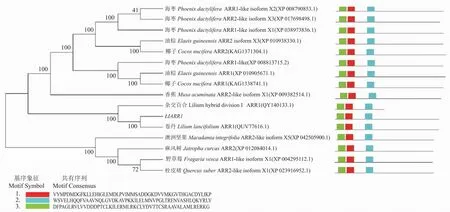
图4 LlARR1与其他物种同源蛋白的系统进化树及模体分析Fig.4 LlARR1 Phylogenetic tree and motif analysis of proteins homologous to other species
2.5 卷丹LlARR1蛋白生物信息学分析
2.5.1 卷丹LlARR1蛋白的理化性质分析 对卷丹LlARR1基因编码的蛋白质的理化性质进行预测分析,结果显示,该蛋白分子式为C3361H5329N987O1102S29,相对分子质量为78 125.98,理论等电点为5.76,脂肪系数为72.24。LlARR1蛋白的氨基酸中共含有82个带负电荷氨基酸残基(Asp+Glu),70个带正电荷氨基酸残基(Arg+Lys)(图5-A)。不稳定系数为50.68,蛋白亲水性平均值为-0.517,因此LlARR1为不稳定疏水性蛋白(图5-B)。LlARR1蛋白从内到外无跨膜区域,有接近1的概率表明该蛋白位于膜外,推测是一种非跨膜蛋白(图5-C)。LlARR1蛋白含有可能发生磷酸化的位点有81个,包括Ser(丝氨酸)60个、Thr(苏氨酸)16个、Tyr(酪氨酸)5个(图5-D)。
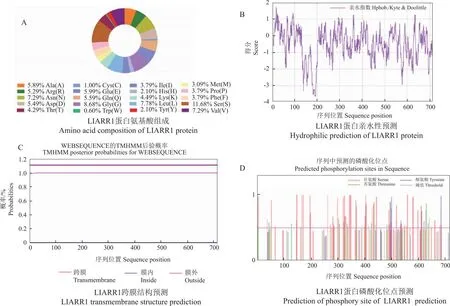
图5 LlARR1蛋白理化性质分析Fig.5 Analysis of physicochemical properties of LlARR1 protein
2.5.2 卷丹LlARR1蛋白质二、三级结构的预测分析 采用NCBI CD search在线网站分析LlARR1基因编码蛋白质的结构域,该蛋白有2个specific-hits,为REC_typeB_ARR-like和myb SHAQKYF位点,分别属于REC superfamily、SANT superfamily(图6-A)。
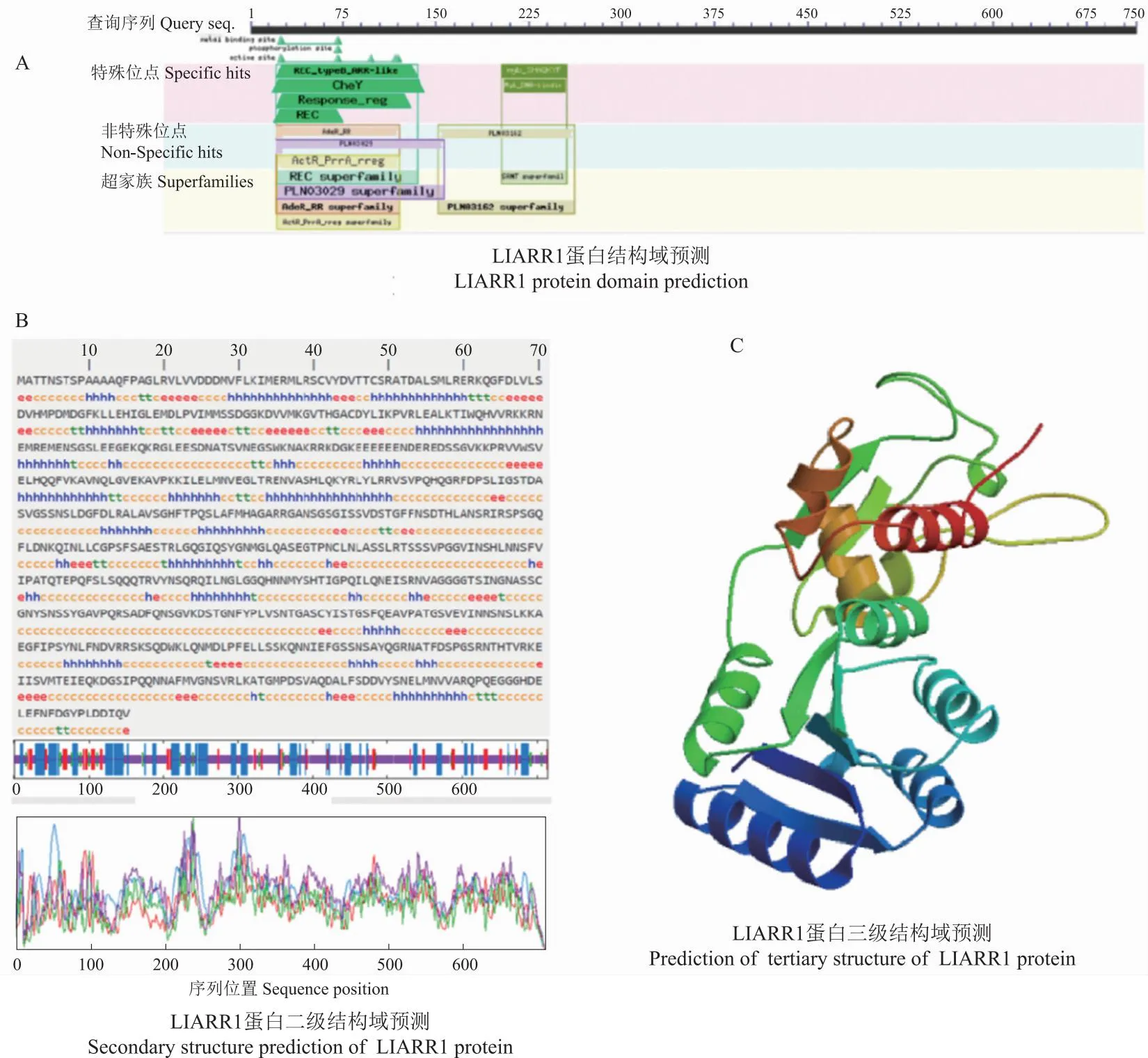
图6 LlARR1蛋白质二三级结构预测Fig.6 Prediction of secondary and tertiary structure of LlARR1 protein
注:不同字母表示在0.05水平上差异显著。下同。Note: The different letters within the same column mean significant difference by Duncan’s multiple range test at p<0.05.The same as below.
利用SOPMA在线软件对蛋白质的二级结构进行预测和分析。结果表明,LlARR1蛋白的二级结构的含量中:无规则卷曲(黄色c)(58.32%)>α螺旋(蓝色h)(26.01%)>延伸链(红色e)(10.63%)>β转角(绿色t)(5.03%),因此可推断,无规则卷曲和α螺旋是LlARR1蛋白的主要组成成分(图6-B)。蛋白质的多肽链在二级结构的基础上进行盘曲或折叠形成具有规律的三维空间结构。利用在线软件SWISS-MODEL对蛋白质三级结构进行预测分析,获得了LlARR1蛋白的三维结构模型,发现LlARR1以DNA-binding response regulator作为目标建模,含有8个α螺旋,6个β折叠 ,组分间均以β转角(loop)相连(图6-C)。采用CellPLoc 2.0亚细胞定位显示,该蛋白主要定位于细胞核。
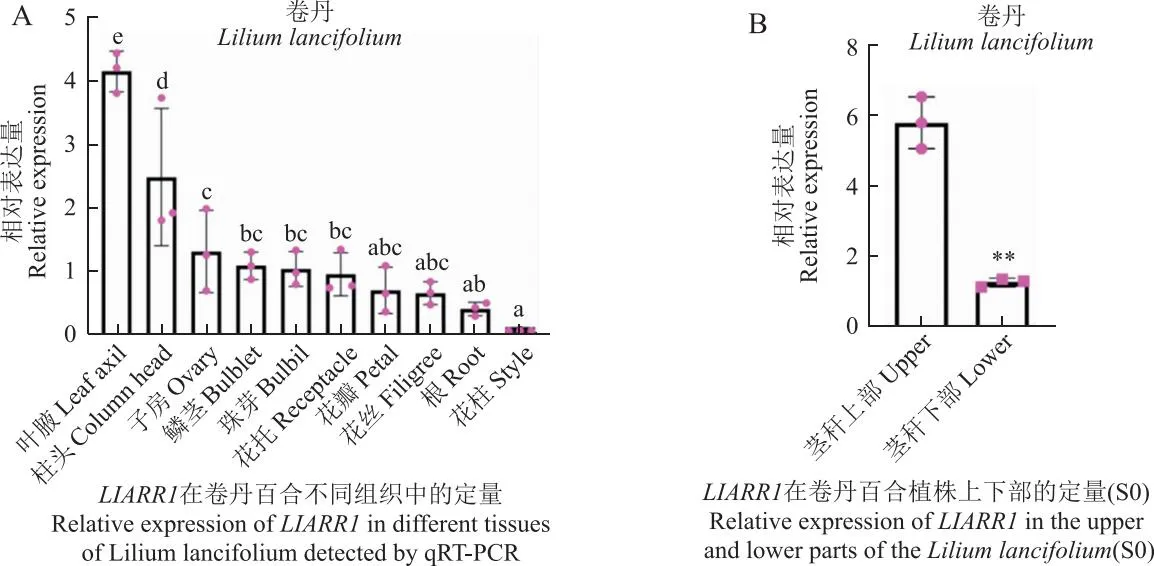
2.6 LlARR1在卷丹百合不同组织及植株上下茎部的表达分析
为探究LlARR1的表达模式,通过qRT-PCR检测了卷丹百合不同组织及同一植株S0时期茎秆上下部中LlARR1的相对表达量。从图中可以看到,LlARR1在叶腋中的表达最高,在花柱中的表达水平最低,并且其在珠芽中的表达水平显著低于叶腋(图7-A),推测LlARR1的表达主要集中于珠芽准备发生时期,即S0时期,但在珠芽发育时期的表达处于较低水平。随后发现,LlARR1在茎秆上部的表达水平显著高于茎秆下部,而卷丹珠芽的发生主要集中于茎秆上部(图7-B)。因此,基于以上结果可以说明,LlARR1可能主要在珠芽发生过程中发挥作用。
2.7 ARR1在不同百合品种中的定量分析
为进一步探究ARR1在珠芽发生中表达模式,选取了不同系列的11种百合来检测茎秆上部ARR1的表达量,分别是A系列品种(‘Flore Pleno’‘Pearl White’‘Red Velvet’)、A复色品种(‘Fore-ver Linda)、Free A系列(‘Easy Whisper’)、Tiger A系列(‘Strawberry Event’)、LA重瓣系列(‘Must See’)、OA系列(‘Hotel California’)、O系列(‘Brasilia’)、OT系列(‘Red Morning’)和T系列(‘Orange Planet’)。以上11种百合中,OA系列的‘Hotel California’和A系列的‘Flora pleno’均可以自然产生珠芽,‘Must See’在植株开花后才能产生珠芽外,而其余8个品种在自然条件下均不能产生珠芽。检测发现,ARR1在2种可以产生珠芽的百合中的表达水平显著高于其他9个品种(图8)。而图9中ARR1在‘Red Velvet’、Strawberry Event’2个不产生珠芽品的相对表达量高于‘Must See’。这可能是由于取样时‘Must See’尚未产生珠芽。该结果进一步说明ARR1可能作用于珠芽的发生过程,并可能发挥一个正调控因子的作用。
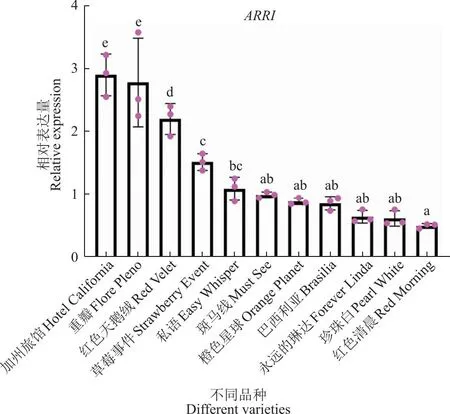
图8 ARR1在不同百合品种中的表达水平Fig.8 Expression levels of ARR1 in different varieties of lily
注:以NPA作为相对的量;*表示差异显著(p<0.05);**表示差异极显著(p<0.01):ns表示无显著差异(p>0.05)。Note: NPA is used as a relative quantity;* indicats significant difference (p<0.05); ** means highly significant difference (p<0.01); ns means no significant difference (p>0.05).
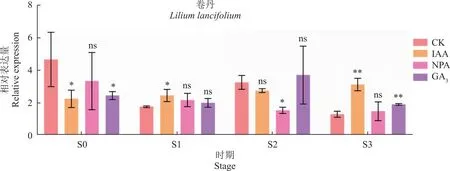
2.8 LlARR1在不同激素处理中的表达分析
珠芽的发生发育受到多种激素的调控[2]。为探究LlARR1对不同激素的响应,对卷丹植株进行了3种激素处理(IAA、NPA、GA3)(图9)。结果发现,对照植株中,LlARR1在S2时期有一个小峰值,推测其在珠芽发育过程中也发挥作用。IAA处理中,LlARR1的表达在S0时期被显著抑制,但在后期的S3时期被显著上调。NPA的处理缓解了S0时期IAA对LlARR1的抑制,但在后期的S2时期,NPA对LlARR1有一定的抑制作用。在赤霉素处理中,LlARR1在S0时期被显著抑制,但在后期的发育过程中均被上调。由该结果推测,LlARR1可能在珠芽准备期发挥的作用与生长素和赤霉素相反,但在后期的珠芽发育过程中发挥相似的作用。
3 结论与讨论
拟南芥中的细胞分裂素信号通路涉及3个组成部分:拟南芥组氨酸激酶(AHKs)、拟南芥组氨酸磷酸转移酶(AHPs)和拟南芥反应调节因子(ARRs)[21-22,17]。拟南芥中的细胞分裂素受体家族由3个组氨酸激酶组成:AHK2、AHK3和AHK4(CRE1或WOL1)[23-24]。AHPs是AHK细胞分裂素受体的下游靶蛋白,并将磷酸基转移到下游的ARRs,B型ARRs是最终的磷酸化受体[25-28]。已有研究表明,ARR1可能是细胞分裂素通路中参与调控珠芽发生的相关基因。YANG等[29]通过转录组分析发现,细胞分裂素的高生物合成和低降解可能促进卷丹上部叶腋处珠芽的形成,HE等[30]的研究发现,细胞分裂素可以促进珠芽的形成,尤其是珠芽的启动,而ARR1是细胞分裂素信号通路中重要的转录因子[31],推测卷丹珠芽形成过程中ARR1基因可能发挥重要作用。而其研究检测了ARR1在卷丹珠芽发生过程中的表达,发现在珠芽发生过程中ARR1的表达表现出先升高后下降的趋势,这与本研究结果有所不同,可能是由于对发生时期的划分有所不同所导致,但这些结果均说明ARR1可能参与卷丹珠芽的发生。在拟南芥的腋芽发生过程中,外源施加细胞分裂素,可以促进ARR1的表达进而激活WUS的表达来促进腋芽的再发生,这可以说明细胞分裂素在腋芽的发生过程中起着重要的作用[18]。目前,对ARR1在卷丹珠芽发生发育过程中的表达模式及其结构性质进行具体的探讨较少,且缺少ARR1参与珠芽发育过程的研究,本研究初步划分了卷丹珠芽的发生发育时期,并对LlARR1在卷丹发生发育时期的表达模式进行了检测。
本研究通过对S0和S1时期卷丹珠芽转录组测序分析,从差异基因中筛选得到了差异显著的基因ARR1,并进行生物信息学分析。氨基酸多序列同源比对发现LlARR1和其他植物的同源性较高,表明植物中ARR1蛋白的氨基酸序列较为保守,在模体分析中这一点也得到了验证。模体结构分析发现,不同植物ARR1蛋白基序大多比较保守。此外,系统进化树及模体结构分析还发现,亲缘关系越近的种属,ARR1蛋白的模体结构越相似,LlARR1与卷丹ARR1的模体结构一致,说明二者确为同一基因,与杂交百合ARR1相似度最高,推测二者可能具有相似的功能。且LlARR1编码一个由715个氨基酸残基组成的蛋白序列,为疏水性不稳定蛋白,无跨膜区,有大量磷酸化作用位点,理论等电点为5.76。亚细胞定位预测结果发现,LlARR1定位于细胞核中。蛋白结构预测分析发现,LlARR1以DNA-binding response regulator作为目标建模,无规则卷曲和α螺旋是LlARR1蛋白的主要组成成分。
LlARR1的表达模式分析发现,相对于S1时期,LlARR1珠芽S0时期有较高的表达水平(图2),主要集中在叶腋处表达,并且其在茎秆上部叶腋处S0时期的表达远高于茎秆下部的表达(图7),推测LlARR1在珠芽启动过程中发挥正调控因子的作用。该推测被LlARR1在不同百合品种中的表达结果进一步验证,即LlARR1在自然发生珠芽的百合品种叶腋处有较高的表达水平。在不同激素处理的卷丹百合中,LlARR1的表达在S0时期均被生长素和赤霉素显著抑制,由该结果推测,LlARR1可能在珠芽准备期发挥的作用与生长素和赤霉素相反。而生长素处理下的卷丹百合在S1时期与对照植株相比有显著上调,由此可以推测LlARR1可能在S1时期协同生长素并促进卷丹珠芽的形成,这与YANG等[29]的结果相一致(图9);且谭长龙等[32]的研究也表明,较低含量的生长素有利于珠芽魔芋侧叶萌发。同时在之后的S3时期进一步与生长素有协同,促进珠芽的发育,但外源喷施生长素是否能增加珠芽的发生还有待探究。而在赤霉素处理中,LlARR1在后期的发育过程中均被上调,由此推测在后期的珠芽发育过程中其与赤霉素可能协同发挥作用。龚明霞等[33]对山药进行外源喷施赤霉素,发现其可能抑制珠芽形成,而在本文研究中还不能证明赤霉素的外源施加对卷丹百合珠芽形成起到正调控或负调控作用,同时赤霉素是否能够促进卷丹珠芽的发生发育还有待探究。
本研究从卷丹叶腋中克隆到细胞分裂素传递通路中的核心转录因子LlARR1,全长2 148 bp,编码715个氨基酸,为不稳定疏水性蛋白,与杂交百合ARR1亲缘关系最近,且在不同植物物种中ARR1氨基酸较为保守。对其表达模式分析表明,LlARR1可能参与了卷丹珠芽的发生和发育,且在珠芽发生发育过程中可能发挥促进作用。但后期仍需对LlARR1在卷丹珠芽发生及发育过程中的功能进行验证,从而了解细胞分裂素在珠芽形成中的作用,以期为珠芽发生调控及其分子机制的研究提供理论依据。
猜你喜欢
种子(2023年8期)2023-11-15
贵州农业科学(2023年10期)2023-10-27
江西农业学报(2022年3期)2022-05-24
草地学报(2021年6期)2021-07-05
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
特种经济动植物(2021年2期)2021-02-25
中学生物学(2016年8期)2016-01-18
海燕(2015年1期)2015-11-18
园艺与种苗(2014年1期)2014-03-06
食品科学(2013年17期)2013-03-11
