
小叶青冈幼林生长和形质性状家系变异与选择*
2024-01-05 11:34徐永宏杨孟晴代丽华姚甲宝周志春王斌
西部林业科学 2023年6期
徐永宏,杨孟晴,代丽华,姚甲宝,周志春,王斌
(1.浙江省建德市林业总场,浙江 建德 311600;2.中国林业科学研究院亚热带林业研究所/浙江省林木育种技术研究重点实验室,浙江 杭州 311400;3.江西省林业科技实验中心,江西 信丰 341600;4.中国林业科学研究院亚热带林业实验中心,江西 分宜 336600)
林木性状的变异受一定程度的遗传控制,变异的丰富程度决定了遗传改良的潜力,而揭示林木性状遗传变异规律有助于制定科学的育种策略[1]。在林木遗传改良中,多基于优良种源选择的基础上,进一步开展优良家系和优良个体选择[2-3]。林木生长周期长,开展家系早期生长和形质性状变异研究和优良家系早期筛选,可有效缩短育种周期,加速育种进程[4-5]。通常变异丰富的林木对环境的适应能力较强[6-7]。通过多点区域试验,可选择遗传性状稳定的优良家系用于造林,也可分别不同立地和地区选择适生的优良家系。不同学者基于多点区域试验开展木荷(Schimasuperba)[8]、马尾松(Pinusmassoniana)[9]和南方红豆杉(Taxuswallichianavar.mairei)[10]优良家系选择均取得较好效果。根据遗传测定选出的优良家系或单株,不仅可供生产直接应用,而且可作为下一个世代的育种材料[11]。
小叶青冈(Cyclobalanopsismyrsinifolia)为青冈属常绿大乔木,是我国亚热带地带性常绿阔叶树种,常生于海拔200~2 500 m的山谷和阴坡等的杂木林中,为青冈(C.glauca)在海拔较高地区垂直分布的替代种,水平分布于浙江、江西和福建等省天然林中且资源较为丰富。小叶青冈树体高大、干形通直、木材坚硬耐腐耐磨,是优良的工业和民用木材之一,其生长较快、适应性强,是我国南方优先发展的珍贵用材树种和生态修复树种之一。现在各地虽有有一定规模的人工林,栽培效果也较好,但缺少应用良种营造的人工林,对其种内遗传变异研究也较少。徐肇友等[12]基于赤皮青冈(C.gilva)、小叶青冈和福建青冈(C.chungii)等3种青冈家系子代的早期测定,发现不同青冈及其家系间生长性状存在丰富的变异,遗传改良潜力较大。本文以建立在浙江建德、江西安远和江西分宜3个试验点的3 a生小叶青冈优树家系测定林为研究对象,分析早期生长和形质性状的家系遗传变异规律,以及家系与地点互作效应等,用以指导小叶青冈遗传改良方案的制定,并初选一批早期速生、形质良好和适应性强的优良家系,以期为其速生优良家系的推广应用提供参考。
1 材料与方法
1.1 试验材料
2017年11月,在小叶青冈天然林资源丰富的江西上饶和分宜、浙江庆元、福建建瓯等地开展小叶青冈优树选择。在以小叶青冈为主的优良天然林分中,沿一定的线路调查,将树形高大、干形通直圆满、枝叶色泽正常、长势良好无病虫害等符合要求的单株作为候选树。采用优势木对比法进行优树选择,所选优树的间距在50 m以上。在优树选择的同时,采集其种子播种育苗。2019年3月利用1 a生轻基质容器苗分别在浙江建德(29°37′N、119°01′E)、江西分宜(27°49′N、114°41′E)和江西安远(25°19′N、115°11′E)营建优树家系测定林,3个试验点均为小叶青冈造林的代表性地区,造林所用家系数分别为 44、35和40个,其中共有家系 35 个(表1)。

表1 参试小叶青冈家系Tab.1 Families of Cyclobalanopsis myrsinifolia used for experimental analysisin
1.2 试验设计与调查
优树家系测定林采用完全随机区组设计,浙江建德点和江西安远点为单列10株小区,江西分宜点为单列8株小区,株行距均为2.5 m×2.5 m,5次重复。2021年11月底对3个地点的测定林开展全林生长(树高、地径和冠幅)和形质(树干通直度、分叉干数、一级侧枝数、最长侧枝长度和最粗侧枝基径)等性状调查。树干通直度从低到高分为5 个等级:严重弯曲(1分)、弯曲(2分)、一般(3分)、较通直(4分)和通直(5分)。
1.3 数据统计分析
选择3个试验点共有家系,采用Microsoft Excel进行数据统计,依据拉依达准则去除异常数据[13]。分叉干数和一级侧枝数经(X+0.5)1/2数据转换,树干通直度经X1/2数据转换。采用 SAS 8.0 软件的GLM、VARCOMP过程对不同地点不同家系生长和形质性状进行单点方差分析,估算家系和单株遗传力、表型和遗传相关系数[10]。
浙江建德点、江西分宜点和安远点生长和形质性状单点方差分析的统计模型如下式。
Yijk=u+Bi+Fj+BFij+eijk
①
式中:Yijk为单点试验第i区组第j家系第k单株的观测值,u为群体平均效应,Bi为第i区组效应,Fj为第j家系效应,BFij为第j家系和第i区组的互作效应,eijk为机误。
家系遗传力计算公式为:
②
单株遗传力计算公式为:
③
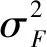
家系同一性状不同试验点两两之间的遗传相关系数按常规的两个性状遗传相关计算,公式为[14]:
rGij=COV(Gij)/δGiδGj
④
上述式中,COV(Gij)为家系同一性状在两个试验点之间的遗传协方差,δGi为该性状在试验点i的遗传方差,δGj为该性状在试验点j的遗传方差。
BLUP值的预测采用固定和随机效应的线性混合效应模型。利用R统计软件的lme4包对测定数据进行一般线性模型拟合,估算家系BLUP值。本研究采用的模型为yij=u+Si+SGij+eij,其中,yij为区组i中基因型j的单株表型值,u为整体均值,Si为区组i的固定效应,SGij为第j个基因型与第i个区组的随机相互作用,eij为残差。
2 结果与分析
2.1 小叶青冈生长和形质性状的家系遗传变异
3个试验点小叶青冈不同家系生长和形质性状均存在极显著的差异(P<0.01),意味着种内存在丰富的遗传变异,为选择优良家系提供了可发掘的变异基础(表2)。3 a生小叶青冈平均树高、地径和冠幅分别为2.18 m、3.02 cm和1.05 m,排序均为建德>安远>分宜。浙江建德点不同家系生长相对较快,与其精细化培育有关,其树高、地径和冠幅的均值分别为2.44 m、3.77 cm和 1.50 m,最大家系分别高出最小家系31.47%、40.47%和40.16%;江西分宜点不同家系生长相对较慢,3 a生平均树高、地径和冠幅分别为1.73 m、2.15 cm 和 0.72 m,最大家系分别高出最小家系43.08%、43.01%和66.76%;江西安远点树高、地径和冠幅的均值分别为2.38 m、3.14 cm和 0.93 m,最大家系分别高出最小家系27.18%、37.67%和44.31%。树干通直度以安远点表现最好,其次为建德点,分宜点的树干通直度相对较低。分叉干数、一级侧枝数量、最长侧枝长度和最粗侧枝基径等在家系间差异同样都达到极显著水平,这有利于选择早期速生和形质优良的小叶青冈家系。分叉干数变异系数以江西安远点最高(73.24%),浙江建德点次之(36.13%),江西分宜点最小(11.65%),局部环境变化或是造林措施对江西安远点小叶青冈家系分叉干数影响较大。
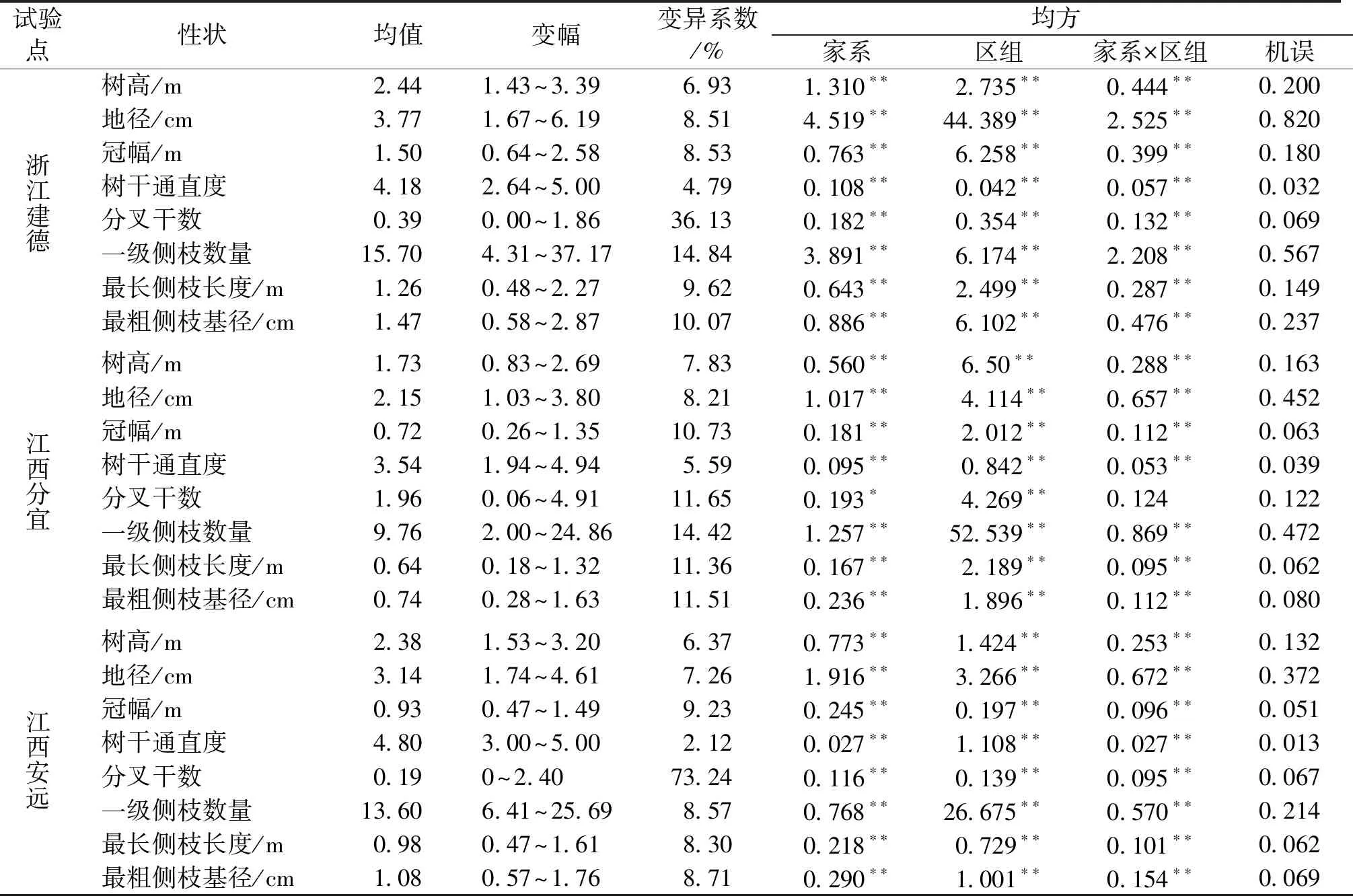
表2 小叶青冈不同家系生长和形质性状的方差分析Tab.2 Variance analysis of growth and form-quality traits of C.myrsinifolia in each test site
2.2 小叶青冈生长和形质性状遗传力估算
遗传力估算结果显示,除江西安远点树干通直度和分叉干数家系遗传力相对较低外(分别为0.19和0.17),小叶青冈各生长和形质性状在各试验点的家系遗传力估算值均大于0.20(表3),可见小叶青冈幼林生长和形质性状受中等至偏强的家系遗传控制,这有利性状稳定优良家系的选择。具体来看,江西安远点树高、地径、冠幅和最长侧枝长度家系遗传力最高,分别为0.71、0.66、0.64和0.57,浙江建德点的树干通直度和一级侧枝数量最高,分别为0.50和0.38,而江西分宜点的分叉干数和最粗侧枝基径最高,分别为0.32和0.53。与家系遗传力相比,3个试验点生长和形质性状的单株遗传力均低于家系遗传力,其值为0.03~0.40,受偏弱至中等强度的遗传控制。单株遗传力较低意味着其基因型值难以通过表型度量值进行预测,应结合优良家系内优良个体的选择以提高遗传增益。
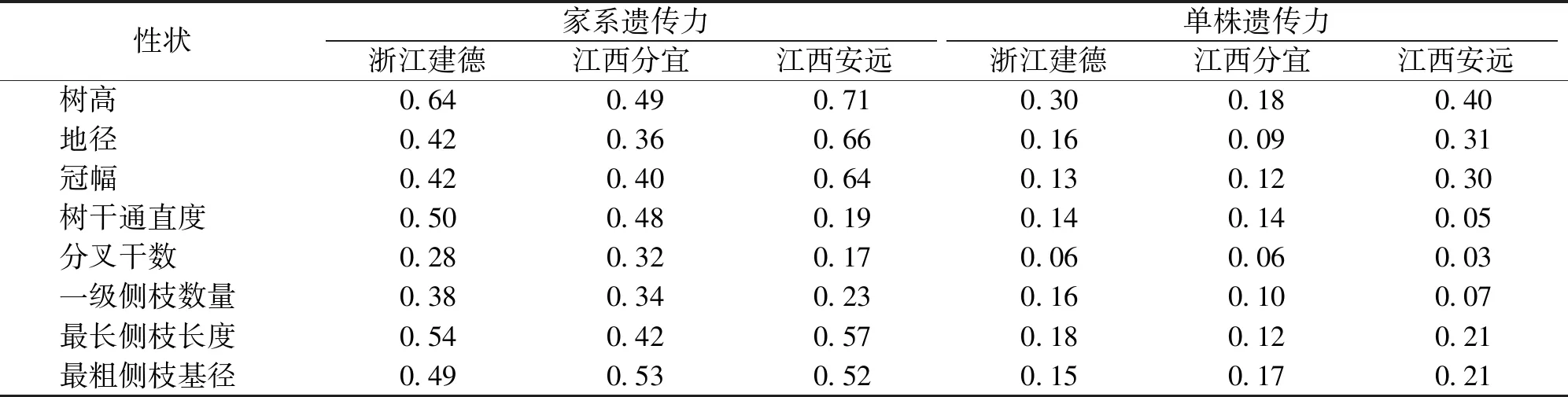
表3 小叶青冈生长和形质性状遗传力估算值Tab.3 Heritability estimation of growth and form-quality traits of C.myrsinifolia
2.3 小叶青冈家系生长和形质性状的表型和遗传相关
相关分析结果表明,不同家系生长性状(树高、地径和冠幅)间呈显著或极显著表型(r=0.421~0.766)和遗传(r=0.460~0.731)正相关(表4)。建德点和分宜点各家系树高、地径和冠幅等生长性状与一级侧枝数、最长侧枝长度和最粗侧枝基径等形质性状均呈显著或极显著表型正相关(r=0.409~0.761),安远点除树高与最粗侧枝基径表型正相关不显著外,树高、地径和冠幅与最长侧枝长度和最粗侧枝基径也呈显著或极显著表型正相关(r=0.365~0.521)。3个试验点除分宜点地径与一级侧枝数遗传正相关不显著外,树高、地径与一级侧枝数、最长侧枝长度间均呈显著或极显著的遗传正相关(r=0.404~0.846),意味着树高和地径生长量越大的家系,其一级侧枝数越多、侧枝较长,若要培育少节或无节的优质干材,应及时加强早期修枝和除萌。3个试验点的家系树干通直度与分叉干数在表型和遗传上均呈弱度和显著的负相关,意味着小叶青冈家系幼树越通直,分叉干数相应地越少。
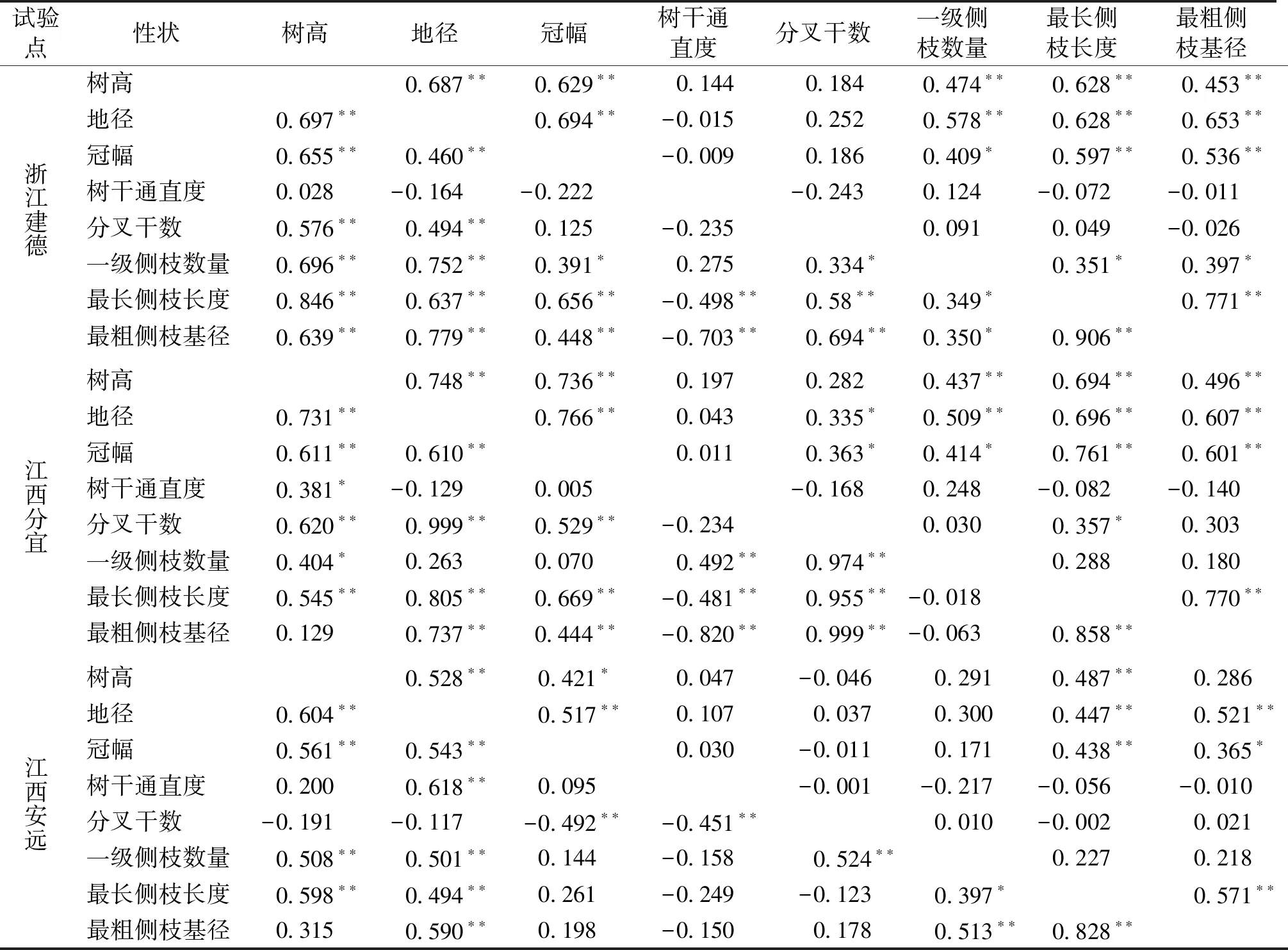
表4 小叶青冈家系生长和形质性状间的表型和遗传相关Tab.4 Phenotypic and genetic correlation coefficients between growth and form-quality traits of C.myrsinifolia
2.4 小叶青冈家系生长和形质性状的地点间遗传相关
表5的地点间遗传相关分析表明,小叶青冈不同家系生长和形质性状在地点间差异较大,两两地点组合间树干通直度均呈显著或极显著正相关,表明其家系×地点互作效应不明显,各家系树干通直度表现稳定,立地条件对其影响较小。两两地点组合间地径、冠幅和最长侧枝长度相关性均不显著,表明存在家系与地点互作效应,即种植生境的改变会对上述指标产生较大的影响。此外,树高、分叉干数、一级侧枝数和最粗侧枝基径等性状在有的地点间相关性显著,有的地点间不显著,家系与立地互作效应无明显变化规律,意味着立地条件的差异对这些指标产生的影响不同。
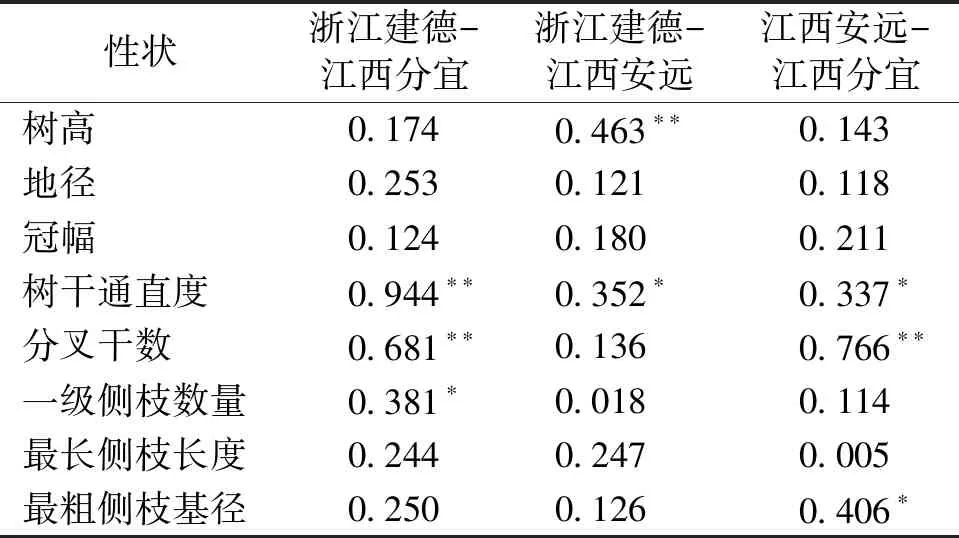
表5 小叶青冈家系生长和形质性状间的地点间遗传相关Tab.5 Correlation coefficient between growth and form-quality traits of C.myrsinifolia
2.5 小叶青冈优良家系
3个试验点根据BLUP方法估算的家系树高和地径BLUP值与实际值呈良好的线性相关,相关系数超过0.9,说明BLUP值能够很好地反映观测值。采用独立淘汰法对3个试验点测定家系的树高和地径BLUP值进行排序,选取排名前20%的家系。按照该标准,浙江建德点共选取89、千9、千10和千11 等4个家系,其平均树高和地径分别为2.64 m和4.15 cm,较家系平均值分别提高了8.32%和10.14%;江西分宜点共选取14、19和千6等3个家系,其平均树高和地径分别为1.89 m和2.38 cm,与家系平均值相比分别提高了9.08%和10.97%;江西安远点共选取14、17、18、千6和90等5个家系,其平均树高和地径分别为2.6 m和3.44 cm,与家系平均值相比分别提高了9.38%和9.64%(图1)。没有在3个试验点均入选的小叶青冈优良家系。

图1 3个试验点小叶青冈优良家系的选择Fig.1 Selection of excellent families of C.myrsinifolia in three experimental sites
3 讨论与结论
3.1 讨论
遗传测定是林木遗传改良的重要环节,目的是通过试验对选择的育种材料的遗传性能进行可靠性评价[15]。林木尤其是阔叶树种幼林生长和形质性状对环境反应敏感[8],早期测定有利于揭示林木早期建成阶段的遗传特点,也有利早期选择。本文通过对浙江建德、江西分宜和江西安远3个试验点建立的3 a生小叶青冈家系测定林进行生长和形质性状研究,发现不同家系间生长和形质性状均存在显著差异,遗传变异丰富,从中选择速生优质的家系和个体对于良种基地建设具有重要意义[16-17]。浙江建德点小叶青冈不同家系的生长表现明显优于江西安远点和分宜点,可能与浙江建德点采用深翻整地造林,并且造林后精细抚育和施肥有关。
遗传力的估算对于育种策略的制定具有重要的指导意义[18]。本研究3个试验点3 a生小叶青冈树高、地径和冠幅的家系遗传力变幅为0.42~0.71,受中等至偏强的遗传控制。3个试验点生长和形质性状家系遗传力均大于单株遗传力,表明通过家系选择可获得较大的遗传增益,单株选择不如家系选择效果好,实际工作中可将家系选择和单株选择结合,以获得更好改良效果。此前对短叶松(P.banksiana)和湿地松(P.elliottii)的研究结果支持这一观点[19-20]。江西安远点家系和单株遗传力总体上高于另外2个试验点,这应与江西安远点重复内立地环境相对一致,试验的环境效应较小有关[2]。其实,研究材料、家系数量、林分年龄、立地条件和试验设计等都能影响性状遗传力的估算值,因此本研究生长和形质性状遗传力估算值也仅作参考。
相关性分析有助于了解不同性状之间的关联程度[21]。本研究小叶青冈不同家系树高、地径与冠幅间呈极显著正相关,可进行联合选择,这与降香黄檀(Dalbergiaodorifera)和红松(P.koraiensis)的研究结果一致[22-23]。树高、地径和冠幅等生长性状与侧枝数量、长度和粗度等也呈显著或极显著的正相关,说明生长性状突出的家系,冠幅相应较大,侧枝多且粗[24]。生产上可通过速生家系选择和合理密植等措施,实现培育高等级小叶青冈优质干材目的。江西分宜点生长和形质性状间的相关关系相对较弱,可能与该试验点幼林生长受环境影响较大有关,不过一级侧枝数与最长侧枝长度和最粗侧枝基径相关性较弱,遗传上较为相互独立,这有利于选择分枝细、分枝角大,即自然整枝容易的家系[10,24]。比较而言,浙江建德和江西安远2个试验点各分枝性状联系紧密,较难进行独立选择。一般来说,如果林木的生长性状遗传力较低,通常意味着基因型×环境互作效应较强[25]。本研究除树干通直度和分叉干数外,其余性状的地点间遗传相关系数为0.005~0.463,整体不高,说明小叶青冈幼林生长和形质性状的家系与立地互作效应较强。从已有研究来看,较之于针叶树种,阔叶树种因对立地反应更敏感,其基因型与环境互作效应相对明显[8-10]。
联合选择可实现林木多性状同步改良效果。对林木各性状设定最低标准值,采用独立淘汰法选择各性状均超过标准值的家系,是多性状选择中最常用的方法,此方法能够使每个性状同时获得改良[26]。本研究基于树高和地径BLUP估算值,分别 3个试验地共初选出12个小叶青冈优良家系,但没有在3个试验点均表现优良的家系,说明小叶青冈家系在不同地点的生长表现差异较大。种植立地和生境的改变对小叶青冈家系生长影响较大,为充分利用基因型×环境互作效应,充分发挥优良家系的良种效益,应分别地点和造林区选育适生的优良家系。小叶青冈虽然是一个相对速生的珍贵阔叶树种,但现阶段3 a生小叶青冈遗传测定林处于幼林阶段,本研究仅是对其生长和形质性状的初步分析和选择,目的是为其速生优良家系的推广应用提供一定参考。
3.2 结论
3 a生小叶青冈生长和分枝性状家系间差异均达到极显著水平,具有丰富的遗传变异。小叶青冈家系遗传力受中等至偏强家系遗传控制,各性状间的相关关系差异较大,树高、地径与一级侧枝数、最长侧枝长度的相关性较强,树高和地径生长突出的家系,冠幅相应较大,侧枝多且粗。种植生境的改变对小叶青冈家系生长影响较大。根据3个试验点的综合表现,初选出12个小叶青冈优良家系,没有在3个试验点均入选的优良家系。为充分发挥赤皮青冈优良家系的良种效益,充分利用基因型与环境的互作效应,建议及时加强其早期修枝和除萌,以培育少节或无节的优质干材。
猜你喜欢
莫愁(2023年9期)2023-03-17
莫愁·家教与成才(2023年3期)2023-03-15
养猪(2022年4期)2022-08-17
文苑·经典美文(2019年9期)2019-09-09
西藏科技(2016年9期)2016-09-26
江苏农业科学(2016年6期)2016-07-25
数学教学通讯·小学版(2016年4期)2016-05-27
中国篆刻·书画教育(2016年2期)2016-03-07
湖北畜牧兽医(2015年11期)2016-01-11
山东农业科学(2014年1期)2015-03-09
