
高州油茶杂交F1代群体构建及遗传评估Ⅱ
2024-01-30 14:28张坤昌廖柏勇黄润生罗慧华涂攀峰王溢代文魁吕宇宙李永泉
经济林研究 2023年3期
关键词:杂交
张坤昌 廖柏勇 黄润生 罗慧华 涂攀峰 王溢 代文魁 吕宇宙 李永泉


摘 要:【目的】选择优良杂交组合,提高油茶产量与品质。【方法】选取16 个油茶品系作为研究对象,包括6 個高州油茶品系、7 个普通油茶品系、1 个小果油茶品系、1 个香花糙果茶品系和1 个南山茶品系,采用单交交配设计,设计27 个杂交组合,估算杂交亲和性的一般配合力和特殊配合力,分析杂交果实大小、质量、果皮厚等性状以及杂交苗生长性状遗传参数,初步筛选高州油茶优良杂交亲本及优良杂交组合。【结果】参试亲本中:高州油茶为六倍体、八倍体和十倍体,其中以十倍体居多;普通油茶为四倍体、六倍体和十倍体,其中以六倍体居多;小果油茶是六倍体;南山茶和香花糙果茶为四倍体。27 个杂交组合总套袋数量为1 360,保存数量1 048,坐果数量519,收获杂交果实285 个,获得籽粒数量1 894,坐果率最高的组合为CS1×CG2(86.27%),成果率最高的组合为CS1×CG2(76.47%)。坐果数量在母本间一般配合力效应值最高的是CG2,为13.74;父本间的一般配合力效应值最高的是CG1,为11.74。坐果数量的特殊配合力效应值以CB1×CG1 最高(31.03),其次是CG2×CS1(22.26)和CS1×CG2(20.43)。杂交组合CS1×CG2 在单果质量、横纵径、果形指数和果皮厚度性状方面表现优良,在苗期生长性状方面,发芽率高、长势好,在子代中生长最旺盛。【结论】参试高州油茶品系中,CG2 和CG1 杂交亲和性表现较好,采用杂交组合CS1×CG2 有望培育出优良的油茶杂交品种。
关键词:高州油茶;杂交;F1 代群体;子代苗;遗传评估
中图分类号:S603.2 文献标志码:A 文章编号:1003—8981(2023)03—0091—15
高州油茶Camellia gauchowensis,异名陆川油茶C. fangchengensis,在《中国植物志》中被修订为越南油茶C. drupifera[1],是我国南缘油茶的重要组成部分。主要分布于海南岛、广东西南部高州至茂名、广西柳州和陆川以及广西与广东交界处[2]。高州油茶树体高大,单位面积产量高,且果实出籽率、种子出仁率较高[3]。其油脂组分主要有油酸、棕榈酸、亚油酸、硬脂酸和亚麻酸,其中不饱和脂肪酸总量占84%[4-5]。我国中果型油茶品种稀缺,从高州油茶资源中选育中大果型油茶品种,成为培育适合机械化采摘品种的重点。选育高州油茶大果型油茶新品种的优良杂交亲本为高产品系培育的关键环节[2]。
杂交育种在油茶育种中应用广泛,部分杂交子代杂种优势明显。常维霞等[6] 对4 个油茶物种进行自交和异交人工控制授粉,结果表明按照结实率和结籽数量由高到低排列,依次为异株异花、自由授粉、同株异花自交、自花自交;李晓春等[7]通过人工杂交控制授粉,筛选出6 个较佳配置组合;江泽鹏等[8] 对2 个地理种源普通油茶进行杂交授粉,研究杂交果实性状以及F1 代苗期生长特性,发现‘岑软系列和‘湘林系列中‘岑软24 号ב湘林210和‘湘林210ב岑软2 号2 个组合在坐果率、成果率、单果质量和鲜出籽率方面表现良好,苗期生长速度均较快;梁恬等[9] 通过对普通油茶优良无性系杂交果实性状进行变异分析,确定了杂交优良母本、优良父本以及优良杂交组合;林萍等[10-11] 通过估算普通油茶‘长林系列种内杂交子代遗传参数,分析其配合力,筛选出优良杂交组合。
油茶为自交不孕树种,自交结实率为0 ~2.9%[12-13],合子前自交不亲和、合子后细胞发育不良、染色体倍性差异和授粉昆虫特异性等问题常导致其授粉失败[14-15],人工杂交试验是分析授粉因素和保障授粉成功的主要方法[16]。除单位面积产量,选育高含油类、高出籽类、大籽类、皮薄类和大果类等五大类果实性状为油茶杂交育种亲本选择的主要目标[17]。杂交试验结果表明,高州油茶种内杂交情况较好[18]。为了检测高州油茶杂交的可靠性,获得更多高产稳产品系和更多杂交种子,本研究中拟扩大高州油茶杂交试验范围,选取高州油茶6 个品系以及小果油茶C.meiocarpa 1 个品系、普通油茶C. oleifera 7 个品系、香花糙果茶C. suaveolens 1 个品系以及南山茶C.semiserrata 1 个品系作为研究对象,根据前期初步杂交试验结果,设计27 个单交杂交组合,估算各亲本杂交亲和性的一般配合力和特殊配合力,分析杂交果实性状和杂交苗生长性状,筛选高州油茶优良杂交亲本和杂交组合。
1 材料与方法
1.1 试验地概况
试验地位于广东省韶关市曲江区国有小坑林场(113°35′E,24°15′N)。该林场地处南岭山脉大庾岭南缘,属亚热带气候区,平均最高气温34.0 ℃,平均最低气温10.0 ℃,年均气温20 ℃,年降水量265.8 mm,砖红壤土质。
1.2 试验材料
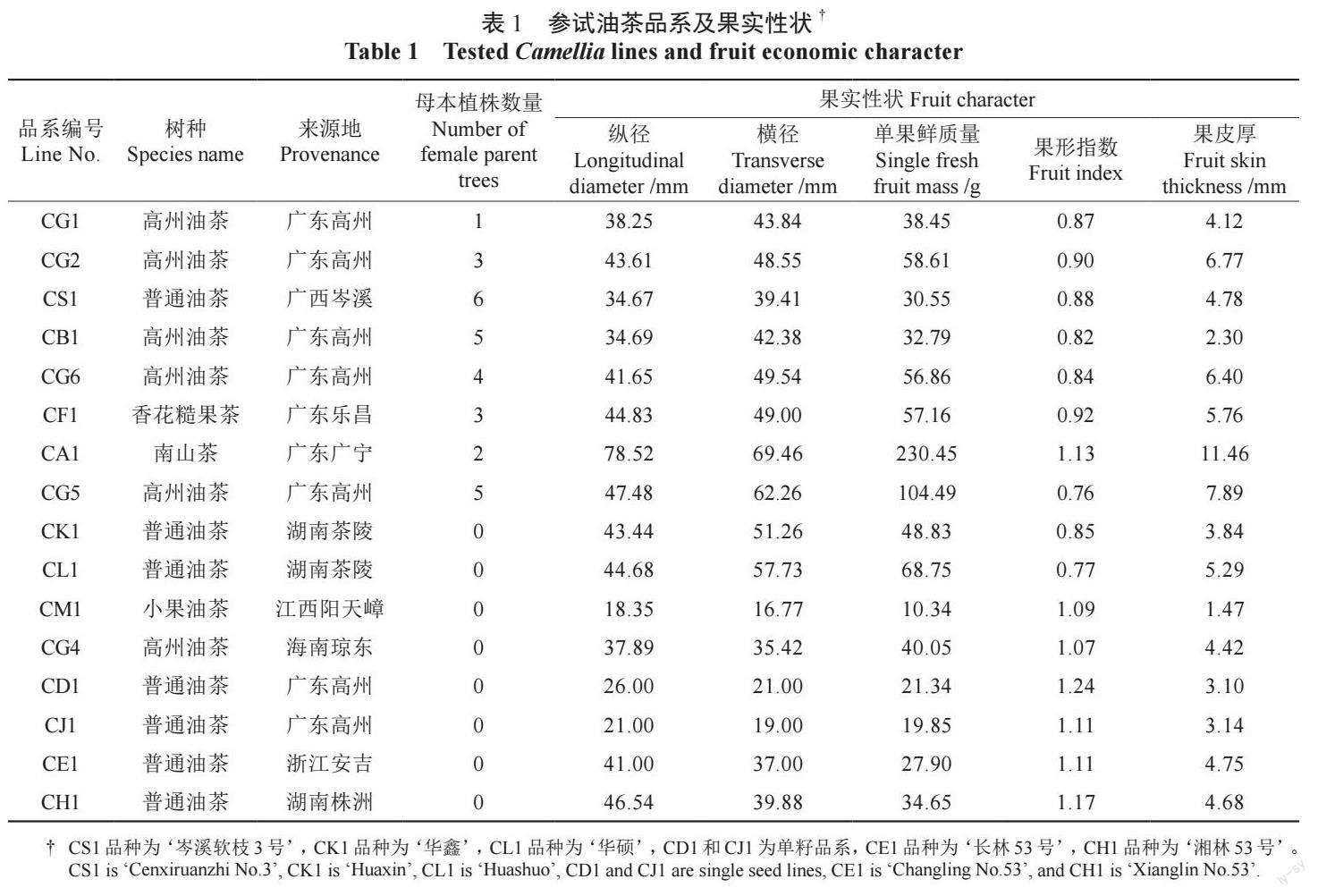
参试亲本共16 个品系,包括6 个高州油茶品系(5 个来自广东高州、1 个来自海南琼东),7 个普通油茶品系(分别来自广西岑溪、湖南茶陵、湖南株洲和浙江安吉),1 个小果油茶品系(江西阳天嶂),1 个香花糙果茶品系(广东乐昌),1 个南山茶品系(广东广宁),具体见表1。授粉母树于2015 年3 月5 日定植。
油茶种内和种间的果实经济性状各有特征,观测母本的果实纵径、横径、单果鲜质量、果形指数和果皮厚性状。发现高州油茶CG5 的横纵径和单果鲜质量较大,但其果皮较厚;香花糙果茶CF1 果实大小、单果鲜质量也较大,果皮厚度中等;普通油茶CS1 的横纵径和单果鲜质量较小,果皮较薄;高州油茶CB1 果皮较薄。南山茶的果实单果鲜质量远高于其他物种。
1.3 杂交试验设计
授粉试验时间为2020 年11 月23—27 日,共5 d。阴天,雾气重,偶尔小雨,温度为8 ~ 16 ℃。參试品系共16 个,设计27 个单交杂交组合,每株母树仅接受1 个父本的花粉,具体见表2。用10 cm×10 cm 的硫酸纸袋进行授粉套袋,统计授粉数量,并挂编号吊牌。
1.4 指标测定和数据分析
参照CyStain? UV Precise P Automate 试剂盒的方法,使用Sysmex CyFlow? 染色体倍性分析仪(德国)分析各亲本染色体倍性[18]。
授粉19 d 后,于2020 年12 月16—17 日,统计保存数量和保存率以及每个杂交组合10 个子房横径;授粉106 d 后,于2021 年3 月13—14 日,统计坐果数量和坐果率以及每个杂交组合10 个杂交果实的果实横径;于2021 年10 月11—12 日采收全部杂交果实,统计其成果率,测量横径、纵径、果皮厚,称量单果鲜质量,统计籽粒数量。2021 年11 月20 日将收集的杂交种子播种,2022 年4 月23 日观测杂交种子发芽率,2022 年5 月18 日测量杂交苗高,同时观测亲本苗高作为对照。
保存率为授粉19 d 后子房发育正常的果实数量占授粉花朵总数量的百分比,坐果率为授粉106 d后子房发育正常的果实数量占授粉花朵数量的百分比,成果率为收获成熟果实数量占授粉花朵数量的百分比,发芽率为正常发芽种子数量占供试种子数量的百分比。
采用Origin 2019、Excel 2010 软件完成图表绘制及数据整理;采用SPSS 22.0 软件统计保存率、坐果率、成果率、子房横径、果实横纵径、单果质量、果皮厚以及苗高。对不同组合间性状差异采用单因素方差分析模型Ymn=η+αm+βn+εmn 估算,其中:Ymn 为第m 次杂交第n 个母本的表型值,η 为总体平均值,αm 表示处理效应,βn 为区组效应,εmn 代表随机误差。进一步对差异显著性状进行多重比较(邓肯氏法),估算遗传力[8] 和遗传增益[19]。
h2=1-1/F;ΔG=R/xp;R=Sh2;S=xo-xp。
式中:h2 为遗传力;F 为组间项均方与组内项均方的比值;ΔG 为遗传增益;R 为选择响应;S 为选择差;xo 为中选平均值;xp 为总体平均值。
采用R 软件运行ASreml-R 4.0 软件包,估算一般配合力和特殊配合力[20-21]。
yijk=μ+Ri+Gj+Gk'+Sjk+eijk。
式中:yijk 是第i 次杂交第j 母本和第k 父本的亲和性观测值;μ 是整体亲和性总体均值的固定效应;Ri 是第j 母本和第k 父本的第i 次重复杂交的固定效应;Gj 是第j 母本的一般配合力(GCA)随机效应值;Gk' 是第k 父本的一般配合力随机效应值;Sjk 是第j 母本和第k 父本的特殊配合力(SCA)的随机效应值;eijk 是随机误差。
2 结果与分析
2.1 参试品种染色体倍性估测
染色体可以为植物进化和系统发育研究提供有价值的信息,可以为品种间亲缘关系、倍性鉴定、种质资源鉴定提供细胞学依据[22-23]。参试油茶样品流式细胞峰值分布如图1 所示。
参试油茶样品染色体倍性估测参数见表3,在3 个对照品种互为对照的情况下,CK1‘华鑫的染色体倍性平均预估值为6.38,CL1‘华硕为6.02,CS1‘岑软3 号为7.51。参试样品中,CM1、CB1、CG5、CD1 和CJ1 为六倍体,CF1、CA1 和CE1 为四倍体,CG1、CG2、CG4 和CH1 为十倍体,CG6 为八倍体。
2.2 参试品种杂交授粉结实状况
各组合杂交授粉后结实情况如表4 所示。共计27 个杂交组合,总套袋数量为1 360,授粉19 d后授粉花朵保存数量为1 048,授粉106 d 后授粉坐果数量为519,收获杂交果实285 个,获得籽粒数量为1 894。保存率较高的组合均为高州油茶与普通油茶的杂交组合,即CG2×CK1、CG2×CL1、CS1×CD1 和CG5×CH1, 均为96.00%;保存率最低的组合为香花糙果茶与小果油茶的杂交组合CF1×CM1,为2.00%。坐果率最高的组合为普通油茶与高州油茶杂交组合CS1×CG2,为86.27%; 坐果率较低的组合有普通油茶与红花油茶、香花糙果茶与小果油茶、香花糙果茶与红花油茶以及红花油茶与小果油茶杂交组合, 即CS1×CA1、CF1×CM1、CF1×CA1 和CA1×CM1,均为0。在成果率方面,成果率较高的组合有CS1×CG2、CS1×CG1、CB1×CM1、CB1×CG2 和CB1×CJ1,分别为76.47%、52.00%、52.00%、46.67% 和43.59%,这5 个杂交组合的坐果率和成果率均排在前5 位。
从母本和父本的杂交结果来看(表5),母本间保存率差异度为abc, 均值为71.64%, 其中保存率最高的CG2 为96.00%,最低的CF1 为31.33%。而坐果率差异度为ab,均值为37.33%,其中坐果率最高的CB1 为63.32%,其次是CG6(47.50%) 和CG2(47.00%), 坐果率最低的CA1 为0。成果率之间不存在显著差异,均值为17.20%,其中成果率最高的CB1 为35.44%,其次是CS1(29.75%),最低的CA1 为0。母本间发芽率差异度为ab,均值为10.02%,其中发芽率最高的CG5 为43.66%,最低为CG1、CA1 和CF1,均为0。
父本间的保存率差异度为ab,均值为81.62%,其中保存率最高的CL1 和CD1 均为96.00%,保存率最低的CA1 为61.22%。坐果率间无显著差异,均值为39.25%,其中坐果率最高的CG2 为79.80%,其次是CJ1(74.36%)和CG1(60.00%),坐果率最低的CD1 为12.00%。成果率间无显著差异,均值为23.43%,其中成果率最高的CG2为61.57%,最低的CD1 为6.00%。父本间发芽率无显著差异,均值为13.89%,其中发芽率最高的CH1 为35.88%,最低为CS1 和CA1,均为0。
南山茶CA1 作为母本和父本的坐果率均较低,分别为0 和12.96%。当其作为母本时,授粉19 d后保存率为56.25%,到授粉106 d 后全部死亡,说明其他树种的花粉在南山茶柱头上能识别生长,但在后续的受精、胚胎发育过程中遇到障碍。南山茶CA1 作为父本时,保存率61.22%,坐果率12.96%,说明南山茶与其他树种的杂交亲和力较小。此外,CG2 作为父本时坐果率最高,达到了79.80%,说明CG2 的花粉亲和性较好,能在不同树种柱头上正常生长、受精和发育。CS1、CA1 和CD1 作为父本时,坐果率均低于25.00%,不建议作为优良品系的授粉树。
2.3 参试品种杂交亲和性的配合力
根据估算的子代某性状配合力,可以预测亲本该性状的子代遗传整体趋势。可采用混合线性模型[24] 计算保存数量和坐果数量这一类阈性状的遗传参数。油茶杂交后保存数量和坐果数量的混合线性模型方差分析结果如表6 所示,其中母本间、父本间和组合间效应值均达到极显著水平,说明母本间、父本间保存数量和坐果数量的一般配合力和组合间的特殊配合力均存在极显著的选择空间。
采用上述保存数量和坐果数量的混合线性模型,估算保存数量和坐果数量的一般配合力效应值和特殊配合力效应值。从表7 可以看出:保存数量在母本间的一般配合力效应值最高的是CG1,达到了21.18,其次是CG2、CS1,分别为17.18和12.85,最低是CA1 的-26.32;父本间的一般配合效应值最高是CL1 和CD1,均为17.18,其次是CK1、CH1,分别为13.78 和13.68,最低是CJ1 的-18.82。从表8 可以看出:保存数量的特殊配合力效应值最高的组合为CB1×CG1(33.55),其次是CS1×CG2(15.90)和CB1×CA1(15.22),特殊配合力效应值最低的组合是CG6×CK1,为-19.28。
从表7 可以看出:坐果数在母本间一般配合力效应值最高的是CG2,为13.74,其次是CG1的11.74 以及CG6 的8.49,最低是CA1 的-15.26;父本间的一般配合力效应值最高的是CG1,为11.74, 接着是CK1(7.94) 和CG4(7.74), 最低是CA1(-11.76)。从表8 可以看出:坐果数量的特殊配合力效应值以CB1×CG1 最高(31.03),其次是CS1×CG2(20.43),坐果数量最低的是CG5×CK1,为-16.14,表明此组合不但人工授粉花朵的保存较难,而且在坐果数量上处于劣势。
2.4 参试品种F1 代子房和果实横径的发育状况
在记录杂交試验保存数量的过程中,随机测量每个杂交组合10 个子房横径、授粉106 d后果实横径和采收时果实横径,结果如表9 所示。在子房横径与授粉106 d 后果实横径之间,子房横径最大的组合为CG1×CK1,授粉106 d后果实横径最大的组合为CB1×CG2,增长幅度较大的组合有CB1×CG1、CB1×CG2、CG5×CB1、CG5×CH1、CG5×CK1、CG6×CB1、CG6×CH1、CG6×CK1、CS1×CG1 和CS1×CG2。另外在测量观察中发现:杂交组合CS1×CD1 和CB1×CA1 授粉果实大小在授粉后87 d 内几乎不变,说明其生长发育较为缓慢;杂交组合CA1×CM1、CF1×CA1 和CS1×CA1 子代均死亡, 可见其生长发育的不适性。
观察在授粉106 d 后与采收时果实横径的动态变化,发现杂交组合CF1×CS1 增长幅度最大,然后依次为CG2×CL1、CG5×CH1、CG6×CB1、CG1×CK1,其他杂交组合均有不同程度的增长,且上述5 个杂交组合采收时果实横径位于前列。
2.5 参试品种F1 代果实性状和子代苗高差异及遗传增益
对各杂交组合果实性状、子代苗高进行单因素方差分析,结果见表10。由表10 可知,单果质量、纵径、横径、果形指数和果皮厚等性状在杂交组合间差异均达到极显著水平(P?0.01),组合内区组间差异不显著。杂交果实的单果质量、纵径、横径、果形指数和果皮厚性状的平均值分别为29.81 g、34.01 mm、38.34 mm、0.89 和4.71 mm,单果质量最大值是最小值的44.24 倍,果皮厚最大值是最小值的7.32 倍,果实大小差异也较大。说明不同杂交组合间亲本的遗传差异较大。不同杂交子代间苗高差异极显著(P < 0.001),这为优良基因型的选择提供了可能。遗传力高表明性状受遗传控制强,选择的可能性也较高。经计算,苗高的遗传力为0.939 4,说明苗高易受遗传控制,能够较稳定遗传。
进一步对差异显著性状进行多重比较,结果见表11。杂交组合CF1×CS1 单果质量最大,达55.84 g,然后是杂交组合CG2×CL1(55.83 g),CG6×CB1(55.27 g), 杂交组合CB1×CJ1 单果质量最小,仅为9.12 g;杂交组合CF1×CS1的横纵径也较大;杂交组合CB1×CG1 果皮最薄,为2.50 mm,杂交组合CG2×CL1 果皮最厚(7.54 mm)。
由表12 可知:果实纵径、横径遗传增益最大的杂交组合是CF1×CS1,比所有杂交组合总体平均值分别高32%、26%, 其次是杂交组合CG2×CL1,分别是总体平均值的1.22、1.23 倍;单果质量遗传增益最大的杂交组合是CF1×CS1,比所有杂交组合总体平均值高87%,其次是杂交组合CG2×CL1,是总体平均值的1.86 倍;果皮厚度遗传增益最小的杂交组合为CB1×CG1,比所有杂交组合总体平均值低46%,其次是杂交组合CB1×CM1,是总体平均值的54%。
综上分析,杂交组合CS1×CG2、CG2×CL1、CG6×CB1、CF1×CS1 和CB1×CG1 在杂交果实性状中表现较为优良,其中CF1×CS1 单果质量最大,CB1×CG1 果皮厚最小,CS1×CG2、CG2×CL1 和CG6×CB1 各性状表现均较优良。
连续2 个月对不同杂交组合苗与亲本苗的高度进行测量,结果见表13。由表13 可知,不同杂交组合间差异较大。除亲本外,杂交组合CS1×CG2的苗高处于最高水平,5 月苗高为11.33 cm,6 月苗高为14.18 cm;其次是杂交组合CS1×CG4,5月苗高为9.67 cm,6 月苗高为13.25 cm。这2 个组合杂交苗的高度在1 个月内变化最为明显,其余组合杂交苗高度的变化不明显,甚至长势渐弱至枯萎死亡。
3 结论与讨论
结合杂交果实性状与苗高进行分析,结果表明CS1×CG2、CF1×CS1 和CB1×CG1 是此次杂交试验中较为优良的杂交组合。CF1×CS1 具有单果质量最大的特性;CB1×CG1 具有果皮最薄的特性;CS1×CG2 不但在保存率、坐果率、保存数量、坐果数量方面具有优势,而且杂交果实性状较为优良,种子发芽率较高,苗期生长速度均较快,是本次杂交试验中最好的组合。远缘杂交和种内杂交可以提高高州油茶产量,改良其综合品质。在参试高州油茶品系中,CG2 与CG1 杂交亲和性表现较好,杂交组合CS1×CG2 有望被培育成优良的油茶杂交品种。
3.1 亲本倍性对杂交授粉成功率和配合力的影响
通过对高州油茶、普通油茶、小果油茶、香花糙果茶和南山茶5 个树种进行杂交试验,筛选具有高亲和性的杂交组合,并对杂交果实部分数量性状和杂交苗生长性状进行遗传分析,选配出高州油茶优良杂交亲本。参试5 个树种种间及种内杂交的各杂交组合间亲和力相差较大。不同树种作为母本时,平均坐果率为37.33%,高州油茶CB1 坐果率最高(63.32%),南山茶CA1 坐果率最低(0);不同树种作为父本时,平均坐果率为39.25%,CG2 坐果率最高(79.80%),普通油茶CD1 坐果率最低(12.00%)。高州油茶作为母本时平均坐果率为41.04%,普通油茶作为母本时为44.38%,而香花糙果茶作为母本时仅6.67%,南山茶作为母本时为0;高州油茶作为父本时平均坐果率为57.20%,普通油茶作为父本时为34.30%,南山茶作为父本时为12.96%,小果油茶作为父本时为28.40%,其中小果油茶作为父本的坐果率的变异幅度为0 ~ 80.00%,说明小果油茶与其他树种杂交成活率差异极大,这与张乃燕等[25] 报道的结果相类似。因此,高州油茶在种间种内杂交中更适合作为父本,普通油茶更适合作为母本,小果油茶、香花糙果茶不适宜作为父本与南山茶进行种间杂交。
双列交配设计在作物和林木改良实践中已被广泛使用[26-30]。本研究中采用非全双列杂交方法对配合力进行了估算。CG1 和CG2 作为父本或者作为母本的保存数量和坐果数量的一般配合力均较高。特殊配合力以CB1×CG1 为最高,其保存数量和坐果数量的特殊配合力分别为33.55、31.03;其次是CS1×CG2,保存数量和坐果数量的特殊配合力分别为15.90、20.43。树种亲本的一般配合力和特殊配合力的相对重要性受测试材料、性状、年龄和地点等因素的影响,通常一般配合力的选择比特殊配合力更重要[31]。因此,CG1 和CG2 在保存数量、坐果数量的一般配合力和特殊配合力方面均存在明显优势,可作为高州油茶品种选育中的高亲和性优势亲本。
3.2 杂交组合的果实性状和子代苗期生长性状的超亲现象
在所有杂交组合中,杂交组合CB1×CG1、CB1×CM1 和CB1×CJ1 果皮厚度均较小, 而以CG5、CG6 和CG2 为母本的杂交果实果皮均较厚,对比亲本果实厚度可以看出,果皮厚的母本,其杂交子代果皮均表现偏厚的趋势。各杂交组合果实果皮厚度随亲本(尤其是母本)果皮厚度呈现不同的變化趋势;在所有杂交组合中,杂交组合CG2×CL1、CG6×CB1 和CF1×CS1 果实较大,其亲本果实也较大,说明其能保持亲本优良性状;杂交组合CS1×CG2、CG5×CH1 和CG5×CB1 的种子不但数量多,而且能保持较高水平的发芽率。在对杂交苗的观察中发现,以母本CS1 杂交组合的小苗总体生长表现较好,其中杂交组合CS1×CG2、CS1×CG4 和CS1×CD1 小苗生长最旺盛,6 月苗高达13 ~ 14 cm,其他组合的苗枯萎凋亡,或高6 ~ 9 cm,明显矮于上述3 个杂交组合。另外,CS1×CG2、CS1×CG4 和CS1×CD1 这3 个杂交组合在苗高方面相对于亲本CS1 具有一定的超亲现象。
3.3 研究局限及研究方向
地理环境、气候等多方面因素均会对油茶子代的表型观测分析产生影响,例如极端干旱、寒冷、洪涝等灾害会对油茶树造成不同程度的损害,本研究中未考虑这些因素的影响。下一步可考虑通过连续几年杂交试验并增加杂交组合,丰富高州油茶杂交群体系统。
致谢:栗永娟、李芳、粟依婷、程俊森和潘秋玲参与了亲本自然结实、花朵数量统计和杂交授粉情况调查。
参考文献:
[1] 中国科学院中国植物志编辑委员会. 中国植物志[M]. 北京:科学出版社,1998:11-13.
Editorial Committee of Chinese Flora, CAS. Flora ReipublicaePopularis Sinicae[M]. Beijing: Science Press,1998:11-13.
[2] 庄瑞林. 中国油茶[M]. 2 版. 北京: 中国林业出版社,2008:55-58.
ZHUANG R L. ChinaCamellia oleifera[M]. 2nd ed. Beijing:China Forestry Publishing House,2008:55-58.
[3] 庄朱辉. 油茶扦插育苗实验研究[J]. 福建林业科技,2003,30(3):83-85,98.
ZHUANG Z H. Experimental study onCamellia oleifera cuttingseedling raising[J]. Journal of Fujian Forestry Science and Technology,2003,30(3):83-85,98.
[4] 陈永忠, 王湘南. 油茶生物技术育种研究前景展望[J]. 湖南林业科技,2005,32(4):5-7.
CHEN Y Z, WANG X N. The outlook of the biotechnologybreeding research on oil teaCamellia[J]. Hunan Forestry Science &Technology,2005,32(4):5-7.
[5] 姚克平. 油茶优良品系引种栽培试验研究[J]. 山东林业科技,2005(6):8-10.
YAO K P. The horticultural experiment onCamellia oleifera fineclonal strain and family[J]. Journal of Shandong Forestry Scienceand Technology,2005(6):8-10.
[6] 常维霞, 姚小华, 龙伟, 等.4 个油茶物种杂交亲和性分析[J].植物研究,2016,36(4):527-534.
CHANG W X, YAO X H, LONG W, et al. Cross-compatibility offour kinds ofCamellia species[J]. Bulletin of Botanical Research,2016,36(4):527-534.
[7] 李晓春, 陈刚, 罗颖, 等. 11 个油茶无性系的杂交亲和力[J].亚热带农业研究,2020,16(1):19-23.
LI X C, CHEN G, LUO Y, et al. Cross compatibility among elevenCamellia oleifera clones[J]. Subtropical Agriculture Research,2020,16(1):19-23.
[8] 江泽鹏, 曾雯珺, 夏莹莹, 等. 不同地理种源油茶无性系杂交果实及F1 代苗期特性[J]. 广西林业科学,2018,47(2):212-215.
JIANG Z P, ZENG W J, XIA Y Y, et al. Hybrid fruits ofCamelliaoleifera from different geographical provenances and seedlinggrowth characteristics of F1 generation[J]. Guangxi Forestry Science,2018,47(2):212-215.
[9] 梁恬, 李祥胜, 李煜, 等. 普通油茶优良无性系有性杂交果实性状变异分析[J]. 森林与环境学报,2022,42(2):166-173.
LIANG T, LI X S, LI Y, et al. Variations in fruit traits in cross-bredCamellia oleifera clones[J]. Journal of Forest and Environment,2022,42(2):166-173.
[10] 林萍, 王開良, 姚小华, 等. 普通油茶杂交子代幼林经济性状的遗传分析[J]. 中南林业科技大学学报,2017,37(12):31-38.
LIN P, WANG K L, YAO X H, et al. Genetic analysis of the economictraits ofCamellia oleifera F1 descendant in half-diallel cross design[J].Journal of Central South University of Forestry & Technology,2017,37(12):31-38.
[11] 林萍, 姚小华, 滕建华, 等. 油茶5×5 全双列杂交子代幼林生长性状的配合力分析[J]. 中南林业科技大学学报,2016,36(5):26-32.
LIN P, YAO X H, TENG J H, et al. Analysis of genetic effects ofgrowth traits ofCamellia oleifera F1 descendants in complete diallelcross design Ⅲ[J]. Journal of Central South University of Forestry &Technology,2016,36(5):26-32.
[12] 曾武, 黎建伟, 林锦容, 等. 高州油茶种质资源及开发利用[J].中国林副特产,2016(1):95-96,100.
ZENG W, LI J W, LIN J R, et al. Resource development andutilization ofCamellia gauchowensis[J]. Forest By-Product andSpeciality in China,2016(1):95-96,100.
[13] 奚如春, 邓小梅. 我国油茶产业化发展中的现状、要素及其优化[J]. 经济林研究,2005,23(1):83-87.
XI R C, DENG X M. Industrialing oilteaCamellia production inChina: current status, limiting factors and optimal approaches[J].Non-wood Forest Research,2005,23(1):83-87.
[14] 原姣姣, 王成章, 陈虹霞, 等. 不同品种油茶籽的含油率和脂肪酸组成分析研究[J]. 中国油脂,2012,37(1):75-79.
YUAN J J, WANG C Z, CHEN H X, et al. Oil content and fattyacid composition analysis of different varieties ofCamelliaoleifera seeds[J]. China Oils and Fats,2012,37(1):75-79.
[15] 杨颖, 张鹏, 奚如春, 等. 高州油茶不同产区果实含油率及脂肪酸组成的变异特征[J]. 经济林研究,2018,36(4):104-108,144.
YANG Y, ZHANG P, XI R C, et al. Variation characteristics ofoil content and fatty acid composition inCamellia gauchowensisfruits at different producing areas[J]. Non-wood Forest Research,2018,36(4):104-108,144.
[16] 王章榮, 白嘉雨. 充分运用杂交技术加速我国林木育种进程[C]// 南方林木遗传育种研究. 北京: 中国林业出版社,2006:4-6.
WANG Z R, BAI J Y. Making full use of hybrid technology tospeed up the process of tree breeding in China[C]//Study onForest Tree Genetic Breeding in Southern China. Beijing: ChinaForestry Publishing House,2006:4-6.
[17] 陈永忠, 许彦明, 张震, 等. 油茶果实主要数量性状分析及育种指标体系筛选[J]. 中南林业科技大学学报,2021,41(3):1-9.
CHEN Y Z, XU Y M, ZHANG Z, et al. Analysis of fruit mainquantitative traits and selection of breeding index inCamelliaoleifera[J]. Journal of Central South University of Forestry &Technology,2021,41(3):1-9.
[18] 张应中, 张坤昌, 廖柏勇, 等. 基于混合线性模型的高州油茶杂交亲和性遗传评估Ⅰ [J]. 经济林研究,2022,40(3):1-13.
ZHANG Y Z, ZHANG K C, LIAO B Y, et al. Geneticevaluation for the cross-compatibility between lines ofCamelliagauchowensis and other species based on mixed linear model[J].Non-wood Forest Research,2022,40(3):1-13.
[19] 李荣丽, 黄寿先, 李志先, 等. 大叶栎家系水平性状变异及优良家系选择研究[J]. 广东农业科学,2014(17):47-57.
LI R L, HUANG S X, LI Z X, et al. Variation of traits amongCastanopsis fissa families and selection of superior families[J].Guangdong Agricultural Sciences,2014(17):47-57.
[20] 林元震, 陈晓阳. R 与ASReml-R 统计分析教程[M]. 北京:中国林业出版社,2014.
LIN Y Z, CHEN X Y. R and ASReml-R statistical analysiscourse[M]. Beijing: China Forestry Publishing House,2014.
[21] 陈晓阳, 沈熙环. 林木育种学[M]. 北京: 高等教育出版社,2006.
CHEN X Y, SHEN X H. Forest breeding science[M]. Beijing:Higher Education Press,2006.
[22] 馬晓雨, 刘焕臻, 孙国语, 等. 美洲黑杨× 大青杨染色体制片优化及核型分析[J]. 中南林业科技大学学报,2021,41(7):142-149.
MA X Y, LIU H Z, SUN G Y, et al. Chromosome preparationoptimization and karyotype analysis ofPopulus americana ×Populus daqing[J]. Journal of Central South University ofForestry & Technology,2021,41(7):142-149.
[23] 李拓键, 屈燕, 王兵益, 等. 不同倍性云南山茶染色体核型分析[J]. 中南林业科技大学学报,2023,43(3):167-174.
LI T J, QU Y, WANG B Y, et al. Chromosome karyotype analysisofCamellia reticulata with different ploidy[J]. Journal of CentralSouth University of Forestry & Technology,2023,43(3):167-174.
[24] HENDERSON C R. A simple method for computing the inverseof a numerator relationship matrix used in prediction of breedingvalues[J]. Biometrics,1976,32(1):69-83.
[25] 张乃燕, 蒙敬彪, 江泽鹏, 等. 广西油茶杂交育种研究现状与发展思路[J]. 林业科技开发,2008,22(3):6-9.
ZHANG N Y, MENG J B, JIANG Z P, et al. Research statusand development ideas of cross breeding of GuangxiCamelliaoleifera[J]. Journal of Forestry Engineering,2008,22(3):6-9.
[26] CHAUDHARY B D, KAKAR S N, SINGH R K. Comparison ofdiallel and its modificiations[J]. Silvae Genetica,1977,26(2/3):112-114.
[27] TALBERT J T. An advance-generation breeding plan for thenorth carolina state university-industry pine tree improvementcooperative[J]. Silvae Genetica,1979,28:72-75.
[28] YEH F C, HEAMAN J C. Estimating genetic parameters of heightgrowth in seven-year old coastal Douglas-fir from disconnecteddiallels[J]. Forest Science,1987,33:946-957.
[29] HUBER D A, WHITE T L, LITTELL R C, et al. Ordinary leastsquares estimation of general and specific combining abilitiesfrom half-diallel mating designs[J]. Silvae Genetica,1992,41(4/5):263-273.
[30] KING J N, CARSON M J, JOHNSON G R. Analysis ofdisconnected diallel mating designs: Ⅱ -results from a thirdgeneration progeny test of the New Zealand radiata pineimprovement programme[J]. Silvae Genetica,1998,47(2/3):80-87.
[31] 齐明. 杉木育种中GCA 与SCA 的相对重要性[J]. 林业科学研究,1996,9(5):498-503.
QI M. The relative importance of GCA and SCA in Chinese firbreeding[J]. Forest Research,1996,9(5):498-503.
[ 本文编校:闻 丽]
猜你喜欢
中学生数理化·八年级物理人教版(2022年6期)2022-06-05
今日农业(2021年10期)2021-11-27
疯狂英语·初中天地(2021年6期)2021-08-06
中央民族大学学报(自然科学版)(2017年1期)2017-06-11
中央民族大学学报(自然科学版)(2016年3期)2016-06-27
兽医导刊(2016年6期)2016-05-17
浙江农业科学(2016年11期)2016-05-04
广西林业科学(2016年2期)2016-03-20
福建稻麦科技(2015年1期)2016-01-06
中央民族大学学报(自然科学版)(2015年2期)2015-06-09
