
植物功能性状系统发育保守性的类群和地理分异研究
——以中国被子植物最大株高为例
2024-01-23 05:35邢冰冰毛岭峰
南京林业大学学报(自然科学版) 2024年1期
邢冰冰,李 垚,毛岭峰
(南京林业大学生态与环境学院,江苏 南京 210037)
植物功能性状(functional trait)是指植物体具有的与其定植、存活、生长以及死亡密切相关的一系列核心植物属性(core plant traits)。这些属性能够显著地影响生态系统功能,并能够反映植物对环境变化的响应[1-3]。与植物种类和数量相比,植物功能性状可以更好地反映生态系统功能,被广泛地应用于种群、群落和生态系统尺度的研究中[3-5]。如近期研究发现,植物功能性状多样性比物种多样性对环境胁迫或扰动更敏感,更能指示生态系统功能的变化[6-7]。植物功能性状也直接反映了植物适应环境和参与生物环境构建的过程[4,8]。不同环境条件下的植物往往具有规律性变化的功能性状[8-9],如叶片和树干的功能性状随环境梯度呈现明显的地理格局[10-11]。
根据物种进化过程中的性状保守性假说,亲缘关系较近的物种往往比亲缘关系较远的物种具有更为接近的功能性状,即功能性状具有系统发育保守性(phylogenetic conservatism),或者说,功能性状会在进化过程中呈现出明显的系统发育信号(phylogenetic signal)[12-13]。目前,越来越多的研究指出,由于物种的功能性状和生态属性等具有系统发育保守性,其生态适应性也具有系统发育保守性[14-15]。然而,也有反例存在,如有研究发现佛罗里达地区栎树的生态位重叠和生境选择并不具有系统发育保守性[16]。
在众多的研究中,物种的系统发育关系(或系统发育距离)常用来替代其生态属性或环境适应能力的相似性,进而用于检验大尺度或者群落构建(community assembly)过程中的生态学基本理论和规律[17-18]。例如,前期学者利用群落物种的系统发育关系结构来解析竞争排斥(competitive exclusion)、环境过滤(environmental filtering)和随机过程(stochasticity)在群落构建中的作用[16,19-20]。上述研究均建立在竞争-生态位相似性假说(competition-niche similarity hypothesis)的基础上[21],即假设生态相似度高的物种对于环境资源的竞争程度要大于生态相似度低的物种,继而不易共存。同时,研究也发现,由于植物功能性状具有系统发育保守性,这种生态相似性也可以用功能性状来代替[22]。然而,这一假设前提面临诸多的挑战,如研究人员发现在很多情况下一些关键植物功能性状不具有系统发育保守性[23-24]。在这种情况下,其生态学属性也可能不具有系统发育保守性。这将导致系统发育关系不能应用于替代功能性状所反映的生态学关系,成为学科发展的一个争论点。此外,由于植物功能性状在生态学研究中极为重要,许多研究需要获取大量的植物功能性状数据,但是受限于采样困难,很多功能性状并不能完全被获取[25]。新的系统发育填补(phylogenetic imputation)方法的发展使得采用数学模型推断性状缺失值成为可能[23-24]。然而,运用这种方法的前提条件依旧是功能性状具有系统发育保守性[12,26]。但是,植物的功能性状保守性在什么类群、什么区域存在,依旧有待基于大数据的系统分析来回答。
被子植物的最大株高是其生态策略的核心部分,它与植物的寿命、种子质量和成熟时间等都密切相关[27],也是决定植物竞争光照能力的关键因素之一[28]。株高在系统发育关系替代功能性状生态属性的研究中常被用来代表物种间的竞争作用[29],在推断功能性状缺失值的研究中也应用广泛[11,24]。当前已有研究表明,中国被子植物花期、果期等物候性状在区域尺度上具有系统发育保守性,且呈现一定的地理格局[30-31],但是最大株高的系统发育保守性及其地理格局尚未明确,仅在样地水平有少数研究[15,32]。另外,最大株高的变异性随纬度而变化[33],很大程度上取决于区域内共存物种的种间竞争,竞争和地点间的气候差异往往都只能解释最大株高变异的一半[33-34]。因此,被子植物最大株高的系统发育保守性在不同的地域可能存在较大的差异。例如,在海南尖峰岭热带雨林地区的乔木层中,最大树高的系统发育保守性不显著[15];在对美洲东部和欧洲地区植物最大株高缺失性状的推断研究中,Swenson等[24]将其对树高的不理想推断结果归咎于当地物种株高的系统发育保守性较弱。
本研究拟以中国被子植物的最大株高为研究对象,利用20 295种被子植物的株高、生长型和地理分布数据,检测不同生长型、不同类群和不同植被区域植物最大株高的系统发育保守性,并探讨以下3个问题:①草本、木本植物以及不同进化分支的被子植物最大株高的系统发育保守性是否存在差异?②被子植物最大株高的系统发育信号呈现怎样的地理格局?③被子植物最大株高的系统发育信号与温度、降水量等环境因子具有怎样的关联?
1 材料与方法
1.1 被子植物最大株高、地理分布数据的收集
被子植物最大株高数据来自基于《中国植物志》及各级地方植物志描述建立的植物高度性状数据库。植物学名依据The Plant List物种名录1.1版本(www.theplantlist.org)进行修正。最终,共获取了20 295种被子植物的最大株高数据。被子植物地理分布数据来自于中国维管植物物种分布数据库[35-36],描述了每种植物在100 km × 100 km栅格分布情况。被子植物生长型数据依据《中国植物志》和《Flora of China》的记录收集整理,包括草本和木本植物两种生长型。本研究共涵盖草本植物13 205种、木本植物7 090种。
1.2 进化树构建与系统发育信号计算
本研究所涉及的被子植物进化树基于全球种子植物系统发育骨架树(GBOTB)[37]构建。该骨架树包含了79 881个种级及种级以下的分类单元,是目前最为完整的被子植物进化树。利用R软件的V.PhyloMaker包[38]中的phylo.maker函数匹配和插入所有20 295种中国被子植物。对于GBOTB中已有的物种,直接匹配到骨架树;对于GBOTB中没有的物种,将其随机插入对应的属或科下。为减小随机插入对结果的影响,独立生成3棵进化树用于后续分析。
利用零模型(null model)检验最大株高是否具有显著的系统发育信号,即检测系统发育关系较近的植物是否具有更为相近的高度值。具体来说,该方法随机分配物种性状值于系统发育树的各个节点,然后通过比较性状在进化树上的实际分布和基于进化随机的布朗运动(Brownian motion)状态来判断性状的系统发育保守性[39-40]。系统发育信号采用基于布朗运动进化模型的Pagel’sλ参数度量[41],0<λ<1。当λ= 1时,表明性状进化符合布朗运动模型;当λ= 0时,表明性状进化与系统发育关系相互独立[41]。前期研究表明,Pagel’sλ对末端多分支结构不敏感,因而用其估算多分支树的系统发育信号的不确定性较低[42]。上述分析采用R软件的phytools包[43]中的phylosig函数完成,零模型运算次数为1 000次。由于不同生长型的植物具有不同的适应策略和进化历史,分别计算全部植物、草本植物、木本植物最大株高的系统发育信号并检验其显著性。同时,为了比较不同类群系统发育保守性的差异,根据APG IV分类系统[44],对树高数据中包含500个物种以上的13个目[禾本目(Poales)、天门冬目(Asparagales)、毛茛目(Ranunculales)、豆目(Fabales)、蔷薇目(Eosales)、金虎尾目(Malpighiales)、虎耳草目(Saxifragales)、石竹目(Caryophyllales)、杜鹃花目(Ericales)、菊目(Asterales)、龙胆目(Gentianales)、伞形目(Apiales)和唇形目(Lamiales)]的最大株高系统发育信号进行单独检验。
1.3 被子植物最大株高系统发育保守性的地理分异分析
为了解析最大株高系统发育保守性的地理格局,在全国尺度上分别计算每个100 km × 100 km网格内被子植物最大株高的系统发育信号,最终全国尺度上使用的网格数为943个。为了更直观地体现不同植被区域系统发育信号的差异,根据中国植被地理区划,将中国陆域划分为热带季雨林雨林区、亚热带常绿阔叶林区、暖温带落叶阔叶林区、温带针叶阔叶混交林区、寒温带针叶林区、温带草原区、温带荒漠区、青藏高原高寒植被区共8个区域。植被地理分区数据来源自中国科学院资源环境科学与数据中心网站(https://www.resdc.cn/)。
为了探究最大株高系统发育保守性地理格局的成因,分别使用局部加权回归[45]和多项式回归模型分析系统发育信号与纬度、气候因子的关系。气候因子包括年平均气温、最冷季平均气温、年降水量和最干月降水量,代表了每个网格的水热条件及其极端状况。气候数据下载自WorldClim 1.4数据库[46],采用ArcGIS 10.2计算每个网格内栅格值的平均值。所有模型均在R中构建,其中局部加权回归模型使用lowess函数完成,平滑跨度(smoother span)f选择0.3。分别对全部植物、草本植物和木本植物进行相关地理分异分析。
2 结果与分析
2.1 中国被子植物最大株高系统发育保守性的类群分异
本研究中共有8 137种被子植物直接匹配到了GBOTB骨架树,有12 158种被子植物随机插入到了对应的属或科下。利用3棵随机插入生成的进化树计算最大株高的系统发育信号,结果表明,全部植物、草本植物和木本植物的(Pagel’sλ)平均值分别为0.893(P<0.001,数量(n) = 20 295)、0.725(P< 0.001,n= 13 205)和0.813(P<0.001,n= 7 090)。草本植物最大株高的系统发育保守性程度略低于木本植物。
物种数大于500种的13个目植物最大株高的系统发育信号均具显著性(P<0.001),但不同类群间系统发育信号的强弱具有明显的分异(图1)。

图1 包含500种以上13个目的被子植物最大株高的系统发育信号Fig. 1 Phylogenetic signals of maximum plant height for Chinese angiosperms in 13 orders with more than 500 species
在这13个目中,蔷薇目的Pagel’sλ值最低,为0.612;豆目最高,达到了0.995。石竹目、天门冬目、唇形目、伞形目、虎耳草目的Pagel’sλ值都高于0.9,说明在这些类群内,最大株高的进化是高度保守的,具有强烈的系统发育信号。
2.2 中国被子植物最大株高系统发育保守性的地理分异
从地理空间来看,对于全部被子植物来说,中国东南部和东北部的最大株高的系统发育信号相对较低,而青藏高原、内蒙古高原、山东半岛地区的系统发育信号相对较强。对于木本植物来说,大部分区域的系统发育信号都非常强烈,青藏高原和中国东北部部分地区的系统发育信号非常低。对于草本植物来说,整个新疆地区的系统发育信号都非常强烈,而在青藏高原大部分区域、内蒙古高原东部和中国东北部地区的系统发育信号较低,但青藏高原东南部(察隅、墨脱地区)的系统发育保守性较强。
二次多项式回归结果表明,在纬度梯度上,所有被子植物(图2a,R2= 0.412,P<0.001)和木本植物(图2c,R2= 0.058,P<0.001)最大株高的系统发育信号均呈明显的单峰分布,说明这些植物类群最大株高的系统发育保守性在中纬度地区较强,而在低纬度和高纬度地区较弱。然而,对于草本植物来说,局部加权回归和多项式回归模拟结果均表明其最大株高的系统发育信号不呈明显的单峰分布(图2b),其在纬度梯度上虽然呈现显著的三次函数关系,但模型拟合程度较低(R2= 0.019,P< 0.001)。该结果表明草本植物最大株高系统发育保守性程度的纬度梯度规律不明显。

红色线和蓝色线分别表示局部加权回归和多项式回归结果。下同。The red and blue lines represent the results of locally weighted regression and polynomial regression, respectively. The same below. 图2 中国被子植物最大株高的系统发育信号与纬度的回归关系Fig. 2 Regressions between phylogenetic signals of maximum plant height and latitude for Chinese angiosperms
2.3 被子植物最大株高的系统发育保守性与气候因素的关系
二次多项式回归结果表明,对于被子植物整体及木本植物而言,最大株高的系统发育信号与年平均气温(被子植物整体R2= 0.196,P<0.001;木本植物R2= 0.060,P<0.001)、最冷季平均气温(被子植物整体R2= 0.334,P<0.001;木本植物R2= 0.078,P< 0.001)均呈明显的单峰曲线关系(图3)。对于草本植物而言,随着年均气温、最冷季平均气温的增加,最大株高的系统发育信号呈现先降低、后上升、再降低的趋势(图3);三次多项式回归结果虽然显著,但拟合程度较低(年均气温R2= 0.097,P<0.001;最冷季平均气温R2= 0.074,P<0.001)。总体来看,被子植物最大株高的系统发育信号与年均气温、最冷季平均气温具有相似的关系,但对于被子植物整体和木本植物来说,最冷季平均气温的模型解释能力相对较强。
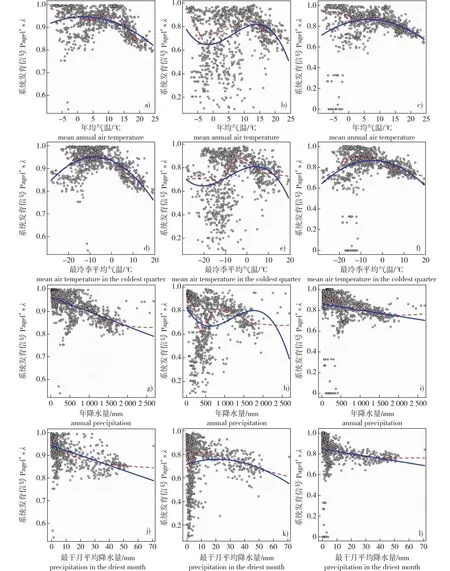
a、d、g、j. 被子植物整体all angiosperms;b、e、h、k. 草本植物herbaceous plants;c、f、i、l. 木本植物woody plants。图3 中国被子植物最大株高的系统发育信号与年平均气温、最冷季平均气温、年降水量、最干月降水量的回归关系Fig. 3 Regressions between phylogenetic signals of maximum plant height and mean annual air temperature, mean air temperature in the coldest quarter, annuel precipitation and precipitation in the driest month for Chinese angiosperms
普通线性回归结果表明,对于所有被子植物和木本植物而言,最大株高的系统发育信号与年降水量(被子植物整体R2= 0.339,P<0.001;木本植物R2= 0.037,P<0.001)、最干月降水量(被子植物整体R2= 0.250,P<0.001;木本植物R2= 0.035,P<0.001)均呈显著的负相关(图3)。对于草本植物而言,随着年降水量的增加,最大株高的系统发育信号呈现先降低、后上升、再降低的趋势(三次多项式回归:R2= 0.067,P<0.001),但其与最干月降水量的关系不显著(三次多项式回归:R2= 0.006,P= 0.147)(图3)。总体来看,被子植物株高的系统发育信号与年降水量、最干月降水量具有相似的关系,但对于所有物种和木本植物来说,年降水量的模型解释能力相对较强。
3 讨 论
株高是植物生态策略的核心部分,它与植物寿命、种子质量和成熟时间密切相关,是物种争夺光能力的主要决定因素[28]。本研究发现,中国被子植物最大株高的系统发育保守性较强(Pagel’sλ= 0.893,P<0.001),表明系统发育关系较近的被子植物倾向于具有更为接近的株高[34]。然而,值得注意的是,最大株高系统发育保守性的强度在不同进化分支、不同植被区域并非一成不变,而是存在明显的类群、地理差异。如豆目株高的系统发育信号值为蔷薇目的1.6倍;中国东南部和东北部被子植物株高的系统发育信号明显较青藏高原、内蒙古高原等地区更高。上述结果表明,在用系统发育关系替代株高所反映的植物生态属性或环境适应能力时,或者用系统发育填补的方法推断株高的缺失值时,应充分考虑所研究类群或所关注地理区域之间的系统发育保守性差异。当所研究的进化分支或所关注的地理区域的被子植物株高系统发育保守性较低时,应谨慎地估计所存在的误差和解释所得到的结果。
本研究发现木本、草本植物最大株高的系统发育保守性强度不一致。木本植物或者草本植物最大株高的系统发育信号低于被子植物整体,且草本植物的系统发育信号低于木本植物。这一结果说明木本植物或者草本植物最大株高的系统发育保守性弱于被子植物整体,而木本植物株高的系统发育保守性较草本植物更强。已有研究表明,在被子植物中,木本植物与草本植物的功能性状在应对环境压力时的表现不同[47-48]。由于木本植物谱系比草本植物谱系的进化速率慢,生长形式和世代时间会对其功能性状的演化起到明显的制约作用[49],其最大株高受极端环境的影响较大,在不同的生态位下往往会出现较大的差异[50-51],但在相似的环境中物种之间可能彼此相近[52]。此外,木本植物在迁移到可用的生态位空间前,其性状可能不会偏离祖先的气候耐受性太远[49,52]。相对于木本植物,草本植物的物种形成和灭绝速率可能高于木本植物[53],在演化过程中株高的多样性比木本植物高;另外,草本植物比木本植物具有更广的生态位分布,更能适应极端气候[49,52]。因此在大尺度下,木本植物的最大株高可能更容易表现出相关物种之间彼此相近的趋势。所以,单独使用草本植物或木本植物的系统发育关系来替代其生态属性等关系所得出的结论可能会出现差别。
本研究发现被子植物最大株高的系统发育保守性具有明显的地理格局,沿环境梯度呈规律性变化。功能性状的形成是表型稳定性(内部选择)和对当前环境条件适应(外部选择)之间的平衡。外部选择依赖于环境,内部选择依赖于物种自身的遗传,也就是进化的约束,不管生物处在什么环境下,进化约束在地理上是不会发生改变的[54]。而功能性状的系统发育信号即保守性表示物种表现出相同功能性状的趋势,既会受到进化约束的影响,也会受到外部环境因素的影响。对系统发育信号和环境因子的相关性分析表明,所有植物和木本植物的最大株高系统发育保守性在纬度梯度上呈明显的单峰分布,或者说,中纬度地区的系统发育保守性强于高、低纬度地区。这一地理格局与中国被子植物花期、果期、果实发育期长度等性状的系统发育保守性地理格局相似[30-31]。由于纬度与最冷季均温呈显著的正相关,本研究还发现最大株高的系统发育保守性在最冷季均温梯度上也呈明显的单峰分布。上述结果表明,在高纬度或严寒地区的恶劣条件下,局部适应或定向选择的作用可能强于系统发育关系的约束作用。换言之,无论物种间的亲缘关系如何,它们均生长在同样严酷的环境下,极端气候条件将同时限制植物的营养和生殖生长。趋同进化作用将成为决定植物生态属性或功能性状演化方向的主要因素[28,55],这将导致包括株高、花果期等功能性状在高纬度或严寒地区的系统发育保守性降低。本研究发现靠近热带地区的被子植物最大株高的系统发育保守性较温带地区弱,这一规律与中国19 631种被子植物开花时间的系统发育信号的纬度格局一致[32]。这种纬度模式可以用系统发育生态位保守性假说来解释[55]。与热带地区相比,温带地区植物在系统发育上更趋向于聚集,分支年龄较低[36],这可能使得株高等功能性状在温带地区更为保守。此外,靠近热带的地区为植物提供了更多的适宜生境[56],优越的环境条件使得株高的外部限制因子较少,在漫长的进化历程中,其性状值可以在较宽的范围内变化[31],这也可能导致低纬度地区被子植物最大株高的系统发育保守性较低。
猜你喜欢
河北环境工程学院学报(2021年1期)2021-03-19
世界科学技术-中医药现代化(2020年2期)2020-07-25
农村百事通(2019年17期)2019-10-08
现代园艺(2018年1期)2018-03-15
科学种养(2017年6期)2017-06-13
临床医药文献杂志(电子版)(2017年11期)2017-05-17
现代农业科技(2017年1期)2017-03-06
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
中学生物学(2016年8期)2016-01-18
