
利用生物声学指标评估暗针叶林鸣禽围攻事件中的鸣声特征
2024-01-30 03:07王江月田佳周正暘马晓昀龙玉王戎疆李晟
北京大学学报(自然科学版) 2024年1期
王江月 田佳 周正暘 马晓昀 龙玉 王戎疆 李晟
北京大学学报(自然科学版) 第60卷 第1期 2024年1月
Acta Scientiarum Naturalium Universitatis Pekinensis, Vol. 60, No. 1 (Jan. 2024)
10.13209/j.0479-8023.2023.099
国家自然科学基金区域创新发展联合基金重点项目(U23A2019)资助
2022–12–29;
2023–03–30
利用生物声学指标评估暗针叶林鸣禽围攻事件中的鸣声特征
王江月 田佳 周正暘 马晓昀 龙玉 王戎疆 李晟†
北京大学生命科学学院, 北京 100871; †通信作者,E-mail: shengli@pku.edu.cn
利用鸣声播放实验, 并结合自动录音技术, 综合使用多种声学指数, 探究亚高山森林生态系统中, 森林鸣禽在围攻事件中鸣声的声学特征及其变化规律。2021 年夏季, 在四川王朗国家级自然保护区冷云杉暗针叶林中分别播放领鸺鹠()鸣声和暗绿柳莺()报警鸣叫来引发鸣禽围攻, 使用自动录音设备记录播放期间及播放前后在实验位点上自由活动的森林鸣禽的鸣声。设置 12 个位点, 每个位点进行领鸺鹠和暗绿柳莺播放实验各一次。24 次实验共记录到分属 3 目 17 科的 28 种鸟类参与围攻行为, 实验录音总有效时长为 1057 分钟。对围攻行为的观察和对录音记录的声学指数分析结果表明: 1)领鸺鹠鸣声和柳莺报警鸣叫都可以有效地引发鸣禽的围攻行为, 柳莺类与山雀类鸣禽是参与围攻的主体; 2)两种鸣声播放期间, 森林鸣禽的鸣声声强均显著高于播放前(领鸺鹠鸣声实验 SPLavg=−31.02±4.87dB(播放期间), SPLavg=−42.74±4.68dB(播放前空白段),=0.001; 暗绿柳莺报警鸣叫回放实验 SPLavg=−33.26±4.05dB (播放期间), SPLavg=−46.38±4.54dB(播放前空白段),=0.001), 鸣声复杂度均显著低于播放前(领鸺鹠鸣声 实验=0.76±0.02(播放期间),=0.80±0.03(播放前空白段),=0.001; 暗绿柳莺报警鸣叫回放实验=0.77±0.02(播放期间),=0.82±0.02(播放前空白段),=0.001); 3)播放领鸺鹠鸣声或暗绿柳莺报警鸣叫并未引发声学指数变化程度的显著不同。研究结果从声学层面为鸟类行为研究提供新的思路, 同时为大范围的声景监测数据提供新的行为学应用场景。
自动录音技术; 声学指数; 鸟类围攻行为; 鸣声播放实验; 森林鸣禽; 王朗国家级自然保护区
围攻行为(mobbing behavior)在鸟类中广泛存在, 通常由被捕食者向捕食者、受侵害者向侵害者或者弱势方向优势方发起, 用来干扰、驱逐潜在的捕食者或侵害者(如种间巢寄生鸟类), 从而保卫自身、同类、幼鸟或繁殖巢的安全[1–2]。在鸟类的围攻事件中, 诱发围攻行为的通常是单只或少数具有潜在威胁的猛禽或其他鸟类, 周围一定范围内的多种鸟类组成针对该目标的围攻群体[3], 以高频次、高强度的大声鸣叫对其进行骚扰、驱赶, 或直接进行物理攻击[4–5]。强烈的围攻行为可以使捕食者放弃捕食机会, 甚至改变巢穴选址[6]。鸟类在围攻时发出的鸣声与平时的鸣声明显不同, 针对不同类型围攻目标的鸣声也不同[7]。在围攻事件中, 最早发现目标并开始鸣叫的鸟类在围攻事件的发起中可能扮演重要角色[8], 其他鸟类可以快速地在同种或非同种鸟类的鸣叫中识别风险信息, 其中部分鸟类继而加入围攻之中[9–11]。
以鸣声为主要表现形式的声音信号是鸟类交流信息和识别风险的主要方式之一[12], 对围攻行为的研究大多从行为观察方面着手[13]。在围攻过程中, 鸟类鸣声会由围攻行为发生之前的低强度和多样化鸣声转变为围攻中的高频率和高强度报警鸣叫。虽然这一现象在相关的行为观察中被广泛记述, 但现有研究缺乏对围攻行为发生期间围攻鸟类鸣声特征及其变化动态的量化描述。随着录音和声学记录技术的发展, 研究人员开发出多种声学指数, 可以从不同的维度反映声音在频率、振幅和时间方面的特征, 并广泛用于物种识别、行为描述和多样性监测等[14–15]。近年来, 有研究者利用声学指数, 量化地描述单一鸟类物种在参与围攻时鸣声的变化[16]。随着声景生态学(soundscape ecology)概念的普及和自动录音技术(automatic acoustic recording, AAR)的成熟与推广[17], 研究人员开始使用声学指数, 大批量地处理和量化声音信息[18], 用于评估鸟类多种行为中鸣声的声学特征与多样性[19]。利用这些在生物声学领域新兴的自动化检测手段和高通量分析方法, 我们可以在鸟类围攻行为的研究中, 通过声学指数衡量鸟类围攻行为中的声学特征, 进而指示小尺度范围鸟类的丰度(数量)、丰富度(物种数)和鸣声模式(鸣唱或鸣叫)。
鸣声播放实验是探究鸟类对潜在风险反应的有效手段[20]。在围攻行为研究中, 经常通过播放捕食者鸣声或围攻鸟类报警鸣叫的方式, 引发自然环境中鸟类的围攻行为, 观察参与围攻的鸟类对激发信号的反应[21–24]。
本研究通过鸣声播放实验, 在中国西南山地的亚高山暗针叶林中引发鸣禽的围攻行为, 通过自动声学记录与生物声学指标量化分析, 期望回答以下两个问题: 1)在围攻事件中, 围攻鸟类的鸣声表现出怎样的声学特征, 围攻前后发生怎样的变化? 2)不同类型(小型猛禽鸣声或者围攻鸟类的报警鸣叫)的鸣声播放实验引发的围攻事件在鸣声特征上有何差异?
1 材料与方法
1.1 实验地点
本研究的野外调查于 2021 年 7 月开展, 实验地点位于四川省平武县王朗国家级自然保护区(32.49° —33.02°N, 103.55°—104.10°E)。王朗保护区成立于1963 年, 以大熊猫()、扭角羚()以及川金丝猴()等珍稀濒危动物及其栖息地为主要保护对象, 是我国最早成立的大熊猫保护区之一[25]。保护区地处岷山山系中部, 面积为 323km2, 海拔范围为 2300~4980m, 属于丹巴–松潘半湿润气候区。区内分布大片原始针叶林, 是川西暗针叶林带的典型代表[26]。保护区内共记录有 271 种鸟类, 隶属于 16目 55 科。雀形目物种数最多, 有 194 种; 鸮形目鸟类有 7 种, 包括红角鸮()、雕鸮()、灰林鸮()、四川林鸮()、领鸺鹠()、长耳鸮()和短耳鸮(), 其中领鸺鹠是体型最小的一种[27]。
选择保护区内大窝凼沟的沟谷林区布设实验点(图 1)。实验区域内海拔为 2800~3000m, 7 月平均气温为 12.8℃, 月降雨量约为 131mm。区域内植被类型为原生亚高山暗针叶林, 乔木层优势种为岷江冷杉(var.)和紫果云杉(), 有缺苞箭竹()和多种灌木密生于林下[28–29]。
1.2 实验音频准备
从 Xeno-canto 鸟类鸣声数据库(http://www.xeno- canto.org/)获取 1 段时长为 35s 的领鸺鹠高质量鸣声录音(数据库记录编号为 XC499489, 采集自峨眉峰自然保护区), 其鸣声频率范围在 0.6~1.6kHz 之间。使用 Adobe Audition 2021 (Adobe Systems, Inc., CA, USA)去除 2.0kHz 以上的频段, 降低背景噪音(图 2(a))。将这段领鸺鹠鸣声录音处理制作为循环播放的时长为 15 分钟的音频, 然后在前后各加入时长 15 分钟的空白音频, 形成总时长为 45 分钟的领鸺鹠鸣声实验音频。在对王朗保护区的前期预调查中, 发现在围攻行为中柳莺类鸟类的报警鸣叫最为频繁和强烈, 于是在实验区域实地录制了暗绿柳莺()的报警鸣叫, 单段时长 60秒, 其鸣叫频率范围为 6.3~9.4kHz 之间。然后, 利用 Adobe Audition 去除 5.0kHz 以下的频段来降低背景噪音, 并增益振幅 10dB 来还原实地录制时的报警鸣叫声强(图 2(b))。参照领鸺鹠鸣声实验音频的制作方法, 制作了总时长为 45 分钟的包含前后各15 分钟空白的暗绿柳莺报警鸣叫实验音频。
1.3 录音播放实验
本研究共选择 12 个调查点(图 1), 相邻位点的间距不小于 100m。在每个实验位点, 选择距离地面约 2m 处的树枝放置 1 只领鸺鹠模型, 在其下方 1m 处设置一部音箱(EXOGEAR, Inc., CA, USA), 用来播放实验音频。在每个实验点设置一台 Wildlife AcousticsTMSM4 鸟类鸣声记录仪(Wildlife Acous-tics, Inc., MA, USA), 采用双通道记录, 文件格式为16bit.wav, 内置麦克风指向特性为全向, 采样频率设置为 32kHz(可记录最高频率为 16kHz 的动物叫声, 记录带宽为 0~16kHz), 灵敏度设置为默认值−35±4dB (1kHz), 信噪比为默认值 80dB (1kHz)。实验装置安装完毕即开始播放实验音频, 实验人员隐蔽在实验装置 10m 之外, 观察并记录实验点周围活动的鸟类以及参与围攻行为鸟类的种类、数量与活动情况。
在每个位点分别播放领鸺鹠鸣声实验音频和暗绿柳莺报警鸣叫实验音频各 1 次, 共完成 24 次实验。为避免鸟类对播放的音频习惯化, 或因领鸺鹠鸣声而对该位点产生趋避, 在每个位点开展领鸺鹠鸣声播放实验后, 至少间隔 2 天, 再进行暗绿柳莺报警鸣叫回放实验。
1.4 数据分析
1.4.1数据处理
在鸟类鸣声记录仪记录到的音频文件中, 删除1.6kHz 以下和 6.3kHz 以上(即领鸺鹠鸣声和暗绿柳莺报警鸣叫对应的频率范围)的频段, 消除其对后续声学指数计算的影响。为避免实验装备设置和拆除时人为噪音的干扰, 将音频文件统一裁剪为时长35 分钟的文件, 包括 10 分钟播放前空白(Pre)、15分钟鸣声播放(During)和 10 分钟播放后空白(Post)这 3 个时期, 获得统一标准的 24 个分析音频。由于部分位点的实验受降雨天气干扰, 个别分析音频的播放后空白段可能不足 10 分钟。
1.4.2声学指数计算与分析
分别使用声压级(sound pressure level, SPL)、声音多样性指数(acoustic diversity index, ADI)和声学熵指数(entropy,)衡量鸟类鸣声的声强、多样性和复杂度。
声压级是反映音量大小的指标, 单位为分贝(dB), 计算公式为
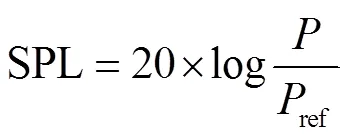
其中,表示声压, 单位为帕斯卡(Pa);ref为参考声压[30]。本研究中,ref表示鸟类鸣声记录仪可记录到的最高声压, 即输入音频为 1kHz 时,ref=1Pa。声压级越高, 表明鸣声越强烈。当每只鸟类鸣声的声强保持一定数值时, 声压级与发出鸣声的鸟类数量正相关。
声音多样性指数[31]是一个反映声音多样性的指标, 计算公式为
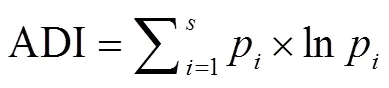
其中,p表示声信号在第种频段所占比例,表示频段总数量。ADI 取值大于 0, 且没有单位。声音多样性指数越高, 说明鸣声的频率越多样化; 由于单一物种鸟类的鸣声频段通常是固定的, 声音多样性指数与发出鸣声的鸟类物种数之间往往存在正相关关系[31]。
底图来自Google Earth
图1 本研究调查位点在四川王朗国家级自然保护区中的位置
Fig. 1 Survey locations of this study in Wanglang National Nature Reserve, Sichuan Province
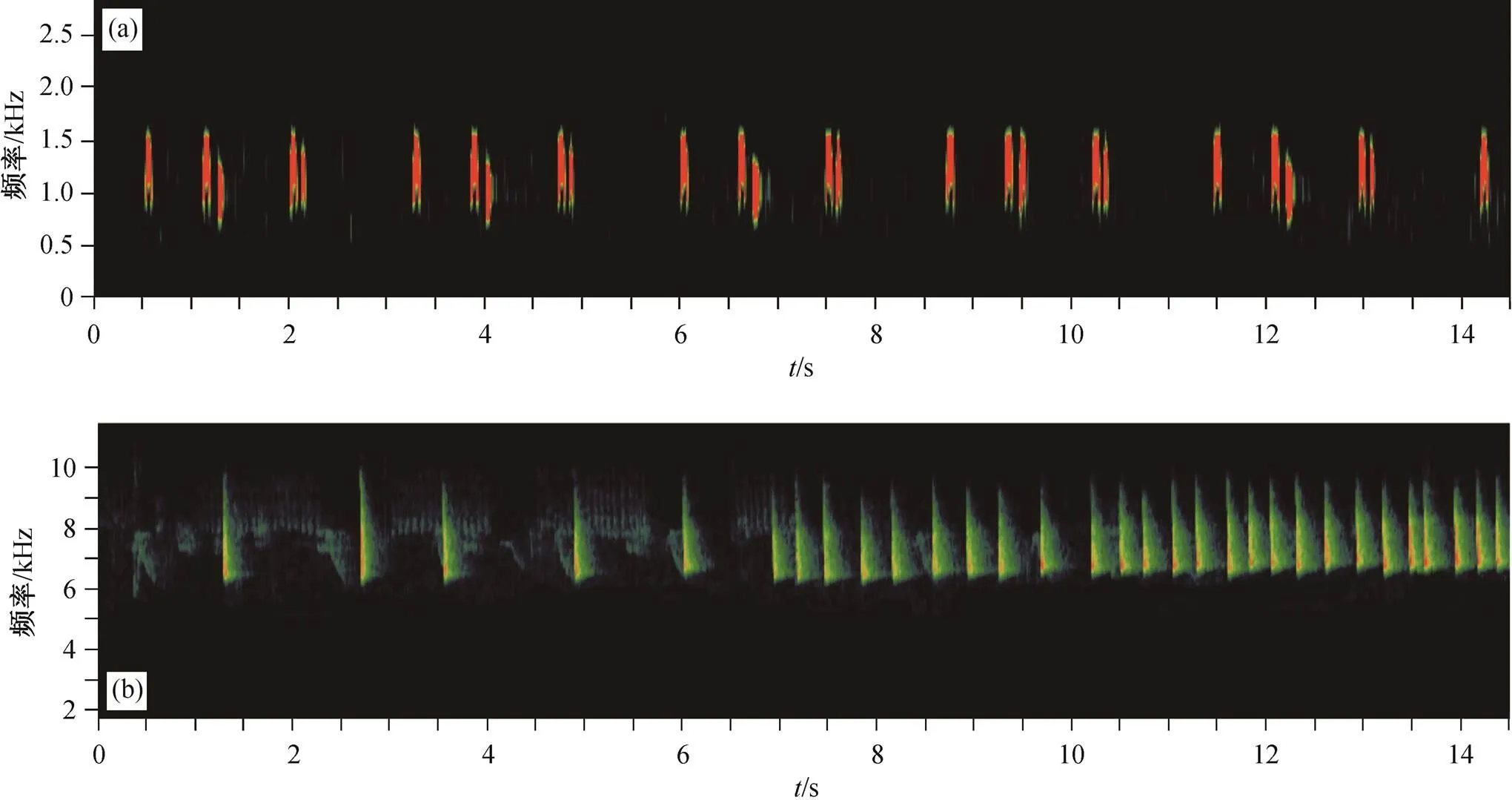
图2 领鸺鹠鸣声(a)和暗绿柳莺报警鸣叫(b)的声谱
声学熵指数[32]是一个反映声信号在时频域的复杂度和均匀度的指标, 是目前在生物多样性监测中使用最多的声学指数[33], 计算公式为
=t×f,
其中,t表示时间熵, 指示振幅的均匀度;f表示频谱熵, 指示频率的均匀度。取值范围在 0~1 的闭区间内, 且没有单位。声学熵指数越高, 说明鸣声越复杂, 在一定时间内产生的鸣声频率和振幅的变化越大。一般来说, 鸟类鸣唱的声学熵指数比鸣叫 更高。
使用 Kaleidoscope Pro软件(Wildlife Acoustics, Inc., MA, USA)计算 24 个分析音频中每分钟的平均声压级, 使用 R 4.1.0[34]中的 soundecology 包[35]计算各分析音频中每个时期的声音多样性指数和声学熵指数。利用配对的 Wilcoxon 符号秩检验, 并用 Bon-ferroni 法校正值, 比较鸣声播放前、播放中和播放后这 3 个时期间各声学指数的差异, 用来指示围攻行为鸣声的声强、多样性和复杂度特征, 进而揭示围攻事件引起的鸟类数量、物种数和鸣声模式变化。利用配对的 Wilcoxon 符号秩检验, 比较领鸺鹠鸣声播放实验和暗绿柳莺报警鸣叫回放实验中实验音频播放引起鸣声声学指数变化程度的差异(即播放时和播放前的数值差, 可以认为不受时间和地点的影响), 以便揭示两种实验引起的鸟类数量、物种数和鸣声模式变化程度是否存在差异。
2 结果
在全部 24 次播放实验中, 领鸺鹠鸣声和暗绿柳莺报警鸣叫均可引起附近鸟类的报警鸣叫与围攻行为, 包括围绕音箱和领鸺鹠模型在树丛间频繁进出、持续发出高声报警鸣叫等。在播放实验中共记录到分属 3 目 17 科的 28 种鸟参与了围攻行为(详见附录) (访问 http://xbna.pku.edu.cn 查看附录)。
在参与围攻的鸟类中, 暗绿柳莺往往最先发出报警鸣叫。柳莺类(包括暗绿柳莺、橙斑翅柳莺等)和山雀类(包括黑冠山雀和褐冠山雀等)是参与围攻的主要类群。在领鸺鹠鸣声或暗绿柳莺报警鸣叫播放期间, 围攻行为会一直持续; 播放结束后, 鸟类的围攻行为也随之结束。
全部 24 次实验中, 鸟类鸣声记录仪的总有效记录时长为 1057 分钟。其中, 位点 1 和 2 的领鸺鹠鸣声播放是人工手动控制的, 鸣声播放期持续20分钟(后裁剪为 15 分钟); 在位点 3, 4 和 12 进行领鸺鹠鸣声实验时, 遇强降雨天气, 鸣声播放后的空白音频分别持续 2, 5 和 5 分钟, 鸣声播放期未受影响。
领鸺鹠鸣声播放实验中, 森林鸣禽的报警鸣叫在鸣声播放期间平均声压级 SPLavg=−31.02±4.87 dB, 显著高于播放前空白段(SPLavg=−42.74±4.68 dB)(Wilcoxon 检验统计量=0,=0.001), 也显著高于播放后的空白段(SPLavg= −34.52±6.83 dB)(= 69,=0.048); 同时, 鸣声播放后空白段的平均声压级显著高于播放前的空白段(=4,=0.010)(图3(a1))。在声音多样性指数上, 鸣声播放前(ADI= 1.69±0.12)、播放期间(ADI=1.59±0.10)与播放后(ADI=1.67±0.14) 3 个时期两两相比, 均无显著差异(图 3(b1))。在声学熵指数上, 鸣声播放前的空白段(=0.80±0.03)显著高于播放期间(=0.76±0.02) (=78,=0.001)(图 3(c1)), 播放期间与播放后(=0.77±0.03)相比无显著差异, 播放前与播放后相比亦无显著差异。
领鸺鹠鸣声播放实验和暗绿柳莺报警鸣叫回放实验中, 实验音频播放时和播放前的声学指数变化程度在两种鸣声下没有表现出显著差异(图 4)。
3 讨论
在本项实验中, 无论是播放领鸺鹠鸣声还是暗绿柳莺报警鸣叫, 均可引发鸟类的围攻行为。鸣声播放前后相比, 播放期间的声学指数存在一定的差异。鸣声播放期间的平均声压级均显著高于播放前后空白段, 声学熵指数也都显著低于播放前后空白段, 表明鸣声播放期间的鸟类鸣声声强变高, 复杂度变低。单一、强烈的报警鸣叫可以在物种间有效地传递受威胁的信息[1], 号召更多鸟类加入围攻并鸣叫, 使得总体上鸣声的平均声压级更高, 从而达到成功驱逐捕食者、保卫自身及幼鸟安全的目的。平均声压级的升高表明, 鸣声播放引发的围攻行为中, 鸟类丰度变高。本项实验开展期间是当地鸟类的繁殖期, 柳莺和山雀等森林鸣禽的鸣声中有较大比例是复杂婉转的鸣唱(song), 具有相对较高的声学复杂度。在围攻事件发生期间, 这些鸟类的鸣唱大大减少甚至终止, 鸣声变成以单调、重复的报警鸣叫(call)为主, 这可能是本研究的播放实验期间记录到鸟类鸣声的声学复杂度降低的主要原因。
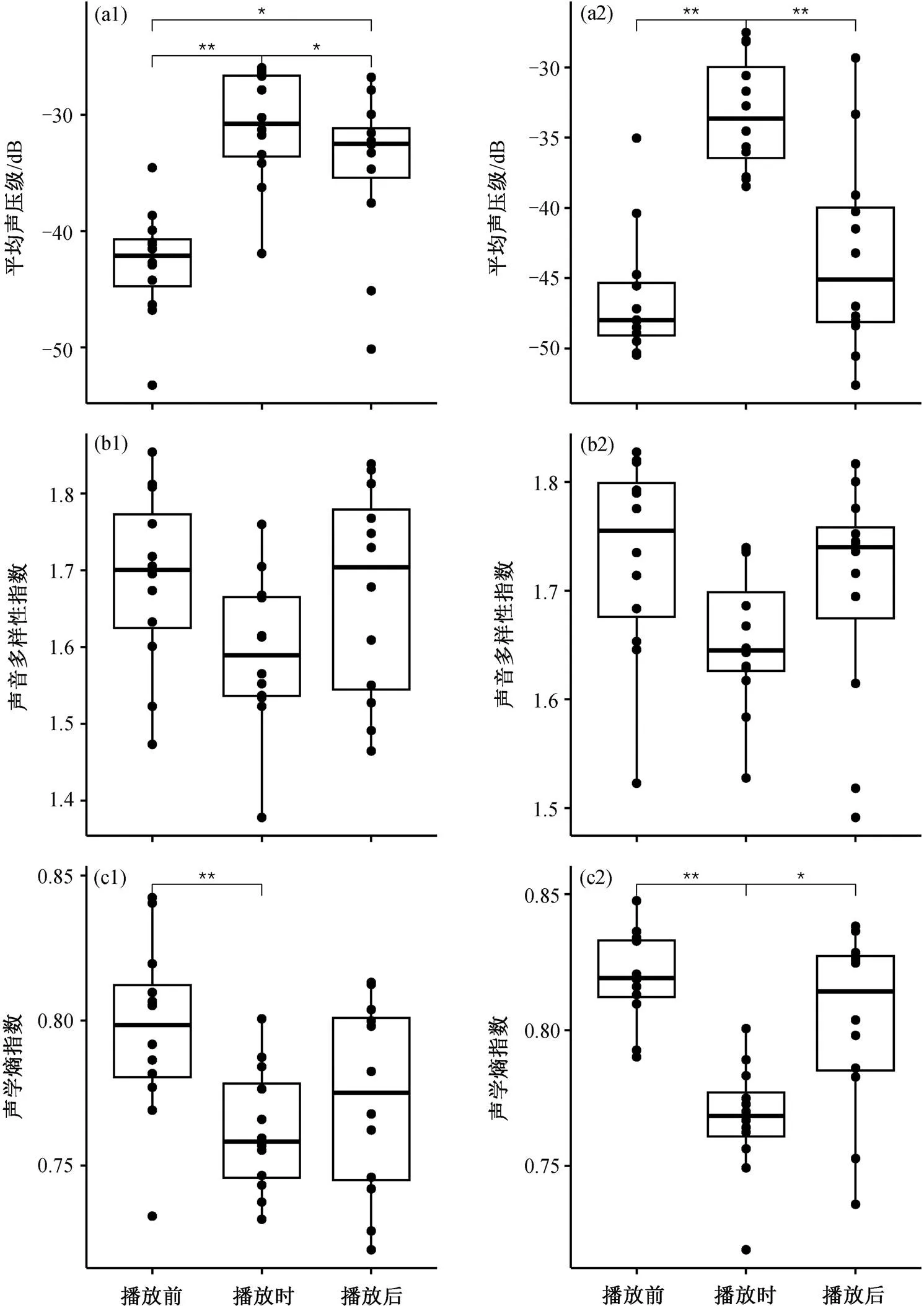
(a1), (b1)和(c1): 领鸺鹠鸣声播放实验; (a2), (b2)和(c2): 暗绿柳莺报警鸣叫回放实验
声音多样性指数可以指示频率的多样化程度。在本项实验鸣声播放期间和播放前、后这 3 个阶段均没有显著差异。围攻事件中, 同一种鸟的报警鸣叫频段较为单一[36], 由于在围攻行为中担任不同的角色, 传递不同的信息[11], 不同物种的鸟的报警鸣叫存在差异[37]。虽然同种鸟的鸣叫与鸣唱明显不同, 但同种鸟的鸣声频段是一定的, 而不同鸟种的主要鸣声频段往往有较大的差异[38]。不同鸟种差异化的鸣声频段组成播放实验前、中、后的鸣声频率范围, 频率的多样性指示了小尺度范围的鸟类丰富度。声音多样性指数在实验前、中、后无显著差异, 说明在鸟类鸣声记录仪工作范围内鸟类物种没有发生显著的变化。这与我们在播放实验期间的目视观察结果一致, 即播放实验没有从远处吸引来新的鸟种加入围攻行为, 实验点周围已有的鸟种也未在播放实验期间离开。
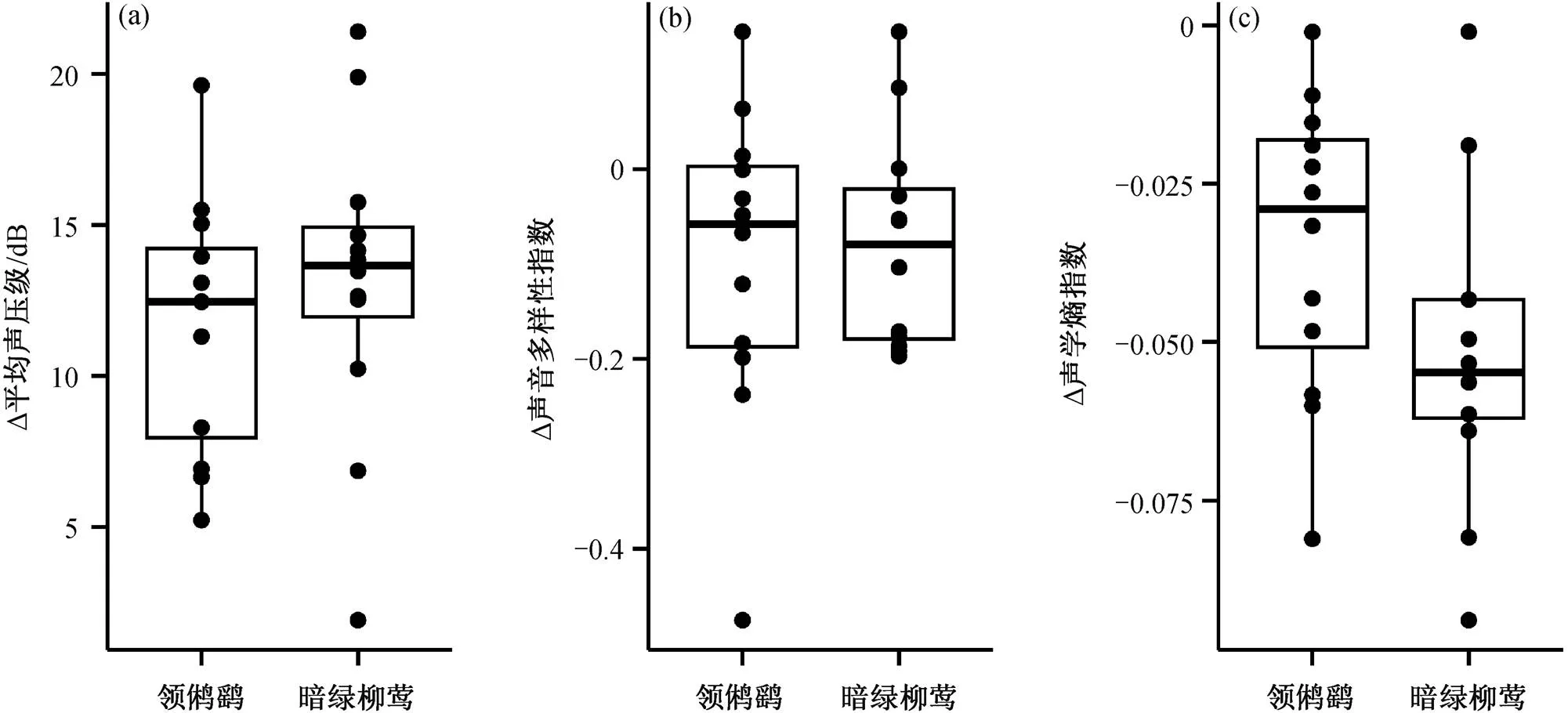
图4 领鸺鹠鸣声播放实验和暗绿柳莺报警鸣叫回放实验中, 实验音频播放前与播放时声学指数变化程度的对比
本文实验选择的领鸺鹠鸣声来源于峨眉峰自然保护区, 与实验地点王朗自然保护区有一定的距离, 两地领鸺鹠的鸣声之间可能存在地域性(方言)差异。对于本地可能参与围攻行为的鸟类来说, 外来的、相对陌生的鸣声可能会引起它们更高强度的报警鸣叫, 也有可能因无法被当地鸟种识别而不能有效地引起围攻行为。实验中播放的暗绿柳莺报警声来源于实验人员布设领鸺鹠模型时, 位点附近的暗绿柳莺对领鸺鹠模型和实验人员发出的报警鸣叫。虽然领鸺鹠鸣声可能存在其他地区的方言, 报警鸣叫仅为暗绿柳莺单一物种, 但本研究的两类鸣声播放实验中均发生围攻行为, 且播放时和播放前的声学指数变化程度在两种鸣声播放条件下没有显著差异, 说明实验选择的领鸺鹠鸣声与当地记录的暗绿柳莺报警鸣叫引发的围攻事件强度没有显著差异。这进一步说明, 作为最先开始报警的鸣禽之一, 暗绿柳莺的报警鸣叫在引发围攻行为上起到的作用与作为被围攻对象的领鸺鹠鸣声相当。在该森林生态系统的鸣禽群落中, 以暗绿柳莺为代表的最先开始鸣叫的柳莺类和山雀类可能担任警戒者角色, 其鸣叫中包含有效的入侵者信息, 且对鸟群具有普适的重要含义, 可以迅速召集周围个体加入围攻事件, 以期驱逐入侵者。然而, 由于本实验选择的领鸺鹠鸣声的单一性和可能存在的地域间差异, 领鸺鹠鸣声播放实验引起的围攻行为强度可能低于应有的水平, 导致对暗绿柳莺报警鸣叫作用的高估。未来对围攻事件具体过程的刻画仍需进一步研究, 例如详细记录当地围攻行为的鸟种、数量和具体行为, 识别围攻前后及围攻时的各个鸣声语句并量化声学特征等。同时, 为深入探究不同鸟种担任警戒者的差别, 可采集多种鸟类报警鸣叫的音频, 参考本研究的流程进行鸣声播放实验, 并利用平均声压级等声学指数, 量化围攻行为中的鸣声特征, 用来识别何种鸟类报警鸣叫引发围攻行为的效力最强。
另外, 我们在播放实验音频的音箱上方摆放了1 只领鸺鹠模型, 目的是为围攻鸟类提供视觉刺激, 同时便于研究人员观察参与围攻事件的鸟种及其数量与行为。本研究的鸣声播放期设置为 15 分钟, 这样的时长远大于自然情况下可引起围攻行为的听觉刺激, 目的是便于观察记录到明显的行为和鸣声变化。鸣声播放前, 单纯的视觉信号未能引发围攻事件或报警鸣叫; 鸣声播放时, 领鸺鹠模型成为围攻行为的物理中心, 更好地模拟了真实围攻行为情景。在暗绿柳莺报警鸣叫回放实验中, 作为入侵者的领鸺鹠仅保留了视觉信号而没有声音信号, 但播放警戒者的报警鸣叫依然可以有效地引发同等强度的围攻事件。这进一步印证了担任警戒者角色的鸟类的报警鸣叫对于群落中其他物种和个体的重要意义。
目前, 声学指数已广泛应用于声景生态学的相关研究与监测中, 如森林鸟类多样性监测、鸟类繁殖物候监测和不同位点的声景比较区分等[39]。本研究将声学熵指数和平均声压级结合使用, 可以有效地区分围攻行为发生和未发生的时段。因此, 在摸清鸟类典型行为过程中声学指标的特征与动态模式的基础上, 利用各种声学指标的组合可以对小尺度下不同时段的鸟类鸣声模式进行快速识别与区分, 进而指示不同的鸟类行为, 如求偶、育雏和围攻等, 为鸟类行为的研究提供声学角度的新思路, 同时也为大范围的声景监测数据提供新的行为学应用场景。
致谢 感谢北京大学生命科学学院“生态学野外实践”课程对本研究的支持。感谢王朗国家级自然保护区在本研究野外工作开展期间提供的支持与保障。
[1] Curio E, Ernst U, Vieth W. The adaptive significance of avian mobbing. II. Cultural transmission of enemy recognition in blackbirds: effectiveness and some constraints. Zeitschrift für Tierpsychologie, 1978, 48: 184–202
[2] Flasskamp A. The adaptive significance of avian mobbing: V. An experimental test of the ‘move on’ hypothesis. Ethology, 1994, 96: 322–333
[3] Betts M G, Hadley A S, Doran P J. Avian mobbing response is restricted by territory boundaries: experi-mental evidence from two species of forest warblers. Ethology, 2005, 111: 821–835
[4] Francis A M, Hailman J P, Woolfenden G E. Mobbing by Florida scrub jays: behavior, sexual asymmetry, role of helpers and ontogeny. Animal Behaviour, 1989, 38: 795–816
[5] Thompson T. Conditioned avoidance of mobbing call by chaffinches. Animal Behaviour, 1969, 17: 517–522
[6] Pavey C R, Smyth A K. Effects of avian mobbing on roost use and diet of powerful owls,. Animal Behaviour, 1998, 55: 313–318
[7] Naguib M, Mundry R, Ostreiher R, et al. Cooperatively breeding Arabian babblers call differently when mob-bing in different predator-induced situations. Behavio-ral Ecology, 1999, 10: 636–640
[8] Wheatcroft D, Price T D. Collective action promoted by key individuals. The American Naturalist, 2018, 192: 401–414
[9] Colombelli-Negrel D, Evans C. Superb fairy-wrens respond more to alarm calls from mate and kin com-pared to unrelated individuals. Behavioral Ecology, 2017, 28: 1101–1112
[10] Pell F S E D, Potvin D A, Ratnayake C P, et al. Birds orient their heads appropriately in response to func-tionally referential alarm calls of heterospecifics. Ani-mal Behaviour, 2018, 140: 109–118
[11] Kalb N, Anger F, Randler C. Subtle variations in mobbing calls are predator-specific in great tits (). Scientific Reports, 2019,9: 6572
[12] Pawlak P, Kwiecinski Z, Husek J. Mobbing of the top predator: a correlation between avian community richness and the number of mobbing species. Journal of Ornithology, 2019, 160: 665–672
[13] Forsman J T, Monkkonen M. Responses by breeding birds to heterospecific song and mobbing call play-backs under varying predation risk. Animal Behaviour, 2001, 62: 1067–1073
[14] Brodie S, Allen-Ankins S, Towsey M, Roe P, et al. Automated species identification of frog choruses in environmental recordings using acoustic indices. Eco-logical Indicators, 2020, 119: 106852
[15] Gan H, Towsey M, Li Y, Zhang J, et al. Animal call recognition with acoustic indices: little spotted kiwi as a case study // 2018 Digital Image Computing: Tech-niques and Applications. Canberra, 2018: 683–690
[16] Billings A C. The low-frequency acoustic structure of mobbing calls differs across habitat types in three pas-serine families. Animal Behaviour, 2018, 138: 39–49
[17] 赵莹, 申小莉, 李晟, 等. 声景生态学研究进展和展望. 生物多样性, 2020, 28(7): 806–820
[18] Fuller S, Axel A C, Tucker D, et al. Connecting soundscape to landscape: which acoustic index best describes landscape configuration?. Ecological Indica-tors, 2015, 58: 207–215
[19] Rajan S C, Athira K, Jaishanker R, et al. Rapid assess-ment of biodiversity using acoustic indices. Biodi-versity Conservation, 2019, 28: 2371–2383
[20] De Rosa A, Castro I, Marsland S. The acoustic play-back technique in avian fieldwork contexts: a syste-matic review and recommendations for best practice. IBIS, 2022, 164: 371–387
[21] Doran P J, Gulezian P Z, Betts M G. A test of the mobbing playback method for estimating bird repro-ductive success. Journal of Field Ornithology, 2005, 76: 227–233
[22] Jiang D M, Sieving K E, Meaux E, et al. Seasonal changes in mixed-species bird flocks and antipredator information. Ecology and Evolution, 2020, 10: 5368–5382
[23] Shimazaki A, Yamaura Y, Senzaki M, et al. Mobbing call experiment suggests the enhancement of forest bird movement by tree cover in urban landscapes across seasons. Avian Conservation and Ecology, 2017, 12: 16
[24] Yves T, Andre D. Playbacks of mobbing calls of black-capped chickadees help estimate the abundance of forest birds in winter. Journal of Field Ornithology, 2002, 73: 303–307
[25] 李晟之. 王朗志: 四川王朗国家级自然保护区志(1965—2015). 成都: 四川科学技术出版社, 2018
[26] 蒋有绪. 川西亚高山暗针叶林的群落特点及其分类原则. 植物生态学与地植物学丛刊, 1963(1): 42–50
[27] 尚晓彤, 罗春平, 李斌, 等. 四川王朗国家级自然保护区鸟类多样性与区系组成. 四川动物, 2020, 39(1): 93–106
[28] Taylor A H, Qin Z, Liu J. Structure and dynamics of subalpine forests in the Wang Lang Natural Reserve, Sichuan, China. Vegetatio, 1996, 124: 25–38
[29] 樊凡, 赵联军, 马添翼, 等. 川西王朗亚高山暗针叶林25.2 hm2动态监测样地物种组成与群落结构特征. 植物生态学报, 2022, 46(9): 1005–1017
[30] 漆安慎, 杜婵英. 普通物理学教程力学. 第 3 版. 北京: 高等教育出版社, 2012
[31] Villanueva-Rivera L J, Pijanowski B C, Doucette J, et al. A primer of acoustic analysis for landscape eco-logists. Landscape Ecology, 2011, 26: 1233–1246
[32] Sueur J, Aubin T, Simonis C. Seewave: a free modular tool for sound analysis and synthesis. Bioacoustics, 2008, 18: 213–226
[33] 伦可环, 张雁云, 夏灿玮. 基于声音指数的鸟类多样性监测. 生物学通报, 2017, 52(11): 1–5
[34] R Core Team. R: a language and environment for statistical computing [CP/OL]. Vienna: R Foundation for Statistical Computing. 2021 [2021–08–19]. https:// www.r-project.org
[35] Villanueva-Rivera L J, Pijanowski B C. Soundecology: soundscape ecology [CP/OL]. 2018 [2021–08–19]. https://cran.r-project.org/web/packages/soundecology
[36] Dutour M, Lena J P, Lengagne T. Mobbing calls: a signal transcending species boundaries. Animal Beha-viour, 2017, 131: 3–11
[37] Suzuki T N. Referential calls coordinate multi-species mobbing in a forest bird community. Journal of Etho-logy, 2016, 34: 79–84
[38] Priyadarshani N, Marsland S, Castro I, et al. Bird- song denoising using wavelets. PLOS ONE, 2016, 11: e0146790
[39] Bradfer-Lawrence T, Gardner N, Bunnefeld L, et al. Guidelines for the use of acoustic indices in environ-mental research. Methods in Ecology and Evolution, 2019, 10: 1796–1807
Evaluating the Characters of Songbirds’ Vocalizations during Mobbing Event in Dark Coniferous Forest Using Bio-acoustic Indicators
WANG Jiangyue, TIAN Jia, ZHOU Zhengyang, MA Xiaoyun, LONG Yu, WANG Rongjiang, LI Sheng†
School of Life Sciences, Peking University, Beijing 100871; † Corresponding author, E-mail: shengli@pku.edu.cn
This study investigated the acoustic variation of songbirds’ vocalization during mobbing event in the sub-alpine forests using playback experiment and automatic acoustic recording (AAR). We playedvocalization andalarm call to induce songbirds’ mobbing behavior and recorded their response vocalizations using automatic acoustic recorders in the-dark coniferous forest in Wanglang National Nature Reserve, Sichuan Province, during the summer of 2021. Altogether 24 experiments were performed on 12 experiment sites, with a total length of 1057 min acoustic recordings, including 12vocalization experiments, and 12alarm call experiments. A total of 28 species of birds belonging to 17 families and 3 orders were observed to participate in mobbing behavior. By analyzing the acoustic indexes of recordings, it was concluded that 1) bothvocalization andalarm call could trigger mobbing event, during which Phylloscopidae and Paridae species were the majority being involved; 2) during playback period, songbirds’ vocalizations are more intense (experiments: SPLavg=−31.02±4.87 dB vs −42.74±4.68 dB,= 0.001;experiments: SPLavg= −33.26±4.05 dB vs −46.38±4.54 dB,= 0.001) and less complicated (experiments:= 0.76±0.02 vs 0.80±0.03,= 0.001;experiments:= 0.77±0.02 vs 0.82±0.02,= 0.001) than that of pre-playback period; 3) the variability of acoustic indices from pre-playback to during-playback didn’t change significantly betweenexperiments andexperiments. The results provide a new insight into avian behavior studies in the acoustic aspect, and novel behavioral application scenarios for large-scale soundscape monitoring data.
automatic acoustic recording (AAR); acoustic indices; avian mobbing behavior; playback experiment; forest songbirds; Wanglang National Nature Reserve
猜你喜欢
东坡赤壁诗词(2023年2期)2023-05-30
小学阅读指南·高年级版(2021年9期)2021-10-09
诗歌月刊(2021年8期)2021-08-25
小天使·二年级语数英综合(2021年6期)2021-08-09
家庭影院技术(2020年6期)2020-07-27
阅读(科学探秘)(2019年4期)2019-06-19
家庭影院技术(2019年1期)2019-01-21
家庭影院技术(2018年11期)2019-01-21
安徽师范大学学报(自然科学版)(2018年5期)2018-12-11
家庭影院技术(2018年10期)2018-11-02
