
青钱柳二倍体与四倍体苗期生长和次生代谢产物积累比较1)
2024-01-12 10:16毛乾兴李卓穗卞国良范文杰徐献刚尚旭岚
东北林业大学学报 2024年3期
毛乾兴 李卓穗 卞国良 范文杰 徐献刚 尚旭岚
(南京林业大学,南京,210037) (句容市磨盘山林场) (南京林业大学)
青钱柳(Cyclocaryapaliuru)属胡桃科(Juglandaceae)青钱柳属(Cyclocarya),是中国特有的单种属植物,主要分布于我国亚热带地区[1]。青钱柳常以叶入药,或制成茶叶代茶饮[2],在民间已有悠久的历史。药理研究表明,青钱柳叶提取物具有降血糖、降血压和抗氧化等生物活性[3-6]。前期研究发现,青钱柳在自然界中存在二倍体与同源四倍体两种细胞型[7]。
多倍化是推动植物进化的一个重要因素。由于染色体倍性的提高,多倍体植物常表现为器官的巨大性和较强的抗逆性,而且由于基因的重组,加大了基因表达上的复杂程度,改变了各种代谢途径中酶的活性,从而提高了次生代谢产物的质量分数[8-9]。但Tan et al.[10]对红橘(Citrusreticulata)的研究发现,基因组加倍抑制了果实中黄酮类物质的积累;刘瑛琦[11]的研究也发现,蓝花丹(Plumbagoauriculata)同源四倍体较二倍体植株地上和地下部分的白花丹素质量分数显著降低。植物次生代谢产物通常是利用初生代谢产物合成或分解代谢而来,其合成也需要能量和额外的营养投入[12]。因此,初生代谢产物的分配需要在初生生长和次生代谢间保持一个平衡[13]。已有研究者对不同种源或家系青钱柳苗期生长规律,和黄酮、三萜、多酚等次生代谢产物的积累规律进行了研究[14-15],但尚未见从不同倍性角度开展相关研究的报道。
本研究以二倍体和四倍体青钱柳当年生容器苗为材料,通过定期测定其苗高和地径,对其年生长规律进行模型拟合,比较两种倍性苗木的生长性状、碳氮养分质量分数和主要次生代谢产物积累的差异,并探究两种倍性青钱柳苗木初生生长和次生代谢的关系,为青钱柳种质资源的选育和开发利用提供参考。
1 材料与方法
试验材料:采种母树均来自湖北五峰土家族自治县,2020年春季取茎尖,采用常规压片法制片进行染色体计数,确定母树倍性分别为二倍体(2n=2x=32)和四倍体(2n=4x=64),于2020年10月分单株采种,阴干去翅后,室外自然层积至种子萌发。2022年4月,将株高为3~5 cm的实生苗移栽至无纺布容器(口径8 cm×高度11 cm)中,摆放于南京林业大学园林实验教学中心遮荫大棚下,遮光度约为60%。所用基质为江苏兴农科技有限公司生产的育苗基质,主要由泥炭、椰糠和珍珠岩组成,并添加腐殖酸有机肥。移栽成活后,从二倍体和四倍体中各选择生长一致的苗木180株进行观测,每个倍性3个重复,每个重复60株。
苗高、地径的测定:从2022年4—12月,每月初分别用钢卷尺和游标卡尺测定所有苗木的苗高和地径。地径和苗高曲线拟合采用逻辑斯谛(Logistic)拟合方程y=k/(1+ae-bx),其中,y为苗高或地径生长量;x为时间;a、b为待定系数;k为既定条件的苗高或地径生长可能达到的极限值,可用等差三点法计算求得。
形态指标和生物量的测定:2022年10月上旬,每个重复选取15株长势一致且健壮的标准株,二倍体和四倍体各45株,取从上往下数第四复叶,用WZNDENDRO根系扫描仪分析系统进行扫描,测其面积。同时,每个重复随机挑选6个标准株带回实验室,将根部清洗干净,主根与须根分离并平铺在同一平面,用根系扫描仪分析系统进行扫描,获得各径级根系的长度、面积、体积。将扫描后的叶片、根系与茎分别测定鲜质量,后将其分别放入烘箱内60 ℃烘干至恒定质量,测其干质量,即获得单株叶生物量、单株茎生物量、单株根系生物量和整株生物量。
全碳、全氮质量分数的测定:将测定生物量后的叶、茎、根,分别研磨成粉末,过孔径为0.15 mm的筛子,将过筛后的样品充分混合。每个样品称取100 mg,用锡箔纸包好,均采用元素分析仪(德国碳氮元素分析仪)测定全碳、全氮质量分数。
次生代谢产物质量分数的测定:取叶、茎、根粉末样品,参照林源等[16]的方法进行提取。总黄酮质量分数的测定参照Zhou et al.[17]的方法,总三萜质量分数的测定参照Fan et al.[18]的方法,总多酚质量分数的测定参照游见明等[19]的方法。次生代谢产物质量分数以1 g青钱柳叶干样中含有次生代谢产物的质量表示(单位为mg·g-1);次生代谢产物积累量以次生代谢产物质量分数与相应叶生物量的乘积表示(单位为mg·株-1)。
数据处理:运用Excel 2019软件将试验所得数据进行统计分析,用SPSS 26.0软件对各指标进行独立样本t检验,苗高和地径进行逻辑斯谛(Logistic)非线性回归分析,并对初生生长指标与次生代谢指标进行相关性(Person)分析。
2 结果与分析
2.1 青钱柳二、四倍体苗期生长节律差异
为了解不同倍性青钱柳1年生苗的年生长节律,运用逻辑斯谛方程分别拟合二倍体和四倍体的苗高和地径生长过程,方差分析结果和拟合方程见表1。两种倍性苗高、地径的逻辑斯谛方程拟合模型回归关系均达到极显著水平。
由图1可知,二倍体与四倍体青钱柳的苗高和地径生长节律基本相似,呈S型曲线增长。5月份的生长速率较缓,6月开始加快,8月达到最大生长速率,9月生长逐渐趋于平缓。根据逻辑斯谛方程求导所得拐点(表1)及移栽时间(4月9日),可将1年生苗的生长阶段划分为生长初期、速生期、生长后期3个时期。
结合实际观测和理论计算,将二倍体1年生苗的苗高生长划分为,生长初期(4月9日—6月2日)、速生期(6月3日—9月2日)、生长后期(9月3日—11月末);四倍体1年生苗的苗高生长划分为,生长初期(4月9日—6月14日)、速生期(6月15日—9月26日)、生长后期(9月27日—11月末)。二倍体苗高速生期持续时间为91d,四倍体苗高速生期持续时间为103 d,四倍体苗高速生期比二倍体速生期延长12d。速生期内,二、四倍体1年生苗苗高生长量分别约占总生长量的48.9%、51.2%。
二倍体1年生苗的地径生长划分为,生长初期(4月9日—6月10日)、速生期(6月11日—8月30日)、生长后期(8月31日—11月末);四倍体1年生苗的地径生长划分为,生长初期(4月9日—6月12日)、速生期(6月13日—9月15日)、生长后期(9月16日—11月末)。二倍体地径速生期持续时间为80 d,四倍体持续时间为94 d,四倍体地径速生期比二倍体速生期延长14 d。速生期内,二、四倍体1年生苗地径生长量分别约占总生长量的47.5%、49.7%。

表1 1年生青钱柳苗高和地径的逻辑斯谛回归方程
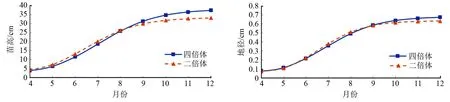
图1 1年生青钱柳苗高和地径的逻辑斯谛拟合曲线
2.2 青钱柳二、四倍体生长性状及生物量差异
青钱柳二倍体和四倍体生长性状情况见表2。四倍体青钱柳复叶面积比二倍体增加了23.6%,根体积增加了16.2%,都达到显著差异水平(P<0.05),直径大于2 mm的根系体积比二倍体增加了57.4%,达到极显著差异水平(P<0.01)。另外,四倍体青钱柳总叶生物量、茎生物量、根生物量及总生物量比二倍体分别增加了44.0%、16.8%、52.6%、40.0%,均达到显著差异水平。结合根系扫描图片(图2)分析,四倍体主根较粗大,根系生物量的积累主要集中在主根,侧根较二倍体少。可见,四倍体青钱柳在叶片和主根的性状上表现出巨大性。

表2 青钱柳二倍体与四倍体生长性状及生物量
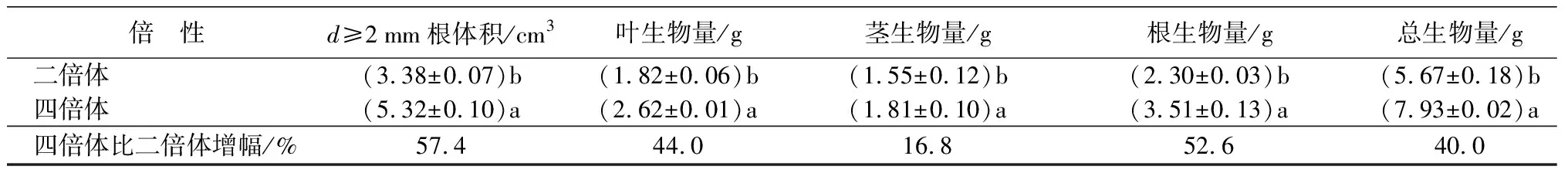
倍 性d≥2mm根体积/cm3叶生物量/g茎生物量/g根生物量/g总生物量/g二倍体(3.38±0.07)b(1.82±0.06)b(1.55±0.12)b(2.30±0.03)b(5.67±0.18)b四倍体(5.32±0.10)a(2.62±0.01)a(1.81±0.10)a(3.51±0.13)a(7.93±0.02)a四倍体比二倍体增幅/%57.4 44.0 16.8 52.640.0

A为青钱柳四倍体叶片形态,B青钱柳二倍体叶片形态;a为青钱柳四倍体根系形态,b为青钱柳二倍体根系形态。
2.3 青钱柳二、四倍体碳氮养分质量分数差异
由表3可知,四倍体青钱柳叶片的氮质量分数显著低于二倍体(P<0.05),碳质量分数在两种倍性间无显著差异,碳、氮质量分数比则四倍体显著高于二倍体(P<0.05);四倍体青钱柳茎的碳质量分数显著高于二倍体(P<0.05),氮质量分数和碳、氮质量分数比在两种倍性间均无显著差异;四倍体青钱柳根系碳、氮质量分数均显著低于二倍体(P<0.05),但碳、氮质量分数比四倍体显著高于二倍体(P<0.05)。

表3 青钱柳二倍体与四倍体碳氮养分质量分数比较
2.4 青钱柳二、四倍体次生代谢产物积累差异
青钱柳植株各器官次生代谢产物质量分数的测定结果见表4,青钱柳四倍体叶片中总黄酮、总多酚质量分数显著低于二倍体(P<0.05),三萜质量分数无显著差异;青钱柳四倍体与二倍体茎总黄酮、总多酚和总三萜质量分数均无显著差异;青钱柳四倍体根系总黄酮、总多酚和总三萜质量分数显著低于二倍体(P<0.05)。
青钱柳次生代谢产物积累量结果显示,四倍体茎和根系黄酮积累量显著高于二倍体(P<0.05);四倍体叶片多酚积累量显著高于二倍体(P<0.05);四倍体叶片三萜积累量显著高于二倍体(P<0.05);其余各器官的次生代谢产物积累量均未呈现显著差异。

表4 青钱柳二、四倍体叶、茎、根次生代谢产物质量分数与积累量

倍性总多酚质量分数/g·kg-1叶茎根总多酚积累量/mg·株-1叶茎根二倍体(35.45±0.40)a(12.50±1.56)a(33.68±1.32)a(64.60±0.72)b(18.44±1.03)a(75.10±3.79)a四倍体(31.35±0.89)b(17.45±2.69)a(23.26±4.18)b(81.99±2.33)a(29.76±5.22)a(79.24±3.94)a

倍性总三萜质量分数/g·kg-1叶茎根总三萜积累量/mg·株-1叶茎根二倍体(25.41±0.29)a(20.01±2.64)a(45.04±2.10)a(46.30±0.53)b(29.92±1.91)a(100.07±2.71)a四倍体(26.98±0.27)a(26.36±4.34)a(28.56±5.55)b(70.54±0.70)a(48.11±6.48)a(98.15±4.06)a
2.5 青钱柳苗期生长性状与次生代谢产物质量分数与积累量的相关性分析
从表5可见,两种倍性青钱柳的苗高、地径、复叶面积、根体积、叶生物量和根系生物量之间均呈极显著正相关;比叶质量与苗高、地径、复叶面积、根体积、叶生物量、根生物量、叶碳、氮质量分数比呈显著或极显著正相关;叶碳、氮质量分数比与苗高、地径、复叶面积、根体积、叶生物量、茎生物量、根生物量呈显著正相关。
青钱柳苗高、地径、复叶面积、叶生物量、根生物量和叶碳、氮质量分数比与叶片黄酮和叶片多酚质量分数呈显著或极显著负相关;苗高、地径、复叶面积、根体积、叶生物量和根生物量与叶三萜质量分数呈显著正相关;茎碳、氮质量分数比与茎黄酮质量分数呈显著负相关。
青钱柳苗高、地径、复叶面积、根体积、叶生物量、比叶质量、茎生物量、根生物量与叶多酚、叶三萜和茎黄酮积累量呈显著或极显著正相关;苗高、地径、复叶面积、叶生物量、根生物量与根黄酮积累量呈显著正相关;叶片氮质量分数与叶三萜积累量、茎黄酮和茎三萜积累量、根黄酮积累量呈显著负相关;叶片碳、氮质量分数比与叶多酚、叶三萜、茎黄酮、茎多酚和茎三萜积累量呈显著正相关。
3 讨论
多倍体植株普遍表现出更优越的农艺性状,如生活力强、生长迅速、器官增大、代谢物质量分数增加等,因而具有较高的育种价值和应用前景[20]。林木多倍体育种在短周期森林工业用材树种、收获次生代谢产物的经济林木和生态防护林木新品种选育等方面都发挥了重要作用[21]。多倍体培育技术已应用于杜仲(Eucommiaulmoides)[22]、黄梁木(Neolamarckiacadamba)[23]、黄连木(Pistaciachinensis)[24]等木本药用植物,获得了生物量和有效成分质量分数高的种质材料。
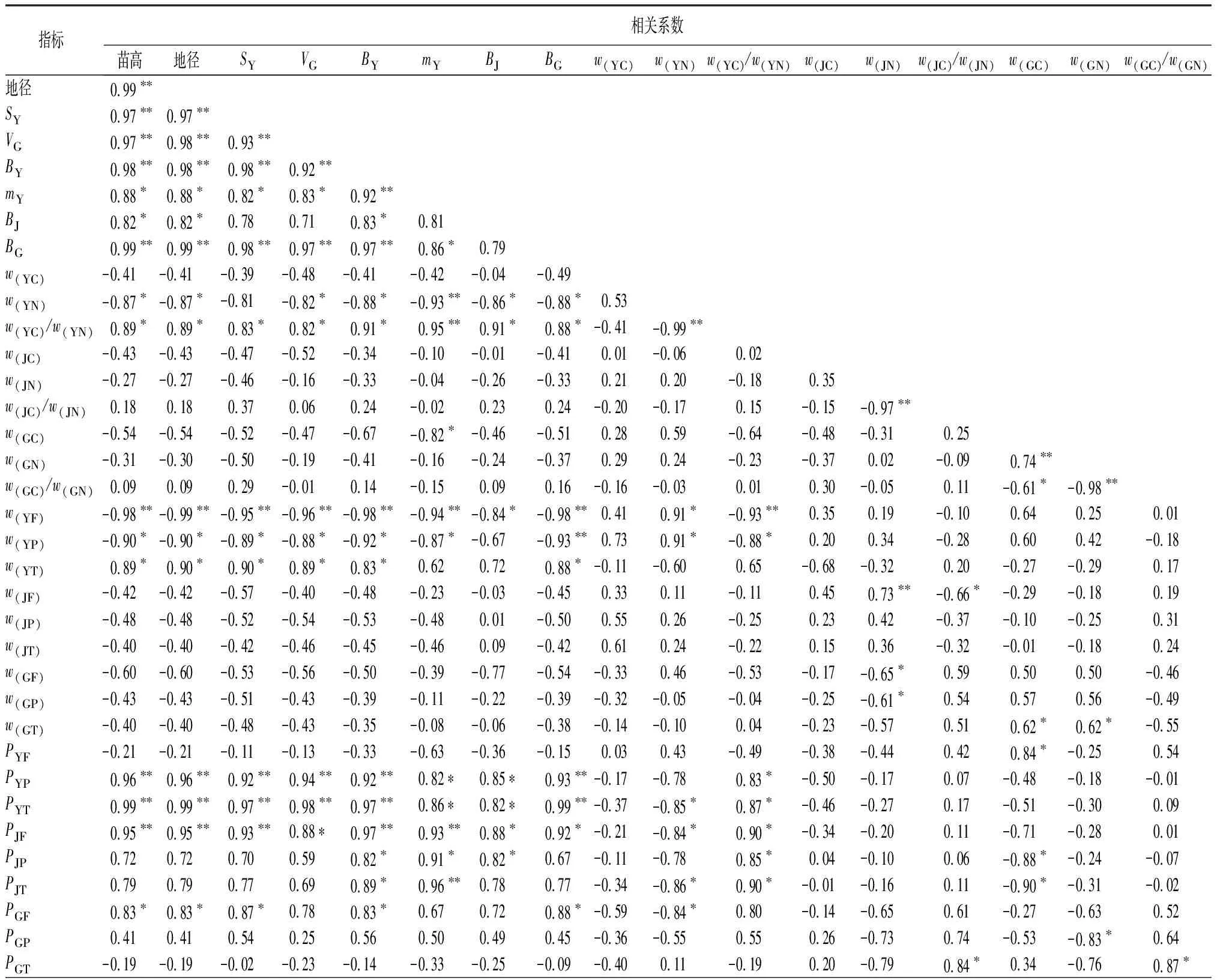
表5 青钱柳苗期生长性状与次生代谢产物积累的相关系数

指标相关系数w(YF)w(YP)w(YT)w(JF)w(JP)w(JT)w(GF)w(GP)w(GT)PYFPYPPYTPJFPJPPJTPGFPGPw(YP)0.91∗w(YT)-0.84∗-0.65w(JF)0.360.45-0.47w(JP)0.500.57-0.370.88∗∗w(JT)0.430.54-0.230.83∗∗0.99∗∗w(GF)0.560.25-0.75-0.70∗-0.70∗-0.68∗w(GP)0.350.10-0.74-0.55-0.54-0.520.92∗∗w(GT)0.320.14-0.68-0.51-0.49-0.460.90∗∗0.99∗∗PYF0.350.260.10-0.200.210.28-0.06-0.31-0.35PYP-0.95∗∗-0.750.95∗∗-0.33-0.37-0.26-0.76-0.56-0.49-0.20PYT-0.98∗∗-0.88∗0.92∗∗-0.42-0.47-0.37-0.63-0.48-0.44-0.180.98∗∗PJF-0.97∗∗-0.810.84∗-0.38-0.46-0.37-0.60-0.44-0.37-0.430.95∗∗0.95∗∗PJP-0.79-0.670.50-0.21-0.35-0.31-0.36-0.13-0.04-0.750.700.700.88∗PJT-0.86∗-0.810.53-0.31-0.48-0.46-0.27-0.09-0.05-0.700.720.770.90∗0.97∗∗PGF-0.79-0.89∗0.69-0.42-0.27-0.20-0.40-0.16-0.140.060.700.82∗0.700.510.63PGP-0.43-0.580.19-0.47-0.25-0.230.100.080.11-0.290.260.380.480.630.680.66PGT0.26-0.08-0.22-0.380.040.040.450.210.140.54-0.36-0.21-0.33-0.34-0.230.350.51
本研究结果表明,青钱柳二倍体与四倍体苗木的生长节律基本相似,都呈“S”型曲线生长,但四倍体较二倍体的速生期延长了约两周。生长期延长,有利于植株生物量的积累,如贾明良等[25]的研究发现,十六倍体半夏(Pinelliaternata)比八倍体半夏的生长期延长,产量显著增加。本研究还表明,四倍体青钱柳复叶面积、根系体积与主根体积均显著高于二倍体,且叶生物量、根生物量和总生物量均有显著提升,这说明四倍体青钱柳地上部分与地下部分都表现出了巨大性的特点。张石虎等[26]对黄梁木的研究表明,与二倍体相比,四倍体茎更粗、叶面积更大、生物量更高。林玉凤[27]对穿心莲(Andrographispaniculata)的研究发现,四倍体茎更粗、叶面积更大、生物量更高,但植株较矮。青钱柳是我国极具推广价值的药食同源植物,目前主要采收叶片进行加工利用,四倍体青钱柳较二倍体有更高的生物量,因此,具有较大的开发利用潜力。
碳和氮是植物器官以及代谢产物的基本组成元素,化学计量比与植物生长和代谢密切相关,植物叶片的碳、氮质量分数比在一定程度上反映了植物的营养利用效率,具有重要的生态学意义[28-29]。本研究结果表明,四倍体青钱柳叶片与根系碳、氮质量分数比显著高于二倍体,相关性分析结果也显示,青钱柳叶碳、氮质量分数比与苗高、地径、生物量等多个生长指标呈极显著或显著正相关,说明四倍体具有更高的营养利用效率,相同的养分条件能获得更大的生物量。
对青钱柳叶、茎、根中次生代谢产物质量分数的测定结果表明,两种倍性苗木的总黄酮与总多酚质量分数均为叶片中最高,而总三萜质量分数为根系中最高。通过比较发现,四倍体青钱柳叶片与根系中的总黄酮和总多酚质量分数均显著低于二倍体,而四倍体根系中总黄酮、总多酚和总三萜质量分数也显著低于二倍体。次生代谢过程被认为是植物在长期进化中对生态环境适应的结果,它在处理植物与生态环境的关系中充当着重要的角色[8]。植物的生长与次生代谢之间普遍存在着权衡关系,而这种关系对药用植物药效成分的积累至关重要[12]。相关性分析结果表明,青钱柳叶片总黄酮和总多酚质量分数与苗高、地径、复叶面积、比叶质量、碳氮质量分数比等生长性状指标呈显著负相关,说明青钱柳初生生长和次生代谢间存在着一种权衡关系。本研究结果显示,虽然四倍体叶中总黄酮和总多酚质量分数较低,但由于四倍体叶生物量显著大于二倍体,因此两种倍性叶中总黄酮积累量无显著差异,四倍体叶总多酚和总三萜积累量显著高于二倍体。但应注意的是,本研究结果仅以1年生容器苗为材料观测所得,随着苗木的生长发育,两种倍性植株的生长和次生代谢会如何改变,还需进行长期的研究。
4 结论
1年生青钱柳二倍体与四倍体苗木的生长节律基本相似,但四倍体的速生期较二倍体延长了约两周;四倍体复叶面积、根体积、根生物量、叶生物量和总生物量显著高于二倍体;四倍体叶片和根系碳氮质量分数比显著高于二倍体,说明四倍体具有更高的营养利用效率,能获得更大的生物量;四倍体青钱柳叶片中的总黄酮和总多酚质量分数均显著低于二倍体,四倍体根系中总黄酮、总多酚和总三萜质量分数也显著低于二倍体,但由于四倍体叶生物量显著大于二倍体,因此两种倍性叶中总黄酮积累量无显著差异,四倍体叶总多酚和总三萜积累量显著高于二倍体。相关性分析结果表明,青钱柳叶片总黄酮和总多酚质量分数与苗高、地径、复叶面积、比叶质量、碳氮质量分数比等生长性状指标呈显著负相关,说明青钱柳初生生长和次生代谢间也存在着一种权衡关系。
猜你喜欢
农业科技通讯(2023年1期)2023-02-12
农业技术与装备(2020年7期)2020-08-18
现代园艺(2017年13期)2018-01-19
江苏农业科学(2017年16期)2017-10-27
中成药(2017年6期)2017-06-13
现代农业科技(2017年8期)2017-06-10
中成药(2017年4期)2017-05-17
中成药(2016年4期)2016-05-17
浙江林业(2015年7期)2015-02-24
中国果业信息(2015年6期)2015-01-23
