
色木槭与元宝槭表型性状分类学研究
2024-01-30 14:31任婷婷鄂思彤陈建华潘晶晶董胜君
经济林研究 2023年3期
关键词:分类
任婷婷 鄂思彤 陈建华 潘晶晶 董胜君
摘 要:【目的】探寻色木槭与元宝槭的分类性状,以期为两种槭树的分类鉴定提供科学依据。【方法】以辽宁省色木槭与元宝槭8 个天然种源共47 个样本为研究对象,采用变异分析、相关分析、主成分分析、聚类分析等方法,对其叶片和果实的25 个表型性状进行研究。【结果】色木槭与元宝槭表型性状的平均变异系数为15.22%,平均Shannon-Wiener 多样性指数为1.98,各种源变异水平较高,表型多样性丰富,其中叶片性状的变异程度大于果实性状。各样本叶片和果实性状受经度和海拔的影响显著,辽东地区以色木槭分布为主,辽西地区以元宝槭分布为主。通过R 型聚类分析筛选出23 个主要性状进行主成分分析,其中第一主成分代表叶裂宽度性状,第二主成分代表叶裂长度和果实长度性状,主成分向量散点图可将各样本按地理位置的东西分布分类,结合逐步回归分析,叶片长等9 个性状可作为评价两种槭树的主要性状。Q 型聚类分析将47 个样本分为3 类,果长、张开角和翅果比在组间差异最大,其分类结果与相关分析和主成分分析结果一致。【结论】叶片长、叶片宽、左下裂片长、叶基部左侧宽度、翅长、果长、着生痕长、果厚、张开角和翅果比可作为区分色木槭与元宝槭的分类依据,其中果长、张开角、翅果比是两种槭树表型的核心分类性状,具有果长≤ 10 mm、张开角≥ 114°、翅果比≥ 1.6 的形态特征可认定为色木槭,果长≥ 14 mm、张开角≤ 84°、翅果比≤ 0.9 的形态特征可认定为元宝槭。本研究结果可为槭属植物表型性状的种间鉴定和分类体系构建提供标准和依据。
关键词:色木槭;元宝槭;表型性状;分类
中图分类号:S603.7 文献标志码:A 文章编号:1003—8981(2023)03—0024—13
槭树科Aceraceae 槭属Acer L. 植物在全世界约有200 种,主要分布于北温带地区,我国是世界上槭属植物最丰富的国家,约有140 种[1]。槭属主要为落叶或常绿乔木、灌木,其木材坚硬,叶型秀丽,果形奇特,枝叶可入药,果实富含油脂、黄酮、蛋白质等有效成分,是集营养、药用和经济价值为一体的植物[2],广泛应用于食品、建筑及工业等诸多领域[3]。
色木槭Acer mono 又名五角枫,其树形优美,深秋叶色丰富[4],木材坚硬,是优良的园林景观树种和建筑材料;种仁富含蛋白质,可榨油[2],同时也是良好的饲料来源;叶片、种皮可作药用[5]。因其含水量大、含油量小且不易燃,可作为防火树种,但目前色木槭种群数量下降且分布零散,在《中国物种红色名录》中被列为“近危种”[2]。元宝槭Acer truncatum 又名元宝枫,其树姿优美,木材纹理美观,具有很高的观赏价值;叶片中黄酮类化合物含量丰富,可制作茶饮[6];种仁油富含人体必需的脂肪酸,是一种优质食用油[7]。色木槭与元宝槭同为槭树科槭属桐状槭组乔木[1],均为单叶,叶常5 裂,翅较果长或近等长,主要分布于吉林、辽宁、内蒙古各省(区)[8],其別名均为五角枫[9],但它们却是两个不同的分类种。因其形态特征相近,别名相同,在应用时往往混为一谈,在已出版的论文和专业书籍中也存在此类问题[10],因此准确区分两种槭树十分必要。从植物分类学角度来看,色木槭与元宝槭的分类地位仍有争议,分类界限不够清晰,需要进一步研究如何准确区分这两种槭属植物。
表型性状是植物遗传多样性的直观体现,是树种鉴别、保护及新品种选育的重要依据[11-12],其受植物内源基因与外在环境互相影响而发生改变[13-14],形态学分类法是开展植物表型研究最主要的手段,依据植物外部表现形态的差异性或相似性,既简单直观又形象地鉴别植物种类[15]。长期以来,植物的表型分类一直是国内外学者的研究焦点,在橄榄[16]、樱属[17]、细裂槭[18]、木芙蓉[19]、软枣猕猴桃[20] 等树种中广泛应用。本文针对色木槭与元宝槭分类鉴定标准不明确、应用易混淆等问题,以辽宁省8 个天然种源的47 个色木槭和元宝槭样本为研究对象,对其叶片和果实表型性状进行调查,分析不同种源和个体间的多样性及变异程度,同时了解这两种槭树在辽宁地区的天然分布,以期为色木槭和元宝槭的分类鉴定和资源利用提供科学依据。
1 材料与方法
1.1 试验材料
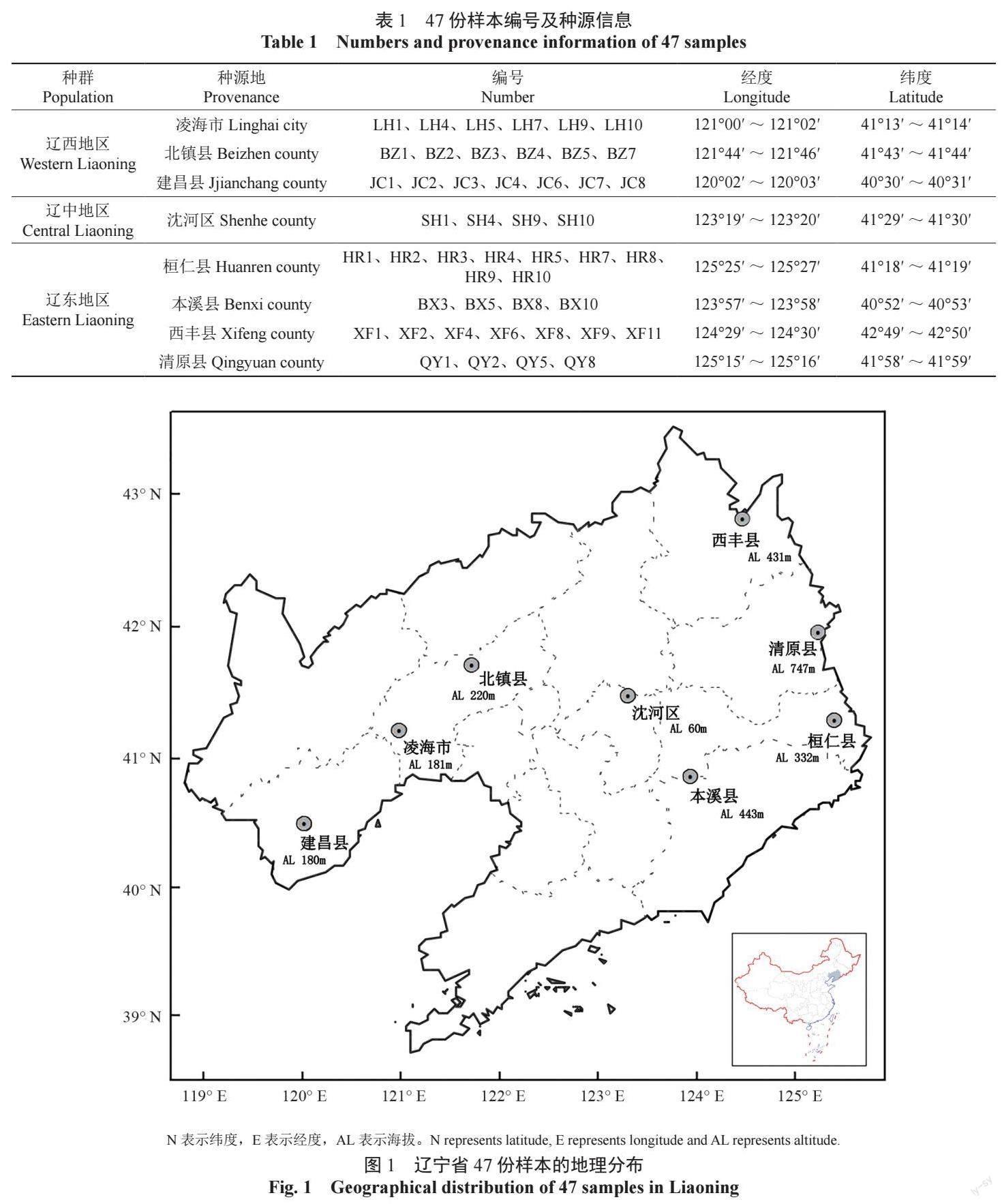
试验材料为辽宁省8 个天然种源的色木槭与元宝槭样本,于2019 年7—8 月选取生长健壮的成年树共47 株(表1),其中锦州市凌海市6 株、北镇市6 株,葫芦岛市建昌县7 株,沈阳市沈河区4 株,本溪市桓仁县9 株、本溪县4 株,铁岭市西丰县7 株,抚顺市清原县4 株。利用GPS 定位记录各样本的地理位置信息,具体地理分布见图1。
1.2 表型性状的测定
采集色木槭与元宝槭的叶片和果实,其中叶片每株6 片,果实每株10 枚,表型性状参照乔谦[5]和姬志峰[21] 的方法进行测定,选取样本HR4 和JC7 作为标准样本,叶片性状见图2,果实性状见图3,每个性状6 次重复,测量方法如下:
叶片:使用游标卡尺测量叶片长、叶片宽、叶柄长、叶柄基部宽、中裂长、中裂宽、左上裂片长、左上裂片宽、左下裂片长、左下裂片宽、叶脉到第二个夹角处距离、叶基部左侧宽度和基部中心到最下方的高度,使用量角器测量左侧角1、左侧角2 和叶片基部角,叶裂深= 叶基部左侧宽度- 叶脉到第二个夹角处距离。
果实:使用游标卡尺测量翅长、果长、着生痕长、果厚和翅宽,使用量角器测量张开角和连接角,翅果比= 翅长/ 果长。
1.3 数据分析
应用Microsoft Excel 2021 软件计算表型性状的均值、Shannon-Wiener 多样性指数(H′)、变异系数和主成分综合得分。采用ArcGIS 10.6 软件制作种源地理分布图。利用SPSS 22.0 软件对原始数据标准化后,采用组间链接以Pearson 相关系数为度量标准,进行R 型聚类分析,采用Ward 法以平方Euclidean 距离为遗传距离,进行Q 型聚类分析。以主成分1 向量、主成分2 向量分别作为横、纵坐标[20],采用Origin 2021 软件制作主成分散点图。
2 结果与分析
2.1 表型性状变异分析
由表2 可知,25 个表型性状的平均变异系数为15.22%,各种源平均变异系数在13.19% ~17.96% 之间。其中凌海市样本叶片基部中心到最下方高度的变异系数最大(178.40%),连接角最小(4.30%);北镇县样本叶片基部中心到最下方高度的变异系数最大(103.50%),叶片长最小(7.70%);建昌县样本叶片基部中心到最下方高度的变异系数最大(49.90%),果长最小(3.70%);沈河区样本叶片基部中心到最下方高度的变异系数最大(84.20%),左侧角2 最小(5.60%);桓仁县样本叶片基部中心到最下方高度的变异系数最大(45.70%),叶裂深最小(5.40%);本溪县样本叶片基部中心到最下方高度的变异系数最大(72.50%),左侧角1 最小(2.20%);西丰县样本果实张开角的变异系数最大(45.50%),左侧角2 最小(3.50%);清原县样本叶片基部中心到最下方高度的变异系数最大(48.70%),果厚最小(2.10%)。除西丰县外,基部中心到最下方高度性状在其他种源中的变异系数均最大,说明该性状的变异程度最大。不同种源表型性状的平均变异系数由大到小依次为:凌海市(18.00%)>西丰县(16.40%)>北镇县(16.20%)>建昌县(15.50%)>沈河区(15.30%)>本溪县(13.70%)>桓仁县(13.40%)>清原县(13.20%),凌海市样本表型性状的变异程度最丰富。
25 个表型性状的Shannon-Wiener 多样性指数变化范围在1.85 ~ 2.10 之间,均值为1.98,叶片和果实性状均具有丰富的多样性。其中遗传多样性较高(H′ ≥ 2.00)的叶片性状有左上裂片宽(2.10)、中裂宽(2.04)、左下裂片宽(2.03)、中裂长(2.02)、左上裂片长(2.02)、叶柄基部宽(2.01)、左下裂片长(2.01);果实性状有翅宽(2.05)、果厚(2.03)、着生痕长(2.02)、翅长(2.01)、张开角(2.00)。
2.2 表型性状与地理因子的相关性分析
将25 个表型性状与经纬度、海拔进行相关性分析发现(图4),所有性状与纬度均无相关关系。其中中裂宽、左上裂片宽、翅果比与经度呈极显著(P < 0.01)正相关,叶基部左侧宽度、左侧角1 与经度呈显著(P < 0.05)正相关,果长与经度呈极显著(P < 0.01)负相关,着生痕长、果厚与经度呈显著(P < 0.05)负相关;翅果比与海拔呈极显著(P < 0.01)正相关,中裂长、果长、着生痕长、果厚与海拔呈极显著(P <0.01)负相关,左上裂片长、连接角与海拔呈显著(P < 0.05)负相关。
共有11 个表型性状与经度、海拔具有显著或极显著的相关性(图4),這些性状与经度、海拔回归方程式的相关系数均在0.57 以上,显著性系数均低于0.05,回归方程有意义且拟合效果较好(表3 ~ 4)。随经度的增加,中裂宽、左上裂片宽、叶基部左侧宽度、左侧角1、翅果比增大,果长、着生痕长和果厚减小;随海拔的增高,翅果比增大,中裂长、左上裂片长、果长、着生痕长、果厚、连接角减小。由于辽西地区各种源的海拔与经度均低于辽东地区的各种源(图1),根据上述规律可知,辽西地区各种源的叶片形态窄长,果实形态较长且厚,翅果比小;辽东地区各种源的叶片形态短宽,果实形态较小且翅果比大。综上,辽东地区各种源样本多为色木槭形态特征,辽西地区各种源样本多为元宝槭形态特征。
2.3 表型性状的筛选
2.3.1 R 型聚类分析
对色木槭与元宝槭47 个样本的25 个性状进行R 型聚类分析,由图5 可知,中裂宽与左上裂片宽(r=0.934)、中裂长与左上裂片长(r=0.913)、果长与着生痕长(r=0.857)的相关程度较高,说明其两两之间差异不大,每组可选取其中一个性状作为分类依据。根据研究结果去除左上裂片长、左上裂片宽2 个性状,果长与着生痕长2 个性状虽相关性较大,但它们可以从不同方面反映出果实的形态,因此保留。其他性状彼此间相关性较小,可以在植物分类中起到各自的作用,最终选择23个表型性状用于后续分析。
2.3.2 主成分分析
利用筛选后的23 个性状进行主成分分析,由表5 可知,提取特征值大于1 的主成分有6 个,其贡献率分别为28.33%、19.60%、11.82%、9.24%、7.25%、5.42%,累计贡献率为81.65%。其中叶基部左侧宽度(0.37)、叶片长(0.34)、叶片宽(0.33)、叶脉到第二个夹角处距离(0.33)、中裂宽(0.31)、叶裂深(0.27)、叶柄长(0.24)在主成分1 中的特征向量绝对值较大,主要反映叶裂的宽窄;中裂长(0.39)、翅果比(-0.34)、果长(0.33)、左侧角1(-0.32)在主成分2 中的特征向量绝对值较大,主要反映叶裂长度和果实长度性状;左侧角2(0.34)、着生痕长(0.33)、果厚(0.27)在主成分3 中的特征向量绝对值较大;基部中心到最下方高度(0.41)、翅长(0.40)、叶柄基部宽(-0.34)、叶片基部角(-0.33)在主成分4 中的特征向量绝对值较大,主要反映叶片基部形态;翅宽(0.50)和左下裂片宽(-0.35)在主成分5 中的特征向量绝对值较大;连接角(0.59)、张开角(0.44)、左下裂片长(0.27)在主成分6 中的特征向量绝对值较大。
分别以第一、二主成分向量作为横、纵坐标作散点图,如图6 所示,47 个样本可按地区大致分为3 类,其中,辽东地区的第一主成分向量最大,第二主成分向量最小,辽西地区反之,说明辽东地区各样本整体呈现出叶裂较宽、果实较短的形态特征,辽西地区各样本整体呈现出叶裂窄长、果实较长的形态特征。从散点图分类结果可看出,辽东、辽西地区样本可明显区分,辽中地区样本位于辽东、辽西地区样本之间且距离辽西地区样本更近,说明辽中地区各样本特征与辽西地区的更接近。
2.3.3 表型性状综合评价及指标筛选
将23 个表型性状的标准化值代入上述6 个主成分,得到各样本6 个主成分得分,其中,第1 主成分得分方程为:F1=0.34x1+0.33x2+0.24x3+0.14x4+0.11x5+0.31x6+0.23x7+0.27x8+0.33x9+0.37x10-0.09x11+0.10x12+0.09x13-0.01x14-0.20x15+0.05x16-0.06x17-0.12x18-0.13x19-0.15x20-0.05x21+0.16x22+0.27x23。
利用隸属函数对6 个主成分得分进行归一化处理,计算出6 个主成分的权重系数(0.35、0.24、0.14、0.11、0.09、0.07),得到各样本的综合得分值(F 综合)并排序(表6)。其中SH9 的F 综合值最高(2.358),其表型性状综合表现最好,JC3 的F 综合值最低(-1.873),其表型性状综合表现最差。
利用得到的综合得分值(F综合)与23 个表型性状构建回归方程,进一步筛选性状。通过逐步回归分析构建的回归方程为:y=-9.634+0.040x1+0.024x2+0.052x7+0.055x10+0.107x16+0.083x18+0.110x19-0.004x20-0.416x22。其中x1、x2、x7、x10、x16、x18、x19、x20、x22 分别代表叶片长、叶片宽、左下裂片长、叶基部左侧宽度、翅长、着生痕长、果厚、张开角、翅果比。方程的相关系数r 和决定系数R2 分别为0.988 和0.977,说明这9 个性状可代表F 值总变异的97.7%,F 值为281.134,方程结果极显著,上述性状可作为区分和评价两种槭树的主要性状。
2.4 表型性状聚类分析
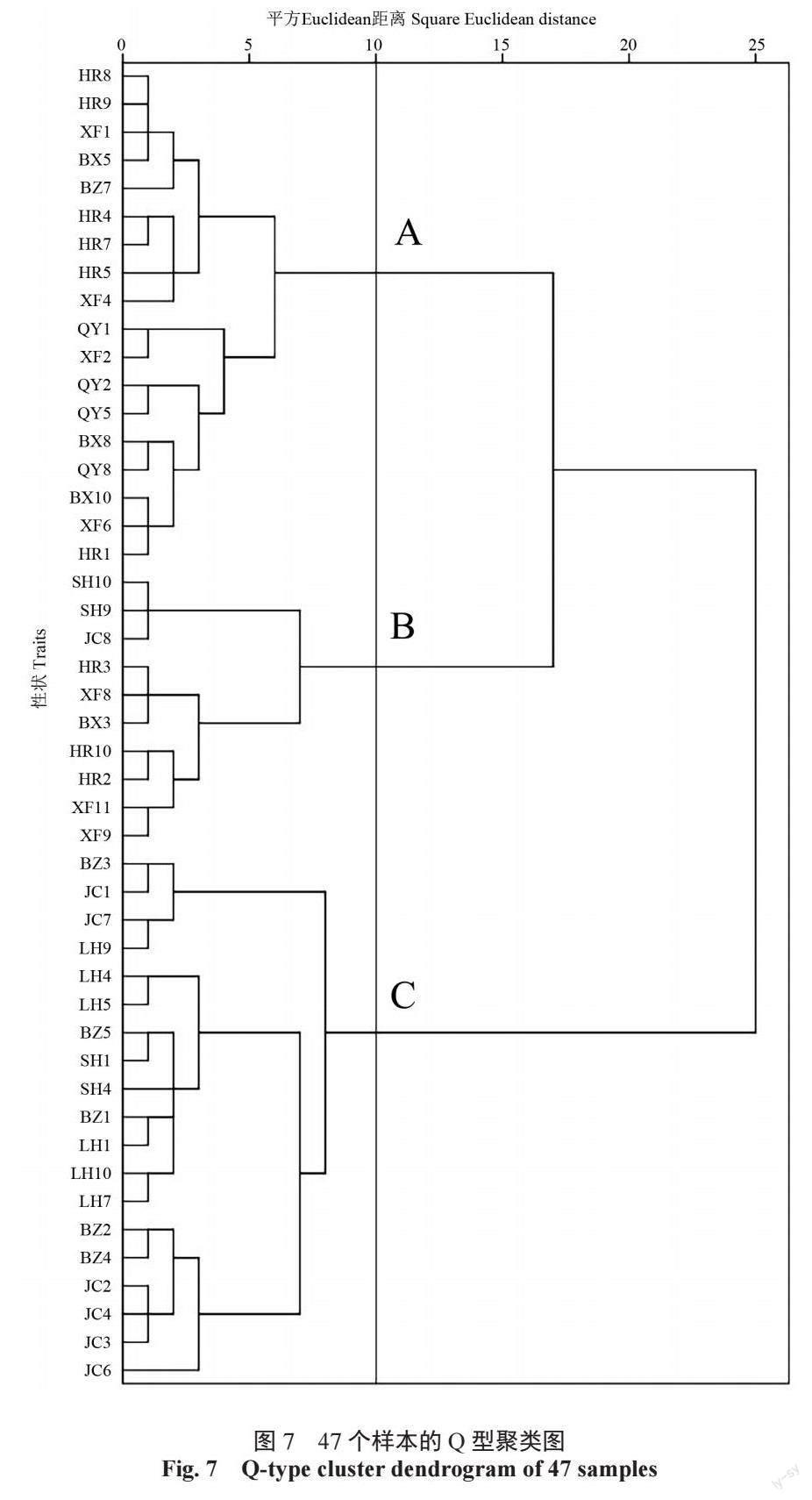
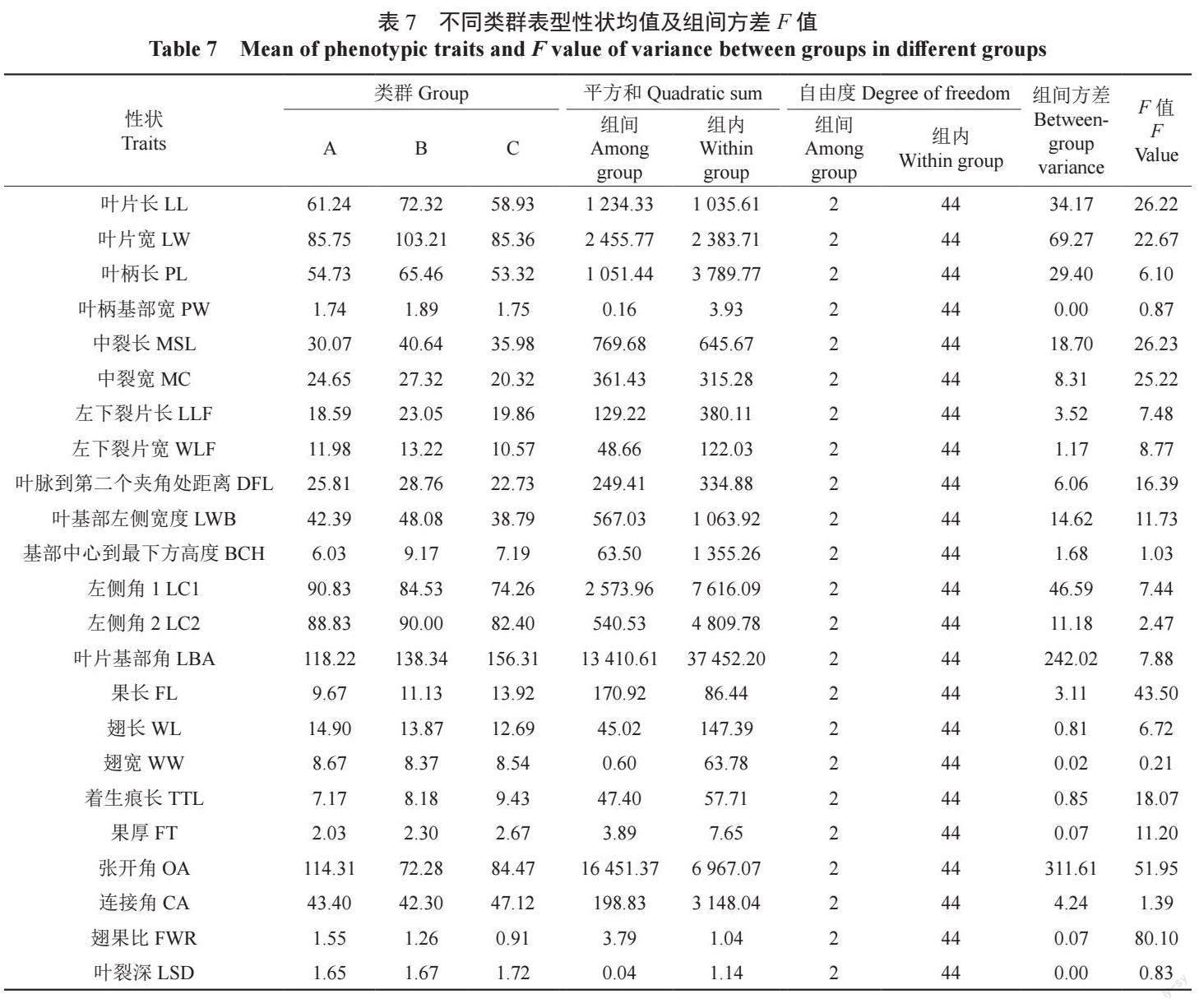
由图7 可见,在遗传距离为10 处,可将47个样本分为A、B、C 三类,各类群表型性状均值及组间方差F 值见表7,其中,果长、张开角、翅果比的组间F 值差异最大。A 类包括18 个样本,其中17 个样本来自辽东地区,仅有1 个样本来自辽西地区,且标准色木槭样本HR4 包括其中,总体特征表现为张开角、翅果比最大,果长最小。
B 类包括10 个样本,分别来自辽东地区的桓仁县、本溪县、西丰县种源,辽中地区的沈河区种源以及辽西地区的建昌县种源,分布范围较广,一些表型特征如翅长、着生痕长、果厚、翅果比介于A、C 类之间,具有两个种的特征。
C 类包括19 个样本,其中17 个样本来自辽西地区,2 个样本来自辽中地区,且标准元宝槭样本JC7 包括其中,总体特征表现为果长最大,翅果比最小。A 类样本表型特征与色木槭更相近,C类样本表型特征与元宝槭更相近。
3 结论与讨论
本研究表明,色木槭与元宝槭的叶片和果实性状具有较高的变异水平且表现出丰富的多样性。其表型性状受经度和海拔的影响显著,辽东地区以色木槭分布为主,辽西地区以元宝槭分布为主。
结合逐步回归分析和Q 型聚类分析结果,叶片长、叶片宽、左下裂片长、叶基部左侧宽度、翅长、果长、着生痕长、果厚、张开角和翅果比共10 个性状可作为区分两种槭树的分类依据,其中果长、张开角、翅果比是色木槭与元宝槭表型的核心分类性状,具有果长≤ 10 mm、张开角≥ 114°、翅果比≥ 1.6的形态特征可认定为色木槭,果长≥ 14 mm、张开角≤ 84°、翅果比≤ 0.9 的形态特征可认定为元宝槭。
植物表型多样性在研究植物差异和对环境的适应性方面具有重要意义[22]。通常变异系数高于10% 说明个体间差异明显[23]。本研究通过对辽宁地区色木槭与元宝槭25 个表型性状进行变异分析发现,两种槭树的叶片和果实性状在种群间和种群内均具有不同程度的丰富变异,且叶片变异程度(16.29%)大于果实(12.95%),这是由于在植物的生长过程中,叶片等营养器官相较于果实等繁殖器官更容易受外界环境影响,即变异幅度更大[24-25]。本文中两种槭树表型性状平均Shannon-Wiener 多样性指数为1.98,与青杨[26]、核桃[27] 相比表现出丰富的多样性,但低于同类槭科植物五角枫[21]、葛萝槭[28],推测这与样本数量、表型性状数量以及种源分布范围等有关。
研究表明,多种木本植物叶片和果实的表型性状受地理气候因子的影响显著,不同植物对所处环境的适应程度和敏感程度有所差异,导致其呈现不同的地理变异模式[29],如茶条槭[25] 叶片及果实性状、苦楝[30] 种子性状与经纬度、海拔及土壤元素等因子具有极强的相关性。本研究通过相关分析和线性回归分析发现元宝槭与色木槭表型性状受经度和海拔的影响显著,与纬度的相关关系不显著,推测是这两种槭树自然分布纬度相近的原因。在表型特征鉴别上,辽东地区各种源样本多为色木槭,而辽西地区各种源样本多为元宝槭,这与元宝槭[31] 的变异规律基本一致,而不同于苦楝[30],说明不同植物对不同环境的适应性不同,从而产生了不同的变异规律。
R 型聚类是对性状进行分类,可以反映出各性状之间的相关性及其选取的合理性[32]。James[33]首先在数量分类中使用R 型聚类,现已广泛应用于橄榄[16]、南京椴[34] 等木本植物中。由于色木槭叶裂深达叶片中段[8],本研究选取叶基部左侧宽度、叶脉到第二个夹角处距离2 个性状,相减可得叶裂深,并参照张世杰[2]、张翠琴[4]、林丽丽[25]选取的性状,选择了25 个表型性状用于区分两种槭树。依据R 型聚类及前人的研究结果,多位学者[6,35-36] 测量了中裂长与中裂宽,而没有选择左上裂片长和左上裂片宽,说明前者更具有代表性,因此可以考虑去除左上裂片长、左上裂片宽2 个性状,其他性状彼此之间较为分散,筛选出的23個性状较为理想,可作为两种槭树的分类指标。
采用主成分分析可以从众多性状指标中筛选核心指标,最大程度地代表原始数据[35]。本研究对23 个表型性状提取6 个主成分,累计贡献率达81.65%,能够反映表型性状的大部分信息。主成分向量散点图将47 个样本依据各种源的地理位置而区分为辽东、辽西地区两大类,辽中地区样本位于辽东、辽西地区样本之间且距离辽西地区种源样本更近,说明辽中地区各样本表型特征与辽西地区更接近。利用主成分分析与逐步回归分析相结合的方法,筛选出叶片长、叶片宽、左下裂片长、叶基部左侧宽度、翅长、着生痕长、果厚、张开角、翅果比9 个性状作为区分和评价两种槭树的主要性状。
Q型聚类可以体现个体间亲缘关系的远近[22],在遗传距离为10 处,可将47 个样本分为3 类。A类样本多来自辽东地区种源,且标准色木槭样本HR4 包括其中,说明该类群表型特征与色木槭更相近;B 类样本具有两个种的特征;C 类样本多来自辽西地区种源,且标准元宝槭样本JC7 包括其中,说明该类群表型特征与元宝槭更相近,这与相关分析和主成分分析的种源分类结果一致。果长、张开角、翅果比在3 个类群间差异最大,可作为辽宁地区色木槭与元宝槭的分类依据,其中具有果长≤ 10 mm、张开角≥ 114°、翅果比≥ 1.6的形态特征可认定为色木槭,果长≥ 14 mm、张开角≤ 84°、翅果比≤ 0.9 的形态特征可认定为元宝槭。
植物的表型性状易受外在环境影响,本研究仅通过表型性状对色木槭与元宝槭进行多样性分析及分类学研究存在一定的局限性,后续考虑将其与花粉形态研究和基因水平研究相结合,综合分析两种槭树的遗传多样性,为其资源鉴定和种间分类提供更全面的科学依据。
参考文献:
[1] 徐廷志. 翅果形态及其在槭树科分类与演化上的意义[J]. 广西植物,1996(2):109-122.
XU T Z. Samara shape of Aceraceae and its implications insystematics and evolution[J]. Guangxi plants,1996(2):109-122.
[2] 张世杰. 五种槭树属植物叶子及种实性状研究[D]. 咸阳: 西北农林科技大学,2017.
ZHANG S J. Five species ofAcer leaves and seed characterresearch[D]. Xianyang: Northwest Agriculture & Forestry University,2017.
[3] 谢影, 于海洋, 庞忠义, 等. 东北地区6 种槭属植物种子表型多样性分析及优良家系选择[J]. 植物科学学报,2021,39(6):610-619.
XIE Y, YU H Y, PANG Z Y, et al. Phenotypic diversity analysisand superior family selection of seed traits in sixAcer species innortheast China[J]. Plant Science Journal,2021,39(6):610-619.
[4] 张翠琴. 五角枫种群遗传多样性研究[D]. 太原: 山西师范大学,2015.
ZHANG C Q. Genetic diversity of population inAcer monoMaxim.[D]. Taiyuan: Shanxi Normal University,2015.
[5] 乔谦, 丰震, 任红剑, 等. 元宝枫天然种群种实表型多样性[J].生态学杂志,2017,36(10):2707-2715.
QIAO Q, FENG Z, REN H J, et al. Phenotypic diversity of wingfruits and seeds in natural populations ofAcer truncatum Bungein China[J]. Chinese Journal of Ecology,2017,36(10):2707-2715.
[6] 任红剑, 丰震, 乔谦, 等. 元宝枫叶片形态特征的地理变异[J].西北林学院学报,2017,32(6):113-119.
REN H J, FENG Z, QIAO Q, et al. Geographic variation trendof leaf morphology in Acer truncatum[J]. Journal of NorthwestForestry University,2017,32(6):113-119.
[7] 王性炎, 王姝清. 新资源食品—元宝枫籽油[J]. 中国油脂,2011,36(9):56-59.
WANG X Y, WANG S Q. New resource food-Acer truncatumBunge seed oil[J]. China Oils and Fats,2011,36(9):56-59.
[8] 方文培. 槭树科·中国植物志[M]. 北京: 科学出版社,1981:66-273.
FANG W P. Aceraceae·Flora of China[M]. Beijing: Science Press,1981:66-273.[9] 中国树木志编辑委员会. 中国树木志[M]. 北京: 中国林业出版社,1997:2005.Editorial Committee of Chinese Tree Records. Chinese TreeRecords[M]. Beijing: China Forestry Publishing House,1997:2005.
[10] 陈清霖. 两种五角枫的形态识别与应用[J]. 辽宁林业科技,2018(6):54-55.
CHEN Q L. Morphological identification and application of twokinds ofAcer mono[J]. Liaoning Forestry Science and Technology,2018(6):54-55.
[11] TAO J, QIAO G, WEN X P, et al. Characterization of geneticrelationship of dragon fruit accessions (Hylocereus spp.)by morphological traits and ISSR markers[J]. Scientia Horticulturae,2014,170:82-88.
[12] PENG L, RU M, WANG B Q, et al. Genetic diversity assessmentof a germplasm collection ofSalvia miltiorrhiza Bunge. based onmorphology, ISSR and SRAP markers[J]. Biochemical Systematicsand Ecology,2014,55:84-92.
[13] PIGLIUCCI M, MURREN C J, SCHLICHTING C D.Phenotypic plasticity and evolution by genetic assimilation[J].Journal of Experimental Biology,2006,209(12):2362-2367.
[14] LEIMAR O. Environmental and genetic cues in the evolution ofphenotypic polymorphism[J]. Evolutionary Ecology,2009,23(1):125-135.
[15] 包文泉. 基于形态、叶绿体基因组及核基因组SSR 的仁用杏分类地位研究[D]. 北京: 中国林业科学研究院,2017.
BAO W Q. A taxonomic study on kernel-using apricot usingmorphological traits, chloroplast genome and nuclear genomeSSR[D]. Beijing: Chinese Academy of Forestry Sciences,2017.
[16] 吴如健, 万继锋, 韦晓霞, 等. 橄榄种质资源果实表型性状多样性分析及其数量分类研究[J]. 果树学报,2015,32(5):797-805.
WU R J, WAN J F, WEI X X, et al. Fruit character diversityanalysis and numerical classification of Chinese olive germplasmresources[J]. Journal of Fruit Science,2015,32(5):797-805.
[17] 柏文富, 張文昕, 禹霖, 等. 不同樱属植物叶形态与解剖结构的比较[J]. 中南林业科技大学学报,2022,42(8):15-26.
BAI W F, ZHANG W X, YU L, et al. Comparison of leaf morphologyand anatomical structures of differentCerasus species[J].Journal of Central South University of Forestry & Technology,2022,42(8):15-26.
[18] 兰庆, 刘果厚, 胥晓, 等. 细裂槭与其近缘种的比较形态学及分类学研究[J]. 西北植物学报,2019,39(1):92-101.
LAN Q, LIU G H, XU X, et al. Comparative morphology ofAcer stenolobum and its relevant species and their taxonomysignificance[J]. Acta Botanica Boreali-Occidentalia Sinica,2019,39(1):92-101.
[19] 薛昱婷, 劉军, 刘晓莉, 等.18 个木芙蓉品种与2 个近缘种的数量分类和主成分分析[J]. 中南林业科技大学学报,2022,42(1):70-77.
XUE Y T, LIU J, LIU X L, et al. Quantitative classification andprincipal component analysis of 18Hibiscus mutabilis cultivarsand 2 related species[J]. Journal of Central South University ofForestry & Technology,2022,42(1):70-77.
[20] 王东来, 周文杰, 姚平, 等. 软枣猕猴桃种质资源表型性状的数量分类研究[J]. 北方园艺,2022(10):33-40.
WANG D L, ZHOU W J, YAO P, et al. Numerical classificationofActinidia arguta planch germplasm resources accordingphenotypic characters[J]. Northern Horticulture,2022(10):33-40.
[21] 姬志峰. 山西五角枫天然种群表型多样性研究[D]. 太原: 山西师范大学,2013.
JI Z F. Study on phenotypic diversity of natural populationinAcer mono Maxim in Shanxi[D]. Taiyuan: Shanxi NormalUniversity, 2013.
[22] 刘莉, 王磊, 吴丹, 等. 不同种源文冠果种子的表型变异[J].经济林研究,2021,39(4):97-105.
LIU L, WANG L, WU D, et al. Phenotypic variation analysis ofseeds from different provenances ofXanthoceras sorbifolium[J].Non-wood Forest Research,2021,39(4):97-105.
[23] 董博文, 李继东, 郑先波, 等. 山茱萸种质资源表型性状多样性及相关性分析[J]. 经济林研究,2014,32(2):163-166.
DONG B W, LI J D, ZHENG X B, et al. Diversity and correlationanalysis of phenotypic characteristics inCornus officinalisgermplasm resources[J]. Non-wood Forest Research,2014,32(2):163-166.
[24] 井振华, 李皓, 邵文豪, 等. 浙江柿天然群体表型多样性研究[J]. 植物研究,2010,30(3):325-331.
JING Z H, LI H, SHAO W H, et al. Phenotypic diversity of naturalpopulations inDiospyros glaucifolia[J]. Bulletin of BotanicalResearch,2010,30(3):325-331.
[25] 林丽丽. 茶条槭种群表型差异研究及潜在分布区预测[D]. 太原: 山西师范大学,2015.
LIN L L. Phenotypic variations and potential distribution areaprediction ofAcer Ginnala populations[D]. Taiyuan: ShanxiNormal University,2015.
[26] 曹德美, 张亚红, 成星奇, 等. 青杨不同种群叶片表型性状的遗传变异[J]. 林业科学,2021,57(8):56-67.
CAO D M, ZHANG Y H, CHENG X Q, et al. Genetic variationof leaf phenotypic traits in different populations ofPopuluscathayana[J]. Scientia Silvae Sinicae,2021,57(8):56-67.
[27] 李根, 邓凤彬, 金强, 等. 基于坚果表型性状的新疆老龄核桃种质资源多样性分析[J]. 中国果树,2021(7):33-38.
LI G, DENG F B, JIN Q, et al. Diversity and comparison ofgermplasm resources of aged walnut in Xinjiang based onphenotypic traits of nuts[J]. China Fruits,2021(7):33-38.
[28] ZHANG Q D, JIA R Z, MENG C, et al. Diversity and populationstructure of a dominant deciduous tree based on morphologicaland genetic data[J]. AoB Plants,2015,7:plv103.
[29] ZENG Y F, ZHANG J G, ABUDUHAMITI B, et al.Phylogeographic patterns of the desert poplar in NorthwestChina shaped by both geology and climatic oscillations[J]. BMCEvolutionary Biology,2018,18(1):1-14.
[30] 陈丽君, 邓小梅, 丁美美, 等. 苦楝种源果核及种子性状地理变异的研究[J]. 北京林业大学学报,2014,36(1):15-20.
CHEN L J, DENG X M, DING M M, et al. Geographic variationin traits of fruit stones and seeds ofMelia azedarach[J]. Journalof Beijing Forestry University,2014,36(1):15-20.
[31] QIAO Q, YE M J, WU C, et al. Analysis of leaf morphologyvariation and genetic diversity via SRAP markers for nearthreatenedplantAcer truncatum[J]. Global Ecology andConservation,2022,33:e01980.
[32] SNEATH P H A. The principles and practice of numericalclassification[M]. Numerical taxonomy,1973,573.
[33] JAMES M T. Numerical Taxonomy[J]. Evolution,1964,18:513-513.
[34] 严灵君, 黄犀, 岳远灏, 等. 南京椴天然居群的表型性状多样性及变异分析[J]. 植物资源与环境学报,2021,30(3):29-37.
YAN L J, HUANG X, YUE Y H, et al. Analyses on diversityand variation of phenotypic traits of natural populations ofTiliamiqueliana[J]. Journal of Plant Resources and Environment,2021,30(3):29-37.
[35] 王宁, 牛立新, 张延龙, 等. 元宝枫天然居群表型多样性研究[J]. 北方园艺,2019(8):90-96.
WANG N, NIU L X, ZHANG Y L, et al. Phenotypic diversity ofAcertruncatum natural populations in China[J]. Northern Horticulture,2019(8):90-96.
[36] 齐新玲, 丰震, 马立敏. 元宝枫无性系幼龄期枝叶性状变异分析[J]. 山东农业科学,2015,47(11):27-31.
QI X L, FENG Z, MA L M. Variation analysis on branch andleaf traits ofAcer truncatum bunge clones at young stage[J].Shandong Agricultural Sciences,2015,47(11):27-31.
[ 本文編校:赵 坤]
猜你喜欢
西北民族大学学报(自然科学版)(2021年4期)2021-12-29
数学小灵通(1-2年级)(2021年4期)2021-06-09
大众健康(2021年6期)2021-06-08
小聪仔(科普版)(2020年12期)2021-01-18
东方少年·布老虎画刊(2020年4期)2020-06-08
学生天地(2019年32期)2019-08-25
中学生数理化·七年级数学人教版(2019年4期)2019-05-20
中学生数理化·七年级数学人教版(2018年6期)2018-06-26
小天使·一年级语数英综合(2017年11期)2017-12-05
初中生世界·七年级(2017年9期)2017-10-13
