
2017-2022年雅砻江甘孜以下干流及部分支流鱼类种类组成及群落结构*
2024-01-13 07:40熊美华董微微刘小帅宋以兴胡兴坤
湖泊科学 2024年1期
熊美华,董微微,刘小帅,朱 滨,宋以兴,胡兴坤,曾 昌,余 丹,邵 科**
(1:水利部中国科学院水工程生态研究所,水利部水工程生态效应与生态修复重点实验室,湖北省水生态保护与修复工程技术研究中心,武汉 430079) (2:雅砻江流域水电开发有限公司,成都 610056)
雅砻江(26°32′~33°58′N,96°52′~102°48′E)处于青藏高原东南缘,是金沙江最大的支流,也是长江的八大支流之一[1]。雅砻江河源至新龙县乐安乡尼托村为上游河段,尼托至理塘河口为中游河段,理塘河口至雅砻江金沙江汇口为下游河段[2]。雅砻江全流域地势总体表现为西北高、东南低,由源头海拔超过5000 m降至河口约980 m,河流下切强烈,地形十分复杂[1];地貌上跨越了高原区、盆地中山区等;气候上跨越南、中、北亚热带,暖温带,寒温带等气候区;上、中、下游水文特点差异较大,造就了雅砻江流域独特的鱼类组成、资源类型、区系分布的突出特征[3-9]。对雅砻江流域层面的鱼类资源调查主要有邓其祥[4]在1968年、1980-1984年在雅砻江干支流开展的鱼类调查,吴江等[5]在1983-1984年间开展的雅砻江渔业自然资源调查,丁瑞华通过收集和检视有关单位的标本整理了包括雅砻江流域在内的四川鱼类[10],张春光等在2008-2017年间开展了金沙江流域包括雅砻江的鱼类调查[11],郭延蜀等通过近20余年的调查和收集整理出版了包括雅砻江鱼类的《四川鱼类原色图志》[12]。其他相关研究多集中在区域层面或局部水域,如雅砻江下游[6-7,9]、锦屏一级和二级电站[13-15]工程水域、二滩库区[16-19]及雅砻江支流如力丘河[20]、鳡鱼河[21]、安宁河[22-23]等的调查。
雅砻江流域水力资源丰富,干流共规划21座水电站,下游5座梯级水电站目前已全部建成运行;中游两河口、杨房沟水电站已于2021年蓄水发电[24-25]。梯级电站的建设运行会导致一系列水文、生境等因素的变化,如阻隔河流连通性,改变河流自然形态从而引起河相向湖相的转变,且电站不同运行时长等因素都会对鱼类群落组成和结构造成影响[26-29]。目前仅邓其祥等[16-17]、蒋红等[18]和何莎[19]基于二滩水库修建前后鱼类资源的变化,分析了二滩水库建成后鱼类种类组成的演变,但缺乏对现阶段流域层面鱼类资源分布状况等的研究报道。
本文基于2017-2022年对雅砻江流域鱼类资源进行的较系统的调查及该期间相关学者所发表的文献资料,分析在雅砻江这种特殊地形地貌背景下鱼类随流域纵向梯度变化的情况,为探讨鱼类群落构建机制及相关影响因素提供基础;结合该流域鱼类历史资料,分析雅砻江鱼类种类组成和结构的变化,为该流域鱼类种质资源保护提供科学依据;结合干流梯级电站建设完成情况,分析鱼类种群聚类情况及优势种,提出在下游梯级电站运行背景下土著鱼类的针对性保护措施建议。
1 材料与方法
1.1 样品采集地点
2017-2022年根据鱼类区系研究方法,对雅砻江流域进行鱼类资源调查,调查范围包括甘孜至雅砻江河口等干流区域及与干流水电工程相关的支流,其中,上游、中游、下游干流分别设置2、3、8个采样点,中游支流鲜水河、卧龙寺沟和霍曲河分别设置2、 1和 1个,下游支流理塘河、永兴河和鳡鱼河分别设置2、1和1个,具体采样点设置情况见图1和附表I。河流等级的划分依据Horton[30]提出的水系组成理论,把干流作为1级河流,汇入干流的支流作为2级河流,汇入2级河流的支流作为3级河流。

图1 雅砻江流域鱼类采样点分布Fig.1 Distribution of fish sampling sites in the Yalong River
1.2 样品采集方法
采取科研特许捕捞方法,采用刺网、地笼、饵钓钩等(附表I)网具在捕捞许可范围内采集鱼类标本、测量生物学信息、做好记录,对于存活鱼类将其放回捕捞水域,对于已死亡鱼类,将其用10%福尔马林溶液固定保存以制作成标本,对不确定种类采集分子样品,并用无水乙醇保存。鱼类种类依据形态特征参考相关文献和书籍[10-12,31-33]进行鉴定,鱼名拉丁名参照Fishbase、郭延蜀等[12]的标准,逐尾测量鱼类体质量(精确至0.1 g)等生物学指标,记录采集时间和采集地点等相关信息。通过对样本的分类鉴定,资料的分析整理,并综合2017-2022年所发表文献[13,20],编制出雅砻江流域鱼类种类组成名录表。其中,本调查中将福建纹胸鮡(Glyptothoraxfokiensis(Rendahl))改为了中华纹胸鮡(Glyptothoraxsinensis(Regan))[34],裂腹鱼属(Schizothorax)鱼类采用形态鉴定和分子鉴定相结合的方式确定种类,分子鉴定所用的标记为线粒体Cytb基因。
1.3 数据处理与分析
采用Shannon多样性指数(H′)[35]、Simpson优势度指数(C)[36]、Margalef种类丰富度指数(R)[37]、Pielou均匀度指数(J)[38]分析不同河段鱼类群落结构特征,各指数值依据每种鱼类尾数,采用Microsoft Excel 2019进行统计和计算获得。
基于原始渔获物丰度矩阵,经平方根转换后以Bray-Curtis相似性系数为基础构建不同采样点的相似性矩阵,采用层次聚类Cluster方法和非度量多维尺度分析(NMDS)对各采样区域鱼类群落进行分组,了解各采样区域鱼类群落结构的变化。依据协强系数(Stress)来衡量NMDS分析结果的可信度:当Stress<0.05时,具有很好的代表性,可信度最高;当Stress<0.1时,可以认为是一个好的排列;当Stress<0.2时,认为该结果有一定的解释意义,但有部分细节不可靠,可用NMDS的二维点图表示[39-40]。采用单因素的相似性分析(one-way ANOSIM)对分组数据进行差异检验,判断该分组结果的正确性[41]。Cluster、NMDS和ANOSIM检验通过生态统计软件Primer 6.0进行。
以相对重要性指数(index of relative importance, IRI)判断鱼类群落优势种[42]。将IRI值≥1000的物种确定为优势种,将1000>IRI值≥500的物种确定为常见种。
按照鱼类栖息的环境、食性和产卵类型进行生态类型划分。依据鱼类栖息的环境可将鱼类划分为急流型、缓流型和静水型;按食性可划分为肉食性、杂食性和植食性;按产卵类型可划分为(黏)沉性卵、浮性卵、黏性卵、漂流性卵、蚌内产卵、筑巢产卵和体内受精等[43],其中,将具有微黏性但沉在水底发育的黏沉性卵与沉性卵划分为同一类,即(黏)沉性卵。
所有分析数据均为本研究野外调查数据,不包含文献记载数据。
2 结果
2.1 种类组成
2017-2022年间在雅砻江流域共收集到渔获物8691尾,体质量达124.28 kg;隶属于3目12科59属94种(亚种),结合相关文献资料[13,20]整理得到2017-2022年间雅砻江流域鱼类98种(亚种)(附表Ⅱ),隶属于4目13科,其中鲤形目鱼类种类数最多,包含鲤科、平鳍鳅科和鳅科3科,共72种,占鱼类种类总数的73.5%;其次为鲇形目鱼类,包括鲇科、鲿科、钝头鮠科、鮰科和鮡科5科,共19种鱼类,占鱼类种类总数的19.4%;鲈形目鱼类包括4科,为丽鱼科、鮨科、太阳鱼科和虾虎鱼科,共6种,占比为6.1%;鲑形目鱼类包括1科1种。其中,境外引入种有6种,为大鳞鲃(Luciobarbuscapito(Güldenstädt))、散鳞镜鲤(CyprinuscarpiospecularisLacepède)、斑点叉尾鮰(Ictaluruspunctatus(Rafinesque))、虹鳟(Oncorhynchusmykiss(Walbaum))、尼罗罗非鱼(Oreochromisniloticus(Linnaeus))、大口黑鲈(Micropterussalmoides(Lacepède));黄河特有种1种,为花斑裸鲤(GymnocypriseckloniHerzenstein)[12]。
在雅砻江上游干流共监测到鱼类9种,均隶属于鲤形目鱼类,包含鲤科和鳅科,其中鳅科仅1种,为泥鳅(Misgurnusanguillicaudatus(Cantor)),其他8种均为鲤科鱼类,其中5种为裂腹鱼属鱼类。中游干流共监测到鱼类26种,除4种为鲇形目鱼类外,其他均为鲤形目鱼类,其中鲤科鱼类最多,有15种,平鳍鳅科鱼类次之,有4种,在属层面,裂腹鱼属鱼类最多,有6种。下游干流共监测到鱼类82种,其中鲤形目鱼类56种,鲇形目鱼类19种,鲈形目鱼类6种,鲑形目鱼类1种。从上游、中游到下游,鱼类种类数逐渐增多。雅砻江支流中,鲜水河共采集到11种鱼类,鱼类种类与上游干流类似,卧龙寺沟共采集到5种鱼类,霍曲河采集到3种鱼类,力丘河记载鱼类13种[20];下游支流理塘河共采集到17种鱼类,鳡鱼河采集到13种鱼类,鳡鱼河支流永兴河采集到7种鱼类。
在上游、中游和下游河段均采集到4种裂腹鱼属鱼类,分别为短须裂腹鱼(Schizothoraxwangchiachii(Fang))、长丝裂腹鱼(SchizothoraxdolichonemaHerzenstein)、齐口裂腹鱼(Schizothoraxprenanti(Tchang))和四川裂腹鱼(SchizothoraxkozloviNikolsky),其中,在支流鲜水河、力丘河和理塘河也采集到了这4种鱼类,另一种细鳞裂腹鱼(Schizothoraxchongi(Fang))在卡拉以上河段未监测到;支流鳡鱼河未监测到裂腹鱼属鱼类,且鳡鱼河与其支流永兴河未监测到共有鱼类。
依据生态类型(图2)进行分析,按栖息类型,急流型鱼类分别占总数量和总体质量的47.91%和61.93%,静水型鱼类分别占总数量和总体质量的29.89%和19.44%,缓流型鱼类分别占总数量和总体质量的22.2%和18.63%;按食性分类,杂食性鱼类分别占总数量和总体质量的78.09%和78.30%,肉食性鱼类分别占总数量和总体质量的21.36%和18.73%,植食性鱼类分别占总数量和总体质量的0.55%和2.97%;按产卵类型,产黏性卵鱼类分别占总数量和总体质量的49.12%和24.75%,产(黏)沉性卵鱼类分别占总数量和总体质量的39.33%和61.24%,产漂流性卵鱼类分别占总数量和总体质量的7.18%和11.94%,产浮性卵鱼类分别占总数量和总体质量的0.51%和0.91%,筑巢产卵鱼类分别占总数量和总体质量的2.16%和0.97%,体内受精产沉性卵鱼类分别占总数量和总体质量的1.19%和0.18%,蚌内产卵鱼类分别占总数量和总体质量的0.52%和0.01%。

图2 雅砻江流域鱼类生态类型组成Fig.2 Composition of fish ecological types in the Yalong River
2.2 珍稀特有鱼类监测结果
2017-2022年间通过监测和整理获得圆口铜鱼(Coreiusguichenoti(SauvageetDabry))、长鳍吻鮈(Rhinogobioventralis(SauvageetDabry))、金沙鲈鲤(Percocyprispingi(Tchang))、细鳞裂腹鱼、重口裂腹鱼(Schizothoraxdavidi(Sauvage))、厚唇裸重唇鱼(GymnodiptychuspachycheilusHerzenstein)、岩原鲤(Procyprisrabaudi(Tchang))、长薄鳅(Leptobotiaelongata(Bleeker))和青石爬鮡(Euchiloglanisdavidi(Sauvage))9种国家二级重点保护鱼类;监测到长江上游特有鱼类雅砻白鱼(AnabariliusliuiyalongensisLietChen)、厚颌鲂(Megalobramapellegrini(Tchang))、圆口铜鱼等28种(附表Ⅱ),其中在干流上、中、下游监测到的特有鱼类种类数分别为5、13、22种,与各河段整体鱼类种类数变化趋势一致,在支流鲜水河、卧龙寺沟、霍曲河、力丘河、理塘河、鳡鱼河和永兴河分别监测和整理得到特有鱼类7、1、3、8、8、1和4种,变化趋势与各支流整体鱼类种类数不同。
2.3 生物多样性分析
从图3可以看出,干支流各区域Pielou均匀度指数和Simpson优势度指数没有明显差异;Margalef丰富度指数和鱼类物种数变化趋势较一致,在雅砻江下游河段最高;支流中鲜水河和理塘河丰富度指数值较高,卧龙寺沟和霍曲河较低,均低于1;干支流各区域Shannon多样性指数值有一定波动,在上、中、下干流区域多样性相差不大,支流理塘河多样性指数值最高,霍曲河最低。

图3 干流上、中、下游及各支流鱼类生物多样性指数Fig.3 Fish community biodiversity index in the Yalong River
将雅砻江鱼类按照干支流分成2组,干流上、中、下游为第1组,6条支流为第2组,对两组所有指数值进行单因素方差分析,结果表明:Shannon指数(F=5.07,P=0.059)、Simpson优势度指数(F=2.80,P=0.138)、Margalef丰富度指数(F=4.48,P=0.072)和Pielou均匀度指数(F=1.47,P=0.265)等在0.05水平上差异均不显著,说明干支流鱼类生物多样性不存在显著差异。
2.4 鱼类群落结构及相似性分析
依据21个采样点的鱼类种类和相对丰度数据分析鱼类群落结构,Cluster聚类分析表明:在44.36%的Bray-Curtis相似性水平上,可将雅砻江干支流21个采样河段分成6组(图4),组1为下游官地至中游卡拉的干支流河段,具体包括干流5个采样点(GZ3、GX1至GX4)和支流理塘河2个采样点(LTH1、LTH2);组2为卡拉往上中上游干流及支流鲜水河河段,具体包括上游干流2个采样点(GS1、GS2)、中游干流2个采样点(GZ1、GZ2)和支流鲜水河2个采样点(XSH1、XSH2),2021年建成的杨房沟水电站位于该聚类组所有采样点以下,两河口水电站2021年运行,仅GZ2采样点样品是运行后采集的,因此该组鱼类所栖息的河段基本处于天然状态,几乎还未受电站蓄水运行影响;组3为鳡鱼河支流永兴河;组4包括GX5(金河)、GX7(得石)、GX8(雅砻江河口)等二滩电站建设运行后依旧留存的3个流水河段;组5为GX6(二滩)和GYH(鳡鱼河)两个静缓流河段;组6为中游霍曲河(HQH)和卧龙寺沟(WLSG)两个支流河段。One-way ANOSIM检验结果表明这6组间的群聚结构在统计学上的差异显著(全局R=0.951,P=0.001,置换检验999次)。NMDS分析的胁强系数0.08<0.1,表明该聚类结果基本可信,是一个好的排列(图5)。
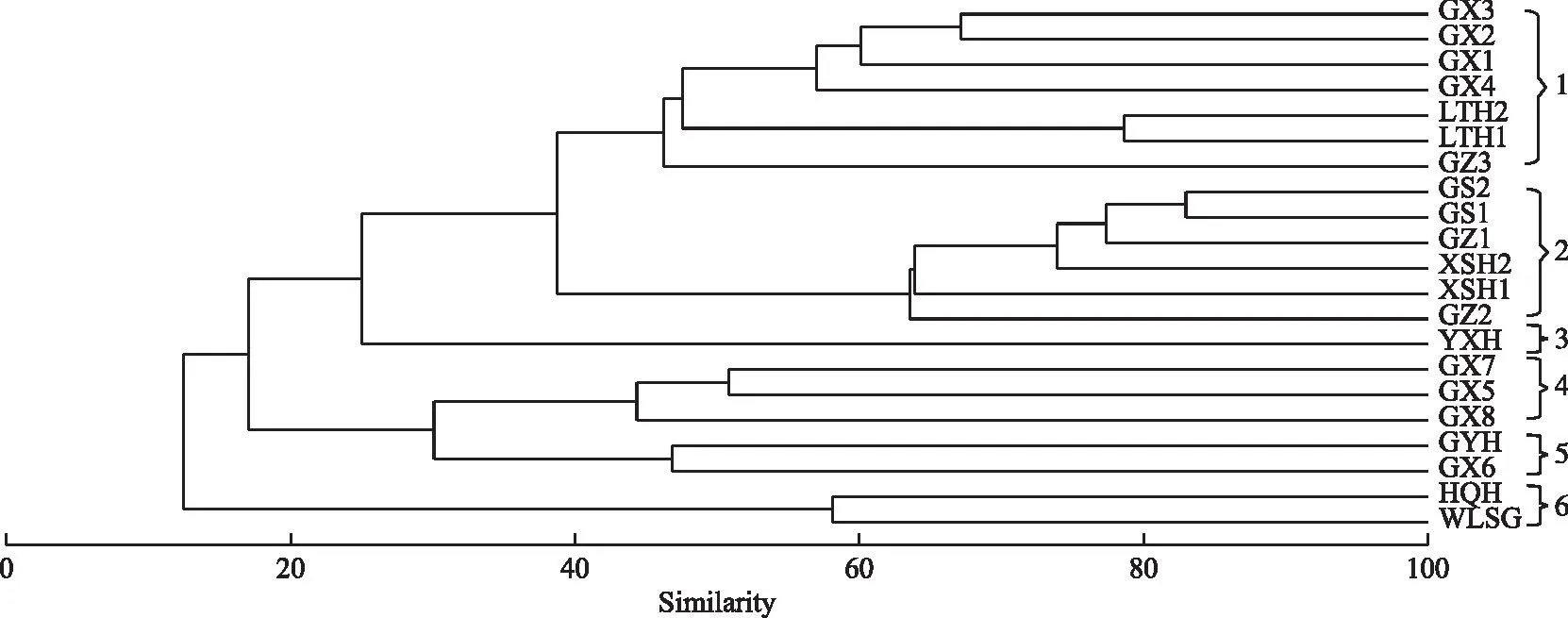
图4 21个采样河段鱼类Cluster聚类分析Fig.4 The cluster analysis of fishes at 21 sampling sites

图5 21个采样河段鱼类非度量多维尺度分析(NMDS)Fig.5 NMDS analysis of fishes at 21 sampling sites
对6个聚类组的海拔和河流等级进行单因素方差分析,结果显示,6个组在海拔上(F=31.53,P<0.001)存在极显著差异,在河流等级上(F=3.57,P<0.05)存在显著性差异。
2.5 优势种及生态类型
依据Cluster聚类分析结果,雅砻江鱼类被分成6组,在此基础上分析每组的优势种、常见种、平均体重及其生态类型情况,结果显示:组1(干流5个采样点(GZ3、GX1~GX4)和支流理塘河(LTH1、LTH2))优势种有:短须裂腹鱼、长丝裂腹鱼、鲤(Cyprinuscarpio(Linnaeus))、金沙鲈鲤,常见种有鲇(SilurusasotusLinnaeus)、齐口裂腹鱼,鲤平均体重最大,3种裂腹鱼个体较小,平均体重均低于250 g;组2(卡拉往上中上游干流河段及支流鲜水河)优势种有:长丝裂腹鱼、短须裂腹鱼、四川裂腹鱼、齐口裂腹鱼,平均体重在200~310 g之间;组3(二级支流永兴河)优势种有:短须裂腹鱼、红尾荷马条鳅(Homatulavariegatus(SauvageetDabry))、短体荷马条鳅(Homatulapotanini(Günther)),常见种有云南盘鮈(Discogobioyunnanensis(Regan)),这4种鱼类个体均偏小,除短须裂腹鱼外,另3种鱼类平均体重在10 g以下;组4(金河、得石、雅砻江河口等流水河段)优势种有(Hemiculterleucisculus(Basilewsky))、短须裂腹鱼、鲤、圆口铜鱼,常见种有鳙(Aristichthysnobilis(Richardson))、凹尾拟鲿(Pseudobagrusemarginatus(Regan))、齐口裂腹鱼、细鳞裂腹鱼、长丝裂腹鱼,鳙个体最大,平均体重达1278.3 g,其次是鲤、细鳞裂腹鱼、短须裂腹鱼和圆口铜鱼,平均体重在400 g以上;组5(二滩和鳡鱼河等静缓流河段)优势种有翘嘴鲌(CulteralburnusBasilewsky)、尼罗罗非鱼、鳙、、鲫(Carassiusauratus(Linnaeus)),常见种有鲢(Hypophthalmichthysmolitrix(Valenciennes))、鲂(MegalobramamantschuricusBasilewsky)、鳜(Sinipercachuatsi(Basilewsky)),此聚类组鳙个体最大,其次为鲢;组6(霍曲河、卧龙寺沟中游2条支流河段)优势种有软刺裸裂尻鱼(SchizopygopsismalacanthusmalacanthusHerzenstein)和短须裂腹鱼,个体均偏小(表1)。
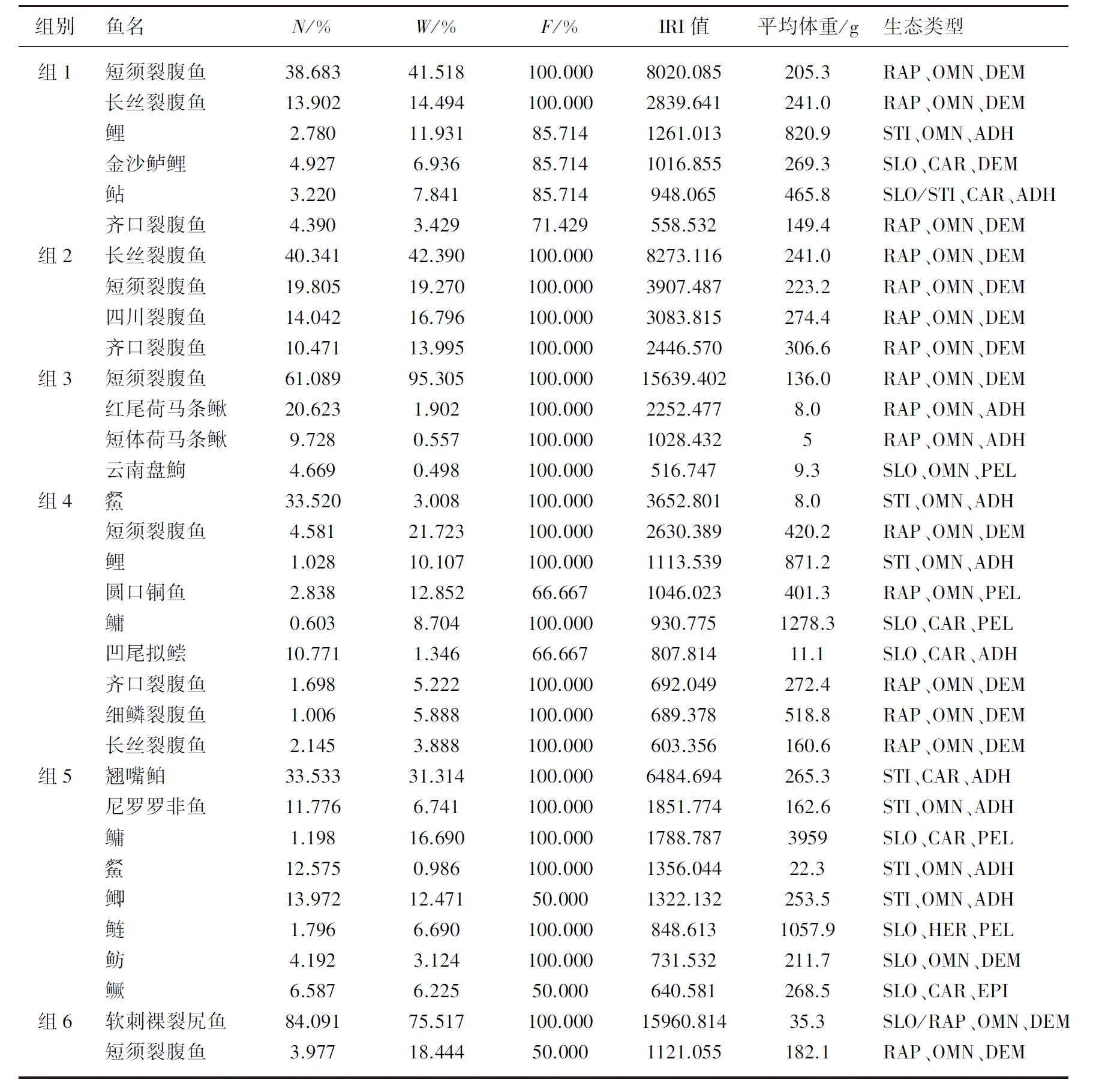
表1 Cluster聚类组优势种组成及生态类型*Tab.1 Composition of the dominant species in 6 cluster groups
对各组优势种的栖息类型、食性和产卵类型进行分析发现,各组优势种与鱼类生态类型有一定的关联性,如组2为卡拉往上中上游干流河段及支流鲜水河基本处于天然状态的河段,优势种和常见种均为急流型、杂食性鱼类,并产(黏)沉性卵;组5为二滩及鳡鱼河的静缓流河段,该组5种优势种均为静水型或缓流型鱼类,产卵类型多为黏性卵,与其他组流水性河段优势种生态类型差异较大。对6个聚类组优势种和常见种从栖息类型、食性、产卵类型上进行单因素方差分析,结果表明6组优势种在栖息类型(F=3.09,P<0.05)和产卵类型(F=2.846,P<0.05)上差异显著,仅在食性上差异不显著。
3 讨论
3.1 鱼类种类组成及其变化
2017-2022年调查期间通过监测和资料调研共获得雅砻江干支流鱼类98种,除去6种境外引入种和1种黄河特有种,还有91种鱼类,明显多于刘成汉[3]在1958-1959年期间调查和整理文献所获得的40种,可能原因与刘成汉仅对支流安宁河进行了实地调查有关,雅砻江干流的鱼类是通过查阅资料所得,因此整体种类数较少。吴江等[5]在1983-1984年开展了雅砻江渔业自然资源调查,共获得鱼类92种,与本调查种类数接近。邓其祥[4]在1968年、1980-1984年间对雅砻江干支流进行调查,并结合历史资料整理出鱼类种类89种,其中还包括泸沽湖和邛海等区域鱼类。张春光等经过实地调查和文献调研报道雅砻江流域分布土著鱼类72种[11],少于本文监测和调研的种类数。不同文献报道雅砻江流域鱼类种类数的差异可能与调查网具、调查强度等有很大关系。
本研究在下游干支流(干流及支流理塘河、鳡鱼河和永兴河)所获得的鱼类共87种,其中贝氏高原鳅(Triplophysableekeri(SauvageetDabry))、青石爬鮡、黄石爬鮡(EuchiloglaniskishinouyeiKimura)和虹鳟来自刘猛等[13]2018-2019年在大河湾的调查,其余83种为笔者调查所得,比杨青瑞等[9]报道的2007-2008年雅砻江下游75种鱼类多,可能与调查时长有关,本调查历时6年,比杨青瑞等开展的调查时间长4年,可能会影响一些偶见种如雅砻白鱼、裸体异鳔鳅鮀(Xenophysogobionudicorpa(HuangetZhang))、中华鳑鲏(RhodeussinensisGünther)和白甲鱼(Onychostomasima(SauvageetDabry))等的调查情况。邓其祥[7]报道1983-1984年间在雅砻江小金河口至雅砻江河口区域获得鱼类118种(亚种),剔除邛海、彝海和泸沽湖3个湖泊独有的7种鱼类和资料记载而未调查到的种类,还剩83种,为实际调查所获得的雅砻江下游干支流鱼类种类,比2017-2022年在下游干支流所获得的种类数稍少。2017-2022年新增的种类主要是飘鱼(PseudolaubucasinensisBleeker)、雅砻白鱼、贝氏(HemiculterbleekeriWarpachowski)、青鱼(Mylopharyngodonpiceus(Richardson))、团头鲂(MegalobramaamblycephalaYih)、鳜等适应静缓流生境的鱼类,可能与下游梯级电站建设形成水库导致生境改变、流速减小有关,且新出现种中外来物种很多,如大鳞鲃、散鳞镜鲤、斑点叉尾鮰、虹鳟、大口黑鲈、尼罗罗非鱼等。本次相对于邓其祥[7]未调查到的种类主要为圆筒吻鮈(RhinogobiocylindricusGünther)、四川白甲鱼(Onychostomaangustistomata(Fang))、短尾高原鳅(Triplophysabrevicauda(Herzenstein))、红唇薄鳅(Leptobotiarubrilabris(Dabry))、峨眉后平鳅(MetahomalopteraomeiensisChang)等适应急流生境的种类。
3.2 下游梯级电站建设对鱼类的影响
梯级电站建设和运行对鱼类资源的不利影响主要表现在阻隔和生境破碎化,及径流分配时空格局的改变引起自然水文情势改变,梯级开发使原有连续的河流生态系统被分隔成片断化的环境单元,造成生态景观的破碎及河流生态系统向水库-河流生态系统的转变[26]。大坝建设对洄游性鱼类产生阻隔,使其不能有效完成生殖和索饵等生活史过程,往往造成资源的严重下降,同时也影响鱼类的分布和不同水域群体间的交流。水库淹没导致流水性鱼类关键生境消失,一些原有的江河流水性鱼类种类数量减少甚至消失,而一些适应静水或缓流的鱼类及广适性鱼类逐步在群落中占据优势地位;同时,水库环境还可能提高外来物种入侵的可能性,加剧对土著种类生存竞争的压力[43]。
雅砻江下游已建成并运行锦屏一级、锦屏二级、官地、二滩和桐子林5级水电站,其中锦屏一级为年调节电站、二滩电站为季调节电站、官地和桐子林为日调节电站、锦屏二级为引水式电站。
对比分析锦屏一级、二级电站工程水域(锦屏一级库尾至锦屏二级厂房)鱼类种类变化,在水电站建设前(1990-1994年、1997年)共采集到31种鱼类[14],本调查该影响区鱼类(组1)共32种,种类数接近。在电站建设前主要鱼类有金沙鲈鲤、短须裂腹鱼、长丝裂腹鱼、四川裂腹鱼、长鳍吻鮈和长薄鳅等,本调查金沙鲈鲤和裂腹鱼属鱼类依旧为优势种类,新增优势种和常见种有鲤和鲇等静缓流适宜库区生境的鱼类,而长鳍吻鮈极少,长薄鳅在该河段未调查到。这种变化应与锦屏一级库区的形成给静缓流鱼类提供了适宜生境有关,因此鲤和鲇成为优势种、常见种,这与廖敏于2021年对锦屏一级库区进行的鱼类调查结果部分吻合,其优势种为、短须裂腹鱼、鲤和麦穗鱼(Pseudorasboraparva(TemmincketSchlegel))等静水型鱼类[15],本文聚类组1中的尾数比也高达6%。另锦屏二级减水河段的存在也给建设前的主要鱼类如金沙鲈鲤、裂腹鱼属鱼类等保留了适宜生境,因此这些鱼类依旧是该江段优势种,这与刘猛等的报道一致[13]。
二滩电站修建前1992-1994年、1996-1998年野外调查采集到鱼类61种[16],建库后2002年、2004年采集到57种[17-18],2007年采集到鱼类36种[19],本次在二滩库区及鱤鱼河采集到24种,鱼类种类数呈显著下降趋势,减少的种类有圆口铜鱼、长鳍吻鮈、圆筒吻鮈、泉水鱼(Pseudogyrinocheilusprocheilus(SauvageetDabry))、墨头鱼(GarraimberbaGarman)及高原鳅属、荷马条鳅属等适应急流生境的鱼类;新增的种类主要为翘嘴鲌、鳜、大眼鳜(SinipercakneriGarman)、长吻拟鲿(Pseudobagruslongirostris(Günther))等栖息静缓流生境的肉食性鱼类;其中,、团头鲂和尼罗罗非鱼为自2007年以来新增种类[19],且尼罗罗非鱼在本调查中数量激增,已成为本江段优势种类。
雅砻江下游梯级电站建设后,仅桐子林坝下至雅砻江江口长约15 km江段处于未开发状态,此江段鱼类与金河、得石采样点鱼类共同聚为组4(图4),金河处于二滩库尾流水江段,其上为官地电站,得石为桐子林坝上江段,官地和桐子林均为日调节电站,库容较小、库区存在一定流速,因此相对于锦屏一级和二滩等年调节、季调节等调节性强的电站,对流水性鱼类的影响相对较小[29,43],如本调查中圆口铜鱼亦是该聚类组优势种。
从以上分析结果可看出下游鱼类聚类结果与梯级电站建设有很大关系,电站的调节性能、减水河段、库尾流水河段长度等均影响着鱼类组成及其结构,因此在进行鱼类保护时要考虑电站的开发方式等。调节性能强的电站需适当控制静缓流生境鱼类的种群数量;对库尾流水江段,需采取措施保护此生境,特别对有洄游特性鱼类的江段,需建设相应过鱼设施使其顺利通过大坝,完成生殖和索饵洄游。
3.3 生物入侵及其影响
雅砻江下游大河湾以下流域属中亚热带湿润气候区,年平均气温18~21℃,河口一带近亚热带气候,具有罗非鱼(Oreochromisspp.)生长繁殖的自然环境[44-45]。下游梯级电站的修建运行造成整体流速减缓,江面变宽,库区水域加深,有机物和营养盐增加,库区浮游动植物等大幅增加,为罗非鱼和翘嘴鲌等静水型外来鱼类或水库常见种的大幅爆发提供适宜生境和饵料基础[26,46]。本调查显示,翘嘴鲌、尼罗罗非鱼等不仅在二滩库区、鳡鱼河等区域形成了自然种群,且成为Cluster聚类组5(二滩及鳡鱼河)中的优势种,IRI值高达6000和1800以上。二滩电站为雅砻江建成的第一座水电站,因此翘嘴鲌和罗非鱼等在二滩电站的种群规模和扩张状态需引起重视,应加强对下游流域外来物种的监测和控制,并采取措施降低外来鱼类的种群数量,避免发生大规模生物入侵现象。特别是Cluster聚类组4(二滩电站和桐子林电站建成后还存在的部分流水河段,如金河、得石江段),该组优势种有短须裂腹鱼、鲤和圆口铜鱼,其中短须裂腹鱼IRI值高达2600以上,因此特别要关注该组优势种组成及其变化,采取相应措施如定期捕捞限定种类等来避免罗非鱼等外来种的扩张,避免对下游流域裂腹鱼等土著鱼类和原有的生态系统造成巨大危害[45-46]。
3.4 鱼类群落结构及其变化
关于群落构建机制,目前存在着中性构建理论和生态位理论(包括竞争排斥学说和环境过滤学说)等,影响鱼类群落结构变化的因素较多,如可利用资源的数量、环境异质性和外界干扰作用等都会直接影响群落动态[47]。对2017-2022年雅砻江流域鱼类群落结构分析显示,在44.36%的相似性水平上鱼类群落可分成6组,这6组生境、水文条件相差较大,因此相似性水平较低,被分成不同的组别,这显示了环境过滤机制在雅砻江流域鱼类群落结构形成过程中的主导作用。Ward和Stanford在河流连续统概念的基础上,基于河流生态系统中普遍存在的人为干扰(如水利建设)进一步提出了河流“序列不连续体概念(serial discontinuity concept)”,该概念较为详尽地描述了人类活动对河流中的环境因素与生物成分及其生态过程的影响[48]。此外,水利建设也引起了栖息地同质化现象,即降低了环境异质性,正如牛红玉等[49]提出的外界干扰(本研究中的大坝修建)等也作为一种环境过滤器,对群落结构产生着影响,由此造成河流区域鱼类区系的同质化[50-51],这也部分解释了中上游干、支流基本处于天然状态的区域中鱼类群落结构与二滩电站影响区至河口段差异较大的现象。
另有研究表明,河道治理[52]和水利建设[53-54]等人类活动[55]改变了局域栖息地条件和水流流态,从而改变了河流中鱼类等水生生物类群的纵向梯度格局。大量研究显示水利工程通过阻隔鱼类的洄游路线[52]、坝上形成水库降低上游流速[56]、降低下游水温[57]和改变水流流态[58]等途径对鱼类的分布和数量产生深远影响。Petts[59]总结了水利工程的三重生态学效应:首先,改变水质、沉积作用的速率和水流流态;其次,改变河床的横截面特征、河床沉积物的移动性能和初级生产力;最后,导致大型底栖无脊椎动物和鱼类等生物群落的改变。本调查所形成的鱼类群落结构聚类结果表明,库区及库区支流聚成一支(组5),各库尾段和河口段流水段聚成一支(组4),主要原因是电站的运行对流速、水温和水流流态等上述因素的改变所导致的库区静水和坝下流水生境差异较大,且库区沉积物等的变化导致初级生产力的改变从而影响库区、库尾和坝下等不同区段浮游和底栖生物类群,进而影响鱼类的饵料供应,因此,库区和坝下鱼类群落相似性低。
一条河流从源头至河口的纵向梯度变化过程可以分成侵蚀、转化和沉积3个过程。由于存在一定的坡度,众多支流逐级合并汇入干流,溪流大小、流量和流域面积均逐级增大,且一系列生态环境因子往往呈现出显著的空间梯度变化。如上中下游的坡度、流速、河床基质、深度、河宽、水温和海拔高度等生态环境因子和水文环境特征均体现出极高的空间异质性与显著的周期性[60-61]。2017-2022年雅砻江鱼类形成的6个聚类组海拔高度和河流等级存在的显著性差异也说明空间异质性造成了鱼类群落结构差异。对组1内部各河段更细致的分类,发现更小的分支也是沿着海拔高度与河流等级进行排列的,如海拔上接近的锦屏大河湾段(GX1~GX4)先聚在一起,再分别与支流理塘河(LTH1和LTH2)的两个采样点鱼类以及海拔稍高的卡拉段(GZ3)相聚。对雅砻江流域从源头至河口鱼类群落随纵向梯度变化的探讨,还需结合更多生态环境因子进行长时间监测数据来进行,并对下游梯级电站建成后与中上游情况进行对比,可以更深入探讨人类活动的影响情况,建议开展雅砻江流域鱼类资源的连续监测,为整个流域鱼类群落结构的空间异质性、周期性变化及梯级电站影响等分析提供基础,并为生物多样性保护提供参考数据。
4 附录
附表Ⅰ、Ⅱ见电子版(DOI: 10.18307/2024.0134)。
猜你喜欢
节能与环保(2022年3期)2022-11-24
水泵技术(2022年2期)2022-06-16
水泵技术(2021年4期)2021-11-20
水产科学(2020年5期)2020-09-23
高原山地气象研究(2020年1期)2020-06-24
新农民(2019年2期)2019-02-20
当代水产(2018年8期)2018-11-02
水利科技与经济(2016年11期)2016-04-22
大众考古(2014年6期)2014-06-26
长江大学学报(自科版)(2013年33期)2013-03-11
